
The Role of the PFNA Operon of Bifidobacteria in the Recognition of Host’s Immune Signals: Prospects for the Use of the FN3 Protein in the Treatment of COVID-19

 ,
,
Abstract
:1. Introduction
2. Bifidobacteria and the Human Immune System
3. Species-Specific PFNA Operon, Structure, Possible Functions
4. FN3: Species Specificity, Divergence, Structure, Function
5. Binding of the FN3 Protein Fragment to Cytokines (TNF-α)
6. TNF-α, IL-6 and Regulation of Cytokine Storms
7. Challenges and Limitations
8. Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tiwari, S.K.; Dicks, L.M.T.; Popov, I.V.; Karaseva, A.; Ermakov, A.M.; Suvorov, A.; Tagg, J.R.; Weeks, R.; Chikindas, M.L. Probiotics at War Against Viruses: What Is Missing From the Picture? Front. Microbiol. 2020, 11, 1877. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Paramanathan, R.; Gupta, R.S. Signature proteins that are distinctive characteristics of Actinobacteria and their subgroups. Antonie Leeuwenhoek 2006, 90, 69–91. [Google Scholar] [CrossRef]
- Gao, B.; Gupta, R.S. Phylogenetic Framework and Molecular Signatures for the Main Clades of the Phylum Actinobacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 66–112. [Google Scholar] [CrossRef] [Green Version]
- Taxonomy Taxonomy Browser (Bifidobacterium). Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=1678 (accessed on 13 July 2021).
- Lugli, G.A.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Turroni, F.; Viappiani, A.; van Sinderen, D.; Ventura, M. Tracking the Taxonomy of the Genus Bifidobacterium Based on a Phylogenomic Approach. Appl. Environ. Microbiol. 2018, 84, e02249–17. [Google Scholar] [CrossRef] [Green Version]
- Mattarelli, P.; Felis, G.E.; Pot, B.; Holzapfel, W.H.; Franz, C.M. International Committee on Systematics of Prokaryotes Subcommittee on the taxonomy of Bifidobacterium, Lactobacillus and related organisms Minutes of the closed meeting, 20 June 2019, Prague, Czech Republic. Int. J. Syst. Evol. Microbiol. 2020, 70, 2949–2951. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Longhi, G.; Ventura, M.; Van Sinderen, D.; Turroni, F. Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut. Microorganisms 2020, 9, 8. [Google Scholar] [CrossRef]
- Ventura, M.; Canchaya, C.; Tauch, A.; Chandra, G.; Fitzgerald, G.F.; Chater, K.F.; van Sinderen, D. Genomics of Actinobacteria: Tracing the Evolutionary History of an Ancient Phylum. Microbiol. Mol. Biol. Rev. 2007, 71, 495–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottacini, F.; Medini, D.; Pavesi, A.; Turroni, F.; Foroni, E.; Riley, D.; Giubellini, V.; Tettelin, H.; Van Sinderen, D.; Ventura, M. Comparative genomics of the genus Bifidobacterium. Microbiology 2010, 156, 3243–3254. [Google Scholar] [CrossRef] [Green Version]
- Turroni, F.; van Sinderen, D.; Ventura, M. Genomics and ecological overview of the genus Bifidobacterium. Int. J. Food Microbiol. 2011, 149, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhang, W.; Guo, C.; Yang, X.; Liu, W.; Wu, Y.; Song, Y.; Kwok, L.Y.; Cui, Y.; Menghe, B.; et al. Comparative Genomic Analysis of 45 Type Strains of the Genus Bifidobacterium: A Snapshot of Its Genetic Diversity and Evolution. PLoS ONE 2015, 10, e0117912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haukioja, A. Probiotics and Oral Health. Eur. J. Dent. 2010, 4, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radaic, A.; Kapila, Y.L. The oralome and its dysbiosis: New insights into oral microbiome-host interactions. Comput. Struct. Biotechnol. J. 2021, 19, 1335–1360. [Google Scholar] [CrossRef]
- Beighton, D.; Al-Haboubi, M.; Mantzourani, M.; Gilbert, S.; Clark, D.; Zoitopoulos, L.; Gallagher, J. Oral Bifidobacteria: Caries-associated bacteria in older adults. J. Dent. Res. 2010, 89, 970–974. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Brody, H.; Lin, G.-H.; Rangé, H.; Kuraji, R.; Ye, C.; Kamarajan, P.; Radaic, A.; Gao, L.; Kapila, Y. Probiotics, including nisin-based probiotics, improve clinical and microbial outcomes relevant to oral and systemic diseases. Periodontology 2000 2020, 82, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Biavati, B.; Mattarelli, P. Chapter 3—Related Genera Within the Family Bifidobacteriaceae. In The Bifidobacteria and Related Organisms; Mattarelli, P., Biavati, B., Holzapfel, W.H., Wood, B.J.B., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 49–66. ISBN 9780128050606. [Google Scholar]
