
Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach


1
Department of Molecular Science and Technology, Ajou University, Suwon 16499, Korea
2
S&K Therapeutics, Ajou University Campus Plaza 418, 199 Worldcup-ro, Yeongtong-gu, Suwon 16502, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(18), 10716; https://doi.org/10.3390/ijms231810716
Submission received: 20 August 2022
/
Revised: 10 September 2022
/
Accepted: 10 September 2022
/
Published: 14 September 2022
(This article belongs to the Special Issue The Role of Toll-Like Receptors (TLR) in Infection and Inflammation)
Abstract
:The innate immune system facilitates defense mechanisms against pathogen invasion and cell damage. Toll-like receptors (TLRs) assist in the activation of the innate immune system by binding to pathogenic ligands. This leads to the generation of intracellular signaling cascades including the biosynthesis of molecular mediators. TLRs on cell membranes are adept at recognizing viral components. Viruses can modulate the innate immune response with the help of proteins and RNAs that downregulate or upregulate the expression of various TLRs. In the case of COVID-19, molecular modulators such as type 1 interferons interfere with signaling pathways in the host cells, leading to an inflammatory response. Coronaviruses are responsible for an enhanced immune signature of inflammatory chemokines and cytokines. TLRs have been employed as therapeutic agents in viral infections as numerous antiviral Food and Drug Administration-approved drugs are TLR agonists. This review highlights the therapeutic approaches associated with SARS-CoV-2 and the TLRs involved in COVID-19 infection.
1. Introduction
Toll-like receptors (TLRs) are central mediators of the innate and adaptive immune responses. The immune system exhibits a defense mechanism for the host against pathogenic materials (exogenous and/or endogenous) at the cellular level [1]. Pattern recognition receptors (PRRs) including DNA sensors, RIG-1-like receptors, and TLRs are part of the innate immune system that protects against microbial infection. PRRs recognize conserved pathogen-associated molecular patterns (PAMPs) from microbes and endogenous danger-associated molecular patterns (DAMPs) produced by necrotic cells [2]. PAMPs are derived from viral, bacterial, parasitic, and fungal pathogens. The chemical nature of PAMPs recognized by TLRs varies greatly among organisms. In phylogenetics, TLRs are considered the most ancient class of PRRs. A large number of TLRs have been reported across a wide range of vertebrate and invertebrate species. The signaling pathways and adaptor proteins related to TLRs are evolutionary conserved, from Porifera to mammals. Moreover, similar domain patterns can be observed in most TLR homologs [3,4].
Viruses are responsible for initiating innate immunity through TLRs. Viruses, via a combination of small and unique proteins, not only escape the innate immune system but also destabilize the paybacks of the virus [5]. Similar to other pathogens, viruses are sensed by TLRs. Some viruses encode unique proteins that target TLR signaling. The hepatitis C virus encodes proteins that inhibit TLR-mediated signaling such as NS5A and protease NS3/4A [6,7], which inhibits MyD88 and cleaves TIR-domain-containing adapter-inducing interferon-β (TRIF), respectively. Moreover, the two vaccinia virus proteins have been reported as inhibitors of the TLR system; for example, A52R was observed to inhibit TLR-mediated NF-κB activation by targeting IRAK2 [8], whereas A46R exhibited a connection with TLR signaling downregulation by employing Toll-interleukin-1 receptor (TIR) domain-containing adaptors [9]. Intracellular TLRs not only sense viral and bacterial nucleic acids, but also identify self-nucleic acids in cellular abnormalities such as autoimmunity [10].