- Lamendella, R.; Domingo, J.W.S.; Kelty, C.; Oerther, D.B. Bifidobacteria in Feces and Environmental Waters. Appl. Environ. Microbiol. 2008, 74, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Koropatkin, N.M.; Cameron, E.A.; Martens, E.C. How glycan metabolism shapes the human gut microbiota. Nat. Rev. Genet. 2012, 10, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Donaldson, G.; Mikulski, Z.; Boyajian, S.; Ley, K.; Mazmanian, S.K. Bacterial colonization factors control specificity and stability of the gut microbiota. Nat. Cell Biol. 2013, 501, 426–429. [Google Scholar] [CrossRef] [Green Version]
- Grimm, V.; Westermann, C.; Riedel, C.U. Bifidobacteria-Host Interactions—An Update on Colonisation Factors. BioMed Res. Int. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Dyakov, I.N.; Mavletova, D.A.; Chernyshova, I.N.; Snegireva, N.A.; Gavrilova, M.V.; Bushkova, K.K.; Dyachkova, M.S.; Alekseeva, M.G.; Danilenko, V.N. FN3 protein fragment containing two type III fibronectin domains from B. longum GT15 binds to human tumor necrosis factor alpha in vitro. Anaerobe 2020, 65, 102247. [Google Scholar] [CrossRef]
- Dyachkova, M.S.; Chekalin, E.V.; Danilenko, V.N. Positive Selection in Bifidobacterium Genes Drives Species-Specific Host–Bacteria Communication. Front. Microbiol. 2019, 10, 2374. [Google Scholar] [CrossRef] [Green Version]
- Lyte, M. Microbial Endocrinology in the Microbiome-Gut-Brain Axis: How Bacterial Production and Utilization of Neurochemicals Influence Behavior. PLoS Pathog. 2013, 9, e1003726. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Stilling, R.; Stanton, C.; Cryan, J. Collective unconscious: How gut microbes shape human behavior. J. Psychiatr. Res. 2015, 63, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelson, D.; Welsh, D.A.; Shellito, J.E. Regulation of lung immunity and host defense by the intestinal microbiota. Front. Microbiol. 2015, 6, 1085. [Google Scholar] [CrossRef] [Green Version]
- Kumar, H.; Collado, M.C.; Wopereis, H.; Salminen, S.; Knol, J.; Roeselers, G. The Bifidogenic Effect Revisited—Ecology and Health Per-spectives of Bifidobacterial Colonization in Early Life. Microorganisms 2020, 8, 1855. [Google Scholar] [CrossRef]
- Averina, O.V.; Ermolenko, E.I.; Ratushniy, A.Y.; Tarasova, E.; Borschev, Y.Y.; Leontieva, G.; Kramskaya, T.A.; Kotyleva, M.P.; Danilenko, V.N.; Suvorov, A.N. INFLUENCE OF PROBIOTICS ON CYTOKINE PRODUCTION IN THE IN VITRO AND IN VIVO SYSTEMS. Med. Immunol. Russ. 2015, 17, 443. [Google Scholar] [CrossRef]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Margolles, A.; Sánchez, B. Proteinaceous Molecules Mediating Bifidobacterium-Host Interactions. Front. Microbiol. 2016, 7, 1193. [Google Scholar] [CrossRef] [PubMed]
- Moriel, D.G.; Heras, B.; Paxman, J.J.; Lo, A.W.; Tan, L.; Sullivan, M.J.; Dando, S.J.; Beatson, S.A.; Ulett, G.C.; Schembri, M.A. Molecular and Structural Characterization of a Novel Escherichia coli Interleukin Receptor Mimic Protein. mBio 2016, 7, e02046–15. [Google Scholar] [CrossRef] [Green Version]
- Lehtoranta, L.; Latvala, S.; Lehtinen, M.J. Role of Probiotics in Stimulating the Immune System in Viral Respiratory Tract Infections: A Narrative Review. Nutrients 2020, 12, 3163. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santamarina, A.; Lamas, A.; del Carmen Mondragón, A.; Cardelle-Cobas, A.; Regal, P.; Rodriguez-Avila, J.A.; Miranda, J.M.; Franco, C.M.; Cepeda, A. Probiotic Effects against Virus Infections: New Weapons for an Old War. Foods 2021, 10, 130. [Google Scholar] [CrossRef]
- Parisa, A.; Roya, G.; Mahdi, R.; Shabnam, R.; Maryam, E.; Malihe, T. Anti-cancer effects of Bifidobacterium species in colon cancer cells and a mouse model of carcinogenesis. PLoS ONE 2020, 15, e0232930. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, S.; Wang, N.; Tan, H.-Y.; Zhang, Z.; Feng, Y. The Cross-Talk Between Gut Microbiota and Lungs in Common Lung Diseases. Front. Microbiol. 2020, 11, 301. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Makizaki, Y.; Oikawa, Y.; Tanaka, Y.; Maeda, A.; Shimakawa, M.; Komoto, S.; Moriguchi, K.; Ohno, H.; Tanigu-chi, K. Oral administration of Bifidobacterium bifidum G9-1 alleviates rotavirus gastroenteritis through regulation of intesti-nal homeostasis by inducing mucosal protective factors. PLoS ONE 2017, 12, e0173979. [Google Scholar] [CrossRef] [Green Version]
- Poluektova, E.; Yunes, R.; Danilenko, V. The Putative Antidepressant Mechanisms of Probiotic Bacteria: Relevant Genes and Proteins. Nutrients 2021, 13, 1591. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.J.; Shin, H.S. Antimicrobial and Immunomodulatory Effects of Bifidobacterium Strains: A Review. J. Microbiol. Biotechnol. 2020, 30, 1–10. [Google Scholar] [CrossRef]
- Akobeng, A.K.; Singh, P.; Kumar, M.; Al Khodor, S. Role of the gut microbiota in the pathogenesis of coeliac disease and potential therapeutic implications. Eur. J. Nutr. 2020, 59, 3369–3390. [Google Scholar] [CrossRef]
- Vaiserman, A.M.; Koliada, A.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef]
- Kato, K.; Odamaki, T.; Mitsuyama, E.; Sugahara, H.; Xiao, J.-Z.; Osawa, R. Age-Related Changes in the Composition of Gut Bifidobacterium Species. Curr. Microbiol. 2017, 74, 987–995. [Google Scholar] [CrossRef]
- Wong, C.; Sugahara, H.; Odamaki, T.; Xiao, J. Different physiological properties of human-residential and non-human-residential bifidobacteria in human health. Benef. Microbes 2018, 9, 111–122. [Google Scholar] [CrossRef]
- Turroni, F.; Milani, C.; Duranti, S.; Mahony, J.; van Sinderen, D.; Ventura, M. Glycan Utilization and Cross-Feeding Activities by Bifidobacteria. Trends Microbiol. 2018, 26, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Duranti, S.; Milani, C.; Lugli, G.A.; Mancabelli, L.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Sánchez, B.; Margolles, A.; et al. Evaluation of genetic diversity among strains of the human gut commensal Bifidobacterium adolescentis. Sci. Rep. 2016, 6, 23971. [Google Scholar] [CrossRef] [Green Version]
- Rabe, H.; Lundell, A.-C.; Sjöberg, F.; Ljung, A.; Strömbeck, A.; Gio-Batta, M.; Maglio, C.; Nordström, I.; Andersson, K.; Nookaew, I.; et al. Neonatal Gut Colonization by Bifidobacterium Is Associated with Higher Childhood Cytokine Responses. Gut Microbes 2020, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Luongo, D.; Coppola, A.; Treppiccione, L.; Bergamo, P.; Sorrentino, A.; Ferrocino, I.; Turroni, S.; Neviani, E.; Di Cagno, R.; Cocolin, L.; et al. Modulation of the cytokine profile in Caco-2 cells by faecal lactobacilli and bifidobacteria from individuals with distinct dietary habits. Cytokine 2017, 90, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, B.; Stanton, C.; Ross, R.P.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium pseudocatenulatum Ameliorates DSS-Induced Colitis by Maintaining Intestinal Mechanical Barrier, Blocking Proinflammatory Cytokines, Inhibiting TLR4/NF-κB Signaling, and Altering Gut Microbiota. J. Agric. Food Chem. 2021, 69, 1496–1512. [Google Scholar] [CrossRef] [PubMed]
- Dogra, S.K.; Doré, J.; Damak, S. Gut Microbiota Resilience: Definition, Link to Health and Strategies for Intervention. Front. Microbiol. 2020, 11, 572921. [Google Scholar] [CrossRef]
- Shang, J.; Wan, F.; Zhao, L.; Meng, X.; Li, B. Potential Immunomodulatory Activity of a Selected Strain Bifidobacterium bifidum H3-R2 as Evidenced in vitro and in Immunosuppressed Mice. Front. Microbiol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome–gut–brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol. Ther. 2016, 158, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Turroni, F.; Ventura, M.; Buttó, L.F.; Duranti, S.; O’Toole, P.; Motherway, M.O.; Van Sinderen, D. Molecular dialogue between the human gut microbiota and the host: A Lactobacillus and Bifidobacterium perspective. Cell. Mol. Life Sci. 2014, 71, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Wang, Y.; Xu, H.; Mei, X.; Yu, D.; Wang, Y.; Li, W. Antioxidant Properties of Probiotic Bacteria. Nutrients 2017, 9, 521. [Google Scholar] [CrossRef]
- Alí, A.; Boutjdir, M.; Aromolaran, A.S. Cardiolipotoxicity, Inflammation, and Arrhythmias: Role for Interleukin-6 Molecular Mechanisms. Front. Physiol. 2019, 9, 1866. [Google Scholar] [CrossRef]
- Högbom, M.; Ihalin, R. Functional and structural characteristics of bacterial proteins that bind host cytokines. Virulence 2017, 8, 1592–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nezametdinova, V.Z.; Mavletova, D.A.; Alekseeva, M.G.; Chekalina, M.S.; Zakharevich, N.V.; Danilenko, V.N. Species-Specific Ser-ine-Threonine Protein Kinase Pkb2 of Bifidobacterium Longum Subsp. Longum: Genetic Environment and Substrate Specificity. Anaerobe 2018, 51, 26–35. [Google Scholar] [CrossRef]
- Danilenko, V.N.; Alekseeva, M.G.; Mavletova, D.A.; Nezametdinova, V.Z.; Yunes, R.A. The PFNA Operon of Bifidobacteria and its Role in Adaptation to the Host. In Proceedings of the 8th International Human Microbiome Congress Ihmc 2021, Barcelona, Spain, 27–29 June 2021. [Google Scholar]
- Yunes, R.A.; Alekseeva, M.G.; Mavletova, D.A.; Nezametdinova, V.Z.; Danilenko, V.N. Role of PFNA Operon of Bifidobacteria in Species-specific Adaptation to Host’s Immune System. In Proceedings of the International Conference on Beneficial Microbes, RostovonDon, Russia, 1–6 June 2021. [Google Scholar]
- Nezametdinova, V.Z.; Zakharevich, N.V.; Alekseeva, M.G.; Averina, O.V.; Mavletova, D.A.; Danilenko, V.N. Identification and Characterization of the Serine/threonine Protein Kinases in Bifidobacterium. Arch. Microbiol. 2014, 196, 125–136. [Google Scholar] [CrossRef]
- Zakharevich, N.V.; Averina, O.V.; Klimina, K.M.; Kudryavtseva, A.V.; Kasianov, A.S.; Makeev, V.J.; Danilenko, V.N. Complete Genome Sequence of Bifidobacterium longum GT15: Identification and Characterization of Unique and Global Regulatory Genes. Microb. Ecol. 2015, 70, 819–834. [Google Scholar] [CrossRef]
- Alekseeva, M.G.; Mavletova, D.A.; Kolchina, N.V.; Nezametdinova, V.Z.; Danilenko, V.N. Isolation and Purification of Recombinant Serine/Threonine Protein Kinases of the Strain Bifidobacterium Longum B379M and Investigation of Their Activity. Biochemistry 2015, 80, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.diagrams.net/blog/open-source (accessed on 23 August 2021).
- Wang, L.-Q.; Zhao, F.; Liu, F.; Meng, X.-C. Live/dead state is not the factor influencing adhesion ability of Bifidobacterium animalis KLDS2.0603. J. Microbiol. 2013, 51, 584–589. [Google Scholar] [CrossRef]
- Kinoshita, H.; Ohuchi, S.; Arakawa, K.; Watanabe, M.; Kitazawa, H.; Saito, T. Isolation of lactic acid bacteria bound to the porcine intestinal mucosa and an analysis of their moonlighting adhesins. Biosci. Microbiota Food Health 2016, 35, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.M.; Enemark, E.J. Fundamental Characteristics of AAA+ Protein Family Structure and Function. Archaea 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Rodriguez, M.; De La Rosa, A.P.B.; Santos, L. Characterization of ATPase activity of the AAA ARC from Bifidobacterium longum subsp. infantis. Acta Biochim. Pol. 2015, 62, 221–227. [Google Scholar] [CrossRef]
- Wong, K.S.; Bhandari, V.; Janga, S.C.; Houry, W.A. The RavA-ViaA Chaperone-Like System Interacts with and Modulates the Activity of the Fumarate Reductase Respiratory Complex. J. Mol. Biol. 2017, 429, 324–344. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lee, G.I.; Van Doren, S.R.; Walker, J.C. The FHA Domain Mediates Phosphoprotein Interactions. J. Cell Sci. 2000, 113 Pt 23, 4143–4149. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, J.; Wang, N.; Cheng, Z.; Yang, R.; Lin, G.; Guo, Y.; Cai, D.; Xie, Y.; Zhao, X. Crystal structure of the second fibronectin type III (FN3) domain from human collagen α1 type XX. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2017, 73, 695–700. [Google Scholar] [CrossRef] [Green Version]
- SMART: FN3 Domain Annotation. Available online: http://smart.embl.de/smart/do_annotation.pl?DOMAIN=SM00060 (accessed on 13 July 2021).
- Hanamura, K.; Washburn, H.R.; Sheffler-Collins, S.I.; Xia, N.L.; Henderson, N.; Tillu, D.V.; Hassler, S.; Spellman, D.S.; Zhang, G.; Neubert, T.A.; et al. Extracellular phosphorylation of a receptor tyrosine kinase controls synaptic localization of NMDA receptors and regulates pathological pain. PLoS Biol. 2017, 15, e2002457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liongue, C.; Sertori, R.; Ward, A.C. Evolution of Cytokine Receptor Signaling. J. Immunol. 2016, 197, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-Y.; Yu, H.-W.; Cheng, C.-N.; Chen, J.-S.; Lin, C.-W.; Chen, P.-C.; Shieh, C.-C. A novel pathogenic mutation on Interleukin-7 receptor leading to severe combined immunodeficiency identified with newborn screening and whole exome sequencing. J. Microbiol. Immunol. Infect. 2020, 53, 99–105. [Google Scholar] [CrossRef]
- Siupka, P.; Hamming, O.T.; Kang, L.; Gad, H.H.; Hartmann, R. A conserved sugar bridge connected to the WSXWS motif has an important role for transport of IL-21R to the plasma membrane. Genes Immun. 2015, 16, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Szabó, R.; Láng, O.; Láng, J.; Illyés, E.E.; Kohidai, L.; Hudecz, F. Effect of SXWS/WSXWS peptides on chemotaxis and adhesion of the macrophage-like cell line J774. J. Mol. Recognit. 2015, 28, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Chaturvedi, N.; Rai, G. De novo modeling and structural characterization of IL9-IL9 receptor complex: A potential drug target for hematopoietic stem cell therapy. Netw. Model. Anal. Health Informatics Bioinform. 2020, 9, 1–10. [Google Scholar] [CrossRef]
- Chandler, P.G.; Buckle, A.M. Development and Differentiation in Monobodies Based on the Fibronectin Type 3 Domain. Cells 2020, 9, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovskaya, L.E.; Zlobinov, A.V.; Shingarova, L.N.; Boldyreva, E.F.; Gapizov, S.; Novototskaya-Vlasova, K.A.; Rivkina, E.M.; Dolgikh, D.A.; Kirpichnikov, M. Fusion with the cold-active esterase facilitates autotransporter-based surface display of the 10th human fibronectin domain in Escherichia coli. Extremophiles 2018, 22, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Shingarova, L.N.; Petrovskaya, L.E.; Zlobinov, A.V.; Gapizov, S.; Kryukova, E.A.; Birikh, K.R.; Boldyreva, E.F.; Yakimov, S.; Dolgikh, D.A.; Kirpichnikov, M.P. Construction of Artificial TNF-Binding Proteins Based on the 10th Human Fibronectin Type III Domain Using Bacterial Display. Biochem. Mosc. 2018, 83, 708–716. [Google Scholar] [CrossRef]
- Sirois, A.R.; Deny, D.A.; Baierl, S.R.; George, K.S.; Moore, S.J. Fn3 proteins engineered to recognize tumor biomarker mesothelin internalize upon binding. PLoS ONE 2018, 13, e0197029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, K.H.V.; Dao, T.K.; Nguyen, H.D.; Nguyen, T.Q.; Nguyen, T.T.; Truong, N.H.; Do, T.H. Some characters of bacterial cellulases in goats’ rumen elucidated by metagenomic DNA analysis and the role of fibronectin 3 module for endoglucanase function. Anim. Biosci. 2021, 34, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Mejía, L.; Valenzuela, S.V.; Frías-López, C.; Diaz, P.; Pastor, F.I.J. A bacterial GH6 cellobiohydrolase with a novel modular structure. Appl. Microbiol. Biotechnol. 2017, 101, 2943–2952. [Google Scholar] [CrossRef]
- Zakharevich, N.V.; Nezametdinova, V.Z.; Averina, O.V.; Chekalina, M.S.; Alekseeva, M.G.; Danilenko, V.N. Complete Genome Sequence of Bifidobacterium angulatum GT102: Potential Genes and Systems of Communication with Host. Russ. J. Genet. 2019, 55, 847–864. [Google Scholar] [CrossRef]
- Michelini, S.; Oki, K.; Yanokura, E.; Shimakawa, Y.; Modesto, M.; Mattarelli, P.; Biavati, B.; Watanabe, K. Bifidobacterium myosotis sp. nov., Bifidobacterium tissieri sp. nov. and Bifidobacterium hapali sp. nov., isolated from faeces of baby common marmosets (Callithrix jacchus L.). Int. J. Syst. Evol. Microbiol. 2016, 66, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Killer, J.; Kopečný, J.; Mrazek, J.; Rada, V.; Benada, O.; Koppová, I.; Havlik, J.; Straka, J. Bifidobacterium bombi sp. nov., from the bumblebee digestive tract. Int. J. Syst. Evol. Microbiol. 2009, 59, 2020–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delcenserie, V.; Gavini, F.; Beerens, H.; Tresse, O.; Franssen, C.; Daube, G. Description of a New Species, Bifidobacterium Crudilactis Sp. Nov., Isolated from Raw Milk and Raw Milk Cheeses. Syst. Appl. Microbiol. 2007, 30, 381–389. [Google Scholar] [CrossRef]
- Laureys, D.; Cnockaert, M.; De Vuyst, L.; Vandamme, P. Bifidobacterium Aquikefiri Sp. Nov., Isolated from Water Kefir. Int. J. Syst. Evol. Microbiol. 2016, 66, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Catanzaro, M.; Fagiani, F.; Racchi, M.; Corsini, E.; Govoni, S.; Lanni, C. Immune Response in COVID-19: Addressing a Pharmacological Challenge by Targeting Pathways Triggered by SARS-CoV-2. Signal Transduct Target 2020, 5, 84. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.S.; Quigley, E.M. The probiotic Bifidobacterium in the management of Coronavirus: A theoretical basis. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420961304. [Google Scholar] [CrossRef] [PubMed]
- Veselovsky, V.A.; Dyachkova, M.S.; Menyaylo, E.A.; Polyaeva, P.S.; Olekhnovich, E.I.; Shitikov, E.A.; Bespiatykh, D.A.; Semashko, T.A.; Kasianov, A.S.; Ilina, E.N.; et al. Gene Networks Underlying the Resistance of Bifidobacterium longum to Inflammatory Factors. Front. Immunol. 2020, 11, 595877. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, T.; Matsushima-Nishiwaki, R.; Yasuda, E.; Matsuura, J.; Toyoda, H.; Kaneoka, Y.; Kumada, T.; Kozawa, O. Heat shock protein 20 (HSPB6) regulates TNF-α-induced intracellular signaling pathway in human hepatocellular carcinoma cells. Arch. Biochem. Biophys. 2014, 565, 1–8. [Google Scholar] [CrossRef]
- Yoon, Y.H.; Byun, J.R. Occurrence of Glutathione Sulphydryl (GSH) and Antioxidant Activities in Probiotic Lactobacillus spp. Asian Australas. J. Anim. Sci. 2004, 17, 1582–1585. [Google Scholar] [CrossRef]
- Neumann, W.; Gulati, A.; Nolan, E.M. Metal homeostasis in infectious disease: Recent advances in bacterial metallophores and the human metal-withholding response. Curr. Opin. Chem. Biol. 2017, 37, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects From Hypertensive Cardiovascular Damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Copaescu, A.; James, F.; Mouhtouris, E.; Vogrin, S.; Smibert, O.C.; Gordon, C.L.; Drewett, G.; Holmes, N.E.; Trubiano, J.A. The Role of Immunological and Clinical Biomarkers to Predict Clinical COVID-19 Severity and Response to Therapy—A Prospective Longitudinal Study. Front. Immunol. 2021, 12, 646095. [Google Scholar] [CrossRef]
- Karki, R.; Sharma, B.R.; Tuladhar, S.; Williams, E.P.; Zalduondo, L.; Samir, P.; Zheng, M.; Sundaram, B.; Banoth, B.; Malireddi, R.K.S.; et al. Synergism of TNF-α and IFN-γ Triggers Inflammatory Cell Death, Tissue Damage, and Mortality in SARS-CoV-2 Infection and Cytokine Shock Syndromes. Cell 2021, 184, 149–168.e17. [Google Scholar] [CrossRef]
- Copaescu, A.; Smibert, O.; Gibson, A.; Phillips, E.J.; Trubiano, J.A. The Role of IL-6 and Other Mediators in the Cytokine Storm Associated with SARS-CoV-2 Infection. J. Allergy Clin. Immunol. 2020, 146, 518–534.e1. [Google Scholar] [CrossRef]
- Mopuru, R.; Chaturvedi, S.; Burkholder, B.M. Relapsing Thrombotic Thrombocytopenic Purpura (TTP) in a Patient Treated with Infliximab for Chronic Uveitis. Ocul. Immunol. Inflamm. 2020, 1–3. [Google Scholar] [CrossRef]
- Fontes, J.; Rose, N.R.; Čiháková, D. The varying faces of IL-6: From cardiac protection to cardiac failure. Cytokine 2015, 74, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Zegeye, M.M.; Lindkvist, M.; Fälker, K.; Kumawat, A.K.; Paramel, G.; Grenegård, M.; Sirsjö, A.; Ljungberg, L.U. Activation of the JAK/STAT3 and PI3K/AKT pathways are crucial for IL-6 trans-signaling-mediated pro-inflammatory response in human vascular endothelial cells. Cell Commun. Signal. 2018, 16, 55. [Google Scholar] [CrossRef] [PubMed]
- Timper, K.; Denson, J.L.; Steculorum, S.; Heilinger, C.; Ruud, L.E.; Wunderlich, C.M.; Rose-John, S.; Wunderlich, F.T.; Brüning, J.C. IL-6 Improves Energy and Glucose Homeostasis in Obesity via Enhanced Central IL-6 trans -Signaling. Cell Rep. 2017, 19, 267–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzani, A.; Viviani, B. Neuromodulatory properties of inflammatory cytokines and their impact on neuronal excitability. Neuropharmacology 2015, 96, 70–82. [Google Scholar] [CrossRef]
- Probert, L. TNF and its receptors in the CNS: The essential, the desirable and the deleterious effects. Neuroscience 2015, 302, 2–22. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Wang, X.; Li, Y.; Xu, L.; Yu, X.; Ge, L.; Li, J.; Zhu, Y.; He, S. Necroptosis Mediates TNF-Induced Toxicity of Hippocampal Neurons. BioMed Res. Int. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Maier, O.; Siegemund, M.; Wajant, H.; Scheurich, P.; Pfizenmaier, K. A TNF Receptor 2 Selective Agonist Rescues Human Neurons from Oxidative Stress-Induced Cell Death. PLoS ONE 2011, 6, e27621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veroni, C.; Gabriele, L.; Canini, I.; Castiello, L.; Coccia, E.M.; Remoli, M.E.; Columba-Cabezas, S.; Aricò, E.; Aloisi, F.; Agresti, C. Activation of TNF receptor 2 in microglia promotes induction of anti-inflammatory pathways. Mol. Cell. Neurosci. 2010, 45, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Fredj, S.; Bescond, J.; Louault, C.; Delwail, A.; Lecron, J.-C.; Potreau, D. Role of interleukin-6 in cardiomyocyte/cardiac fibroblast interactions during myocyte hypertrophy and fibroblast proliferation. J. Cell. Physiol. 2005, 204, 428–436. [Google Scholar] [CrossRef]
- Alessandri, G.; Ossiprandi, M.C.; Mac Sharry, J.; Van Sinderen, D.; Ventura, M. Bifidobacterial Dialogue With Its Human Host and Consequent Modulation of the Immune System. Front. Immunol. 2019, 10, 2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanmugam, G.; Narasimhan, M.; Sakthivel, R.; Kumar, R.R.; Davidson, C.; Palaniappan, S.; Claycomb, W.W.; Hoidal, J.R.; Darley-Usmar, V.M.; Rajasekaranacg, N.S.; et al. A Biphasic Effect of TNF-α in Regulation of the Keap1/Nrf2 Pathway in Cardiomyocytes. Redox Biol. 2016, 9, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.; Li, N.; Dobrev, D. Role of inflammatory signaling in atrial fibrillation. Int. J. Cardiol. 2019, 287, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Dams-Kozlowska, H.; Gryska, K.; Kwiatkowska-Borowczyk, E.; Izycki, D.; Rose-John, S.; Mackiewicz, A. A designer hyper interleukin 11 (H11) is a biologically active cytokine. BMC Biotechnol. 2012, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Lan, S.-H.; Lai, C.-C.; Huang, H.-T.; Chang, S.-P.; Lu, L.-C.; Hsueh, P.-R. Tocilizumab for severe COVID-19: A systematic review and meta-analysis. Int. J. Antimicrob. Agents 2020, 56, 106103. [Google Scholar] [CrossRef]
- Gremese, E.; Cingolani, A.; Bosello, S.L.; Alivernini, S.; Tolusso, B.; Perniola, S.; Landi, F.; Pompili, M.; Murri, R.; Santoliquido, A.; et al. Sarilumab Use in Severe SARS-CoV-2 Pneumonia. EClinicalMedicine 2020, 27, 100553. [Google Scholar] [CrossRef]
- Ghosn, L.; Chaimani, A.; Evrenoglou, T.; Davidson, M.; Graña, C.; Schmucker, C.; Bollig, C.; Henschke, N.; Sguassero, Y.; Nejstgaard, C.H.; et al. Interleukin-6 blocking agents for treating COVID-19: A living systematic review. Cochrane Database Syst. Rev. 2021, 2021, CD013881. [Google Scholar] [CrossRef]
- Chen, F.; Teachey, D.; Pequignot, E.; Frey, N.; Porter, D.; Maude, S.L.; Grupp, S.A.; June, C.H.; Melenhorst, J.J.; Lacey, S.F. Measuring IL-6 and sIL-6R in serum from patients treated with tocilizumab and/or siltuximab following CAR T cell therapy. J. Immunol. Methods 2016, 434, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Palanques-Pastor, T.; López-Briz, E.; Poveda Andrés, J.L. Involvement of Interleukin 6 in SARS-CoV-2 Infection: Siltuximab as a Therapeutic Option against COVID-19. Eur. J. Hosp. Pharm. Sci. Pract. 2020, 27, 297–298. [Google Scholar] [CrossRef]
- Bezzio, C.; Manes, G.; Bini, F.; Pellegrini, L.; Saibeni, S. Infliximab for Severe Ulcerative Colitis and Subsequent SARS-CoV-2 Pneumonia: A Stone for Two Birds. Gut 2021, 70, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.A.; Goodhand, J.R.; Bewshea, C.; Nice, R.; Chee, D.; Lin, S.; Chanchlani, N.; Butterworth, J.; Cooney, R.; Croft, N.M.; et al. Anti-SARS-CoV-2 Antibody Responses Are Attenuated in Patients with IBD Treated with Infliximab. Gut 2021, 70, 865–875. [Google Scholar] [CrossRef]
- Meredith, J.; Khedim, C.-A.; Henderson, P.; Wilson, D.C.; Russell, R.K. Paediatric Inflammatory Multisystem Syndrome Temporally Associated with SARS-CoV-2 [PIMS-TS] in a Patient Receiving Infliximab Therapy for Inflammatory Bowel Disease. J. Crohns. Colitis 2021, 15, 687–691. [Google Scholar] [CrossRef]
- Robinson, P.C.; Liew, D.F.; Liew, J.W.; Monaco, C.; Richards, D.; Shivakumar, S.; Tanner, H.L.; Feldmann, M. The Potential for Repurposing Anti-TNF as a Therapy for the Treatment of COVID-19. Med 2020, 1, 90–102. [Google Scholar] [CrossRef]
- Turkmen, M.; Dogan, S. Certolizumab pegol in the treatment of psoriasis: Real-life data. Dermatol. Ther. 2021, 34, e14929. [Google Scholar] [CrossRef]
- Bossa, F.; Carparelli, S.; Latiano, A.; Palmieri, O.; Tavano, F.; Panza, A.; Pastore, M.; Marseglia, A.; D’Altilia, M.; Latiano, T.; et al. Impact of the COVID-19 Outbreak and the Serum Prevalence of SARS-CoV-2 Antibodies in Patients with Inflammatory Bowel Disease Treated with Biologic Drugs. Dig. Liver Dis. 2021, 53, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Hariharan, A.; Hakeem, A.R.; Radhakrishnan, S.; Reddy, M.S.; Rela, M. The Role and Therapeutic Potential of NF-kappa-B Pathway in Severe COVID-19 Patients. Inflammopharmacology 2021, 29, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hansel, T.T.; Kropshofer, H.; Singer, T.; Mitchell, J.A.; George, A. The safety and side effects of monoclonal antibodies. Nat. Rev. Drug Discov. 2010, 9, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Richerson, P.J.; Gavrilets, S.; de Waal, F.B.M. Modern theories of human evolution foreshadowed by Darwin’s Descent of Man. Science 2021, 372, eaba3776. [Google Scholar] [CrossRef] [PubMed]






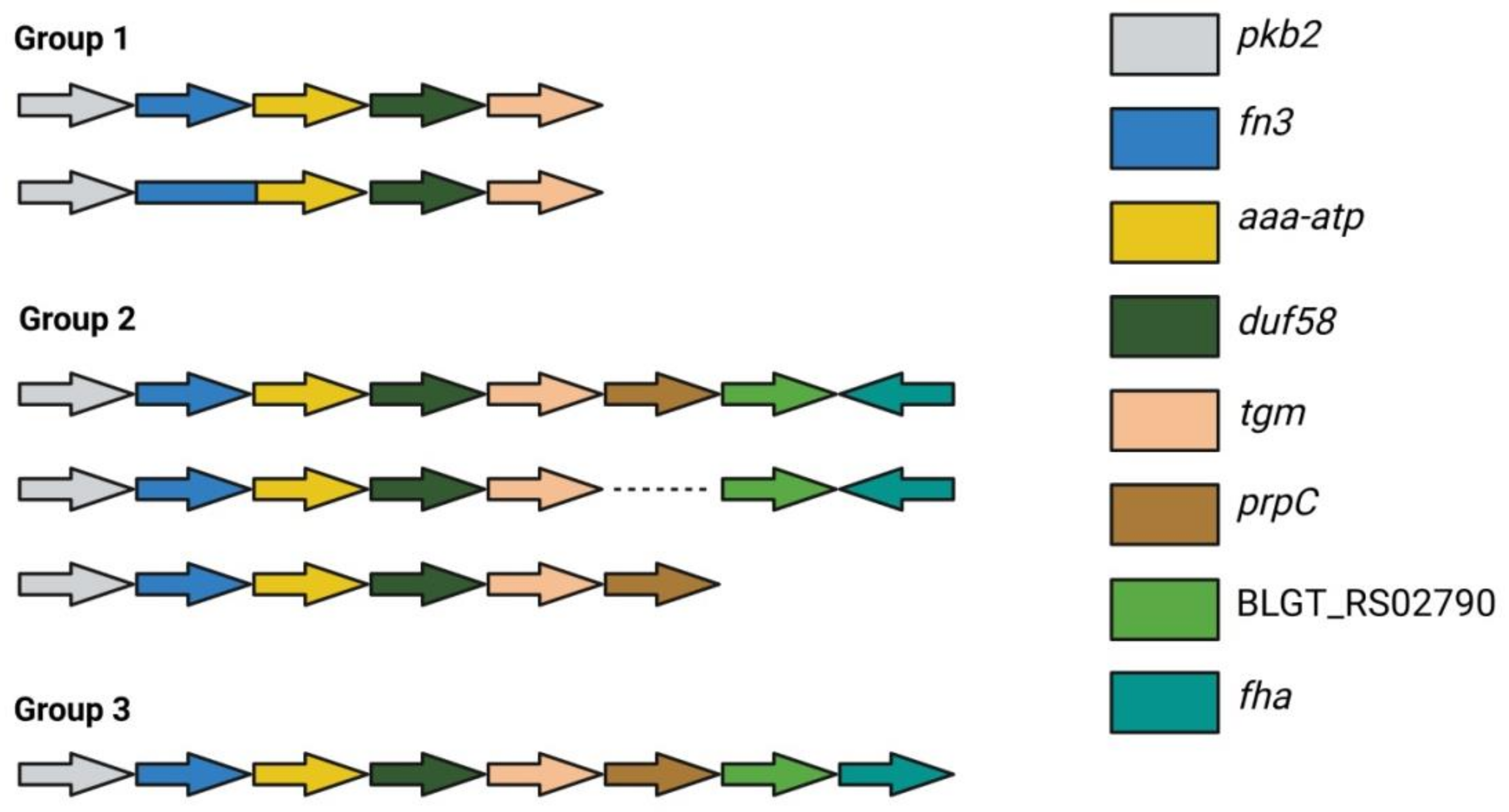{kind=link}
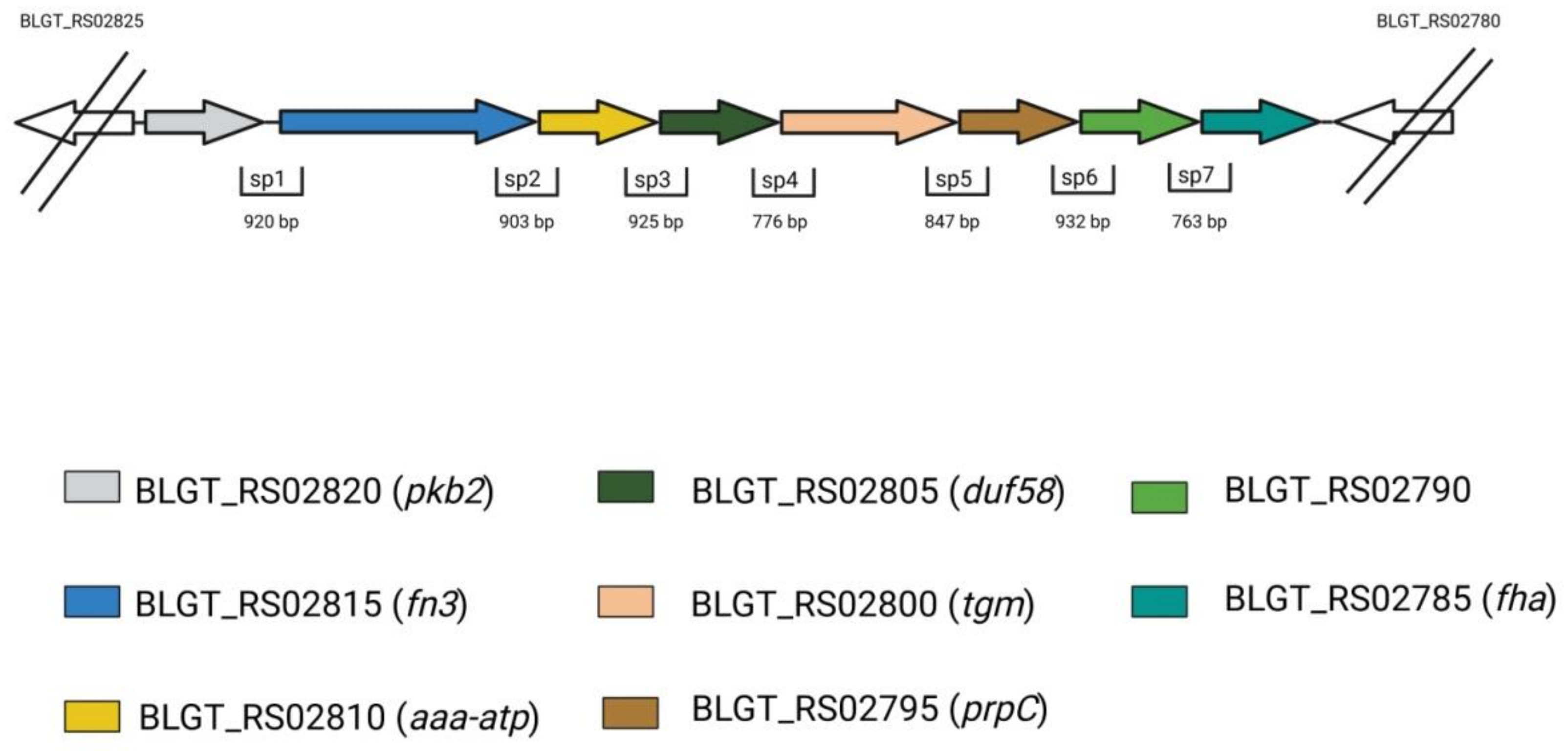{kind=link}
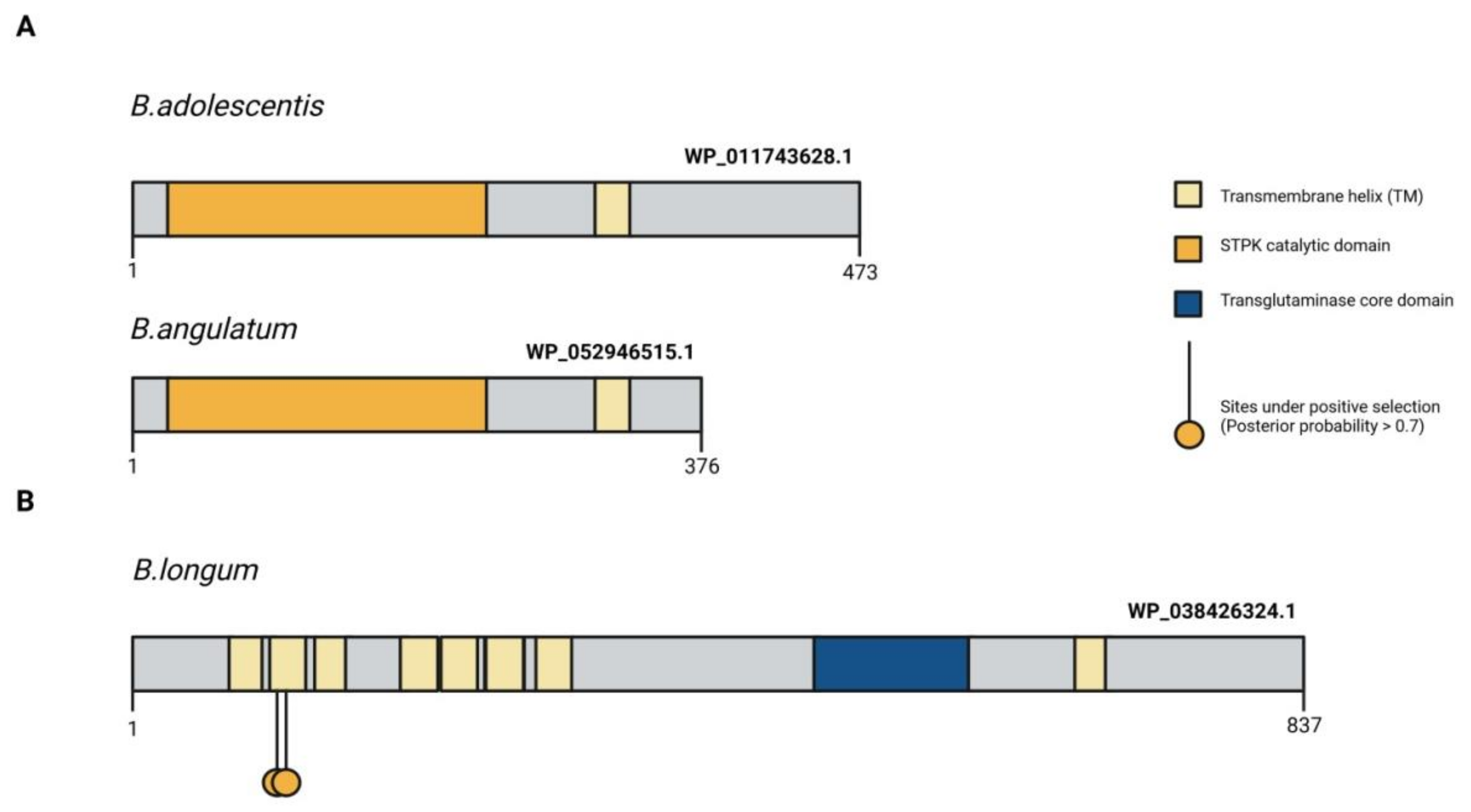{kind=link}
{kind=link}
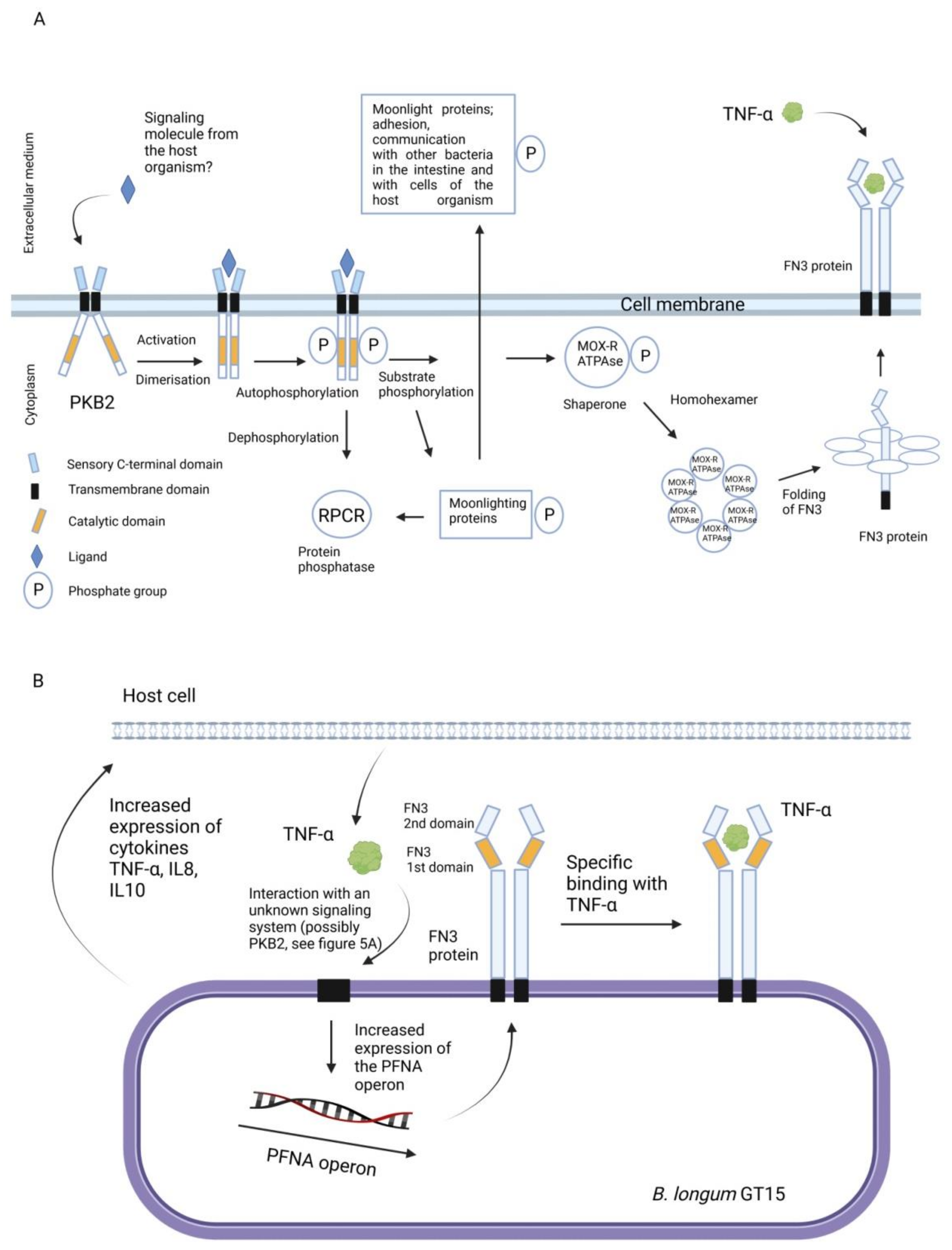{kind=link}
| Species | Source of Isolation | Motifs of Cytokine Receptors | Class | Phylogenetic Group According to [8] | |
|---|---|---|---|---|---|
| 1st FN3 Domain | 2nd FN3 Domain | ||||
| B. adolescentis | Human | WS-DS | WS-PS | I | B. adolescentis |
| B. catenulatum_catenulatum | Human | WS-ES | WS-PS | I | |
| B. catenulatum_kashiwanohense | Human | WS-ES | WS-PS | I | |
| B. dentium | Human | WS-PS | WS-PS | I | |
| B. moukalabense | Monkey | WS-PS | WS-PS | I | |
| B. pseudocatenulatum | Human | WS-ES | WS-PS | I | |
| B. ruminantium | Cow | WS-ES | WS-PS | I | |
| B. boum | Cow | WS-ES | WS-PS | I | B. boum |
| B. porcinum | Pig | WS-ES | WS-PS | I | |
| B. thermacidophilum | Soy whey | WS-ES | WS-PS | I | |
| B. thermophilum | Pig | WS-ES | WS-PS | I | |
| B. pullorum_gallinarum | Chicken | Both the PFNA cluster and the gene encoding FN3 are absent | B. pullorum | ||
| B. pullorum_ saeculare | Rabbit | ||||
| B. pullorum_ pullorum | Chicken | ||||
| B. asteroides | Bee | WS-AS | SG-VA | II | B. asteroides |
| B. actinocoloniiforme | Bumblebee | WS-PS | SG-AA | II | |
| B. coryneforme | Bee | WS-AS | AG-AR | III | |
| B. indicum | Insect | WS-AS | AG-AR | III | |
| B. longum_infantis | Human | WS-PS | WS-ES | I | B. longum |
| B. longum_longum | Human | WS-PS | WS-ES | I | |
| B. longum_suis | Pig | WS-PS | WS-ES | I | |
| B. breve | Human | WS-AS | WS-ES | I | |
| B. aesculapii | Monkey | WS-ES | DG-SA | IV | |
| B. stellenboschense | Monkey | WS-DS | DG-GA | IV | |
| B. angulatum | Human | WS-YS | SG-QA | II | |
| B. merycicum | Cow | WS-YS | SG-QA | II | |
| B. psychraerophilum | Pig | The gene encoding FN3 is absent | B. psychraerophilum | ||
| B. aquikefiri | Kefir | ||||
| B. crudilactis | Milk | ||||
| B. bifidum | Human | WS-PS | EG-PS | I | B. bifidum |
| B. biavatii | Monkey | WS-PS | VG-HG | V | |
| B. scardovii | Blood | WS-PS | DG-PG | IV | |
| B. animalis_animalis | Rat | WS-DS | AS-PS | III | B. pseudolongum |
| B. animalis_lactis | Milk | WS-DS | AS-PS | III | |
| B. pseudolongum_pseudolongum | Pig | WS-SS | TG-PS | VI | |
| B. pseudolongum_globosum | Cow | WS-SS | TG-PS | VI | |
| B. choerinum | Pig | WS-SS | Domain absent | VII | |
| B. cuniculi | Rabbit | DS-WS | Domain absent | VII | |
| B. gallicum | Human | VS-PS | Domain absent | VIII | |
| B. bombi | Bumblebee | WS-PS | DG-VS | IV | B. bombi |
| B. commune | Bumblebee | WS-PS | DG-VS | IV | |
| B. tissieri | Marmoset monkey | WS-PS | DG-EA | IV | B. tissieri |
| B. vansinderenii | Bare-faced marmoset | WS-PS | DG-EA | IV | |
| B. catulorum | Marmoset monkey | WS-PS | DG-EG | IV | |
| B. primatium | Bare-faced marmoset | WS-PS | DG-EG | IV | |
| Name of the Drug | Target | Origin | Target Disease | Other Trade Names | Results of Anti-COVID-19 Trials | References |
|---|---|---|---|---|---|---|
| Tocilizumab | IL6R | Recombinant humanized monoclonal antibody | CRS Rheumatoid arthritis Giant cell arteritis Juvenile idiopathic arthritis CAR T cell induced cytokine storm | Actemra RoActemra | Insufficient evidence of efficacy | [111] |
| Sarilumab | IL6R | Human IgG1 monoclonal antibody | Rheumatoid arthritis | Kevzara | Unknown, preliminary results are positive | [112] |
| Левилимаб/Levilimab | IL6R | Human monoclonal antibody | Rheumatoid arthritis | Ilsira (Biocad) | Unknown, approved for clinical trials | [113] |
| Siltuximab | IL6 | Human–murine chimeric monoclonal antibody | Multicentric Castleman disease; CAR T cell induced cytokine storm | Sylvant | Unknown, a candidate | [114,115] |
| Infliximab | TNFα | Chimeric murine/human IgG1 | Rheumatoid arthritis, ankylosing spondylitis, Crohn’s disease and ulcerative colitis, psoriasis and psoriatic arthritis | Remicade | Unknown, undergoing clinical trials (1) a positive effect; (2) mild formation of antibodies against the virus is observed; inflammation and multisystem dysfunction | [116,117,118,119] |
| Adalimumab | TNFα | Fully Human IgG1 | Autoimmune inflammatory diseases, including rheumatoid arthritis, Crohn’s disease, and psoriatic arthritis | Humira | Clinical trials are scheduled | [119] |
| Certolizumab-pegol | TNFα | Humanized, PEGylated Fab | Autoimmune inflammatory diseases, including rheumatoid arthritis, Crohn’s disease, and psoriatic arthritis | Cimzia | Unknown, preliminary studies suggest lack of efficacy as well as risk factors | [120] |
| Golimumab | TNFα | Fully Human IgG1 | Autoimmune inflammatory diseases, including rheumatoid arthritis, Crohn’s disease, and psoriatic arthritis | Simponi | Unknown, preliminary studies suggest lack of efficacy as well as risk factors | [121] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nezametdinova, V.Z.; Yunes, R.A.; Dukhinova, M.S.; Alekseeva, M.G.; Danilenko, V.N. The Role of the PFNA Operon of Bifidobacteria in the Recognition of Host’s Immune Signals: Prospects for the Use of the FN3 Protein in the Treatment of COVID-19. Int. J. Mol. Sci. 2021, 22, 9219. https://doi.org/10.3390/ijms22179219
Nezametdinova VZ, Yunes RA, Dukhinova MS, Alekseeva MG, Danilenko VN. The Role of the PFNA Operon of Bifidobacteria in the Recognition of Host’s Immune Signals: Prospects for the Use of the FN3 Protein in the Treatment of COVID-19. International Journal of Molecular Sciences. 2021; 22(17):9219. https://doi.org/10.3390/ijms22179219
Chicago/Turabian StyleNezametdinova, Venera Z., Roman A. Yunes, Marina S. Dukhinova, Maria G. Alekseeva, and Valery N. Danilenko. 2021. "The Role of the PFNA Operon of Bifidobacteria in the Recognition of Host’s Immune Signals: Prospects for the Use of the FN3 Protein in the Treatment of COVID-19" International Journal of Molecular Sciences 22, no. 17: 9219. https://doi.org/10.3390/ijms22179219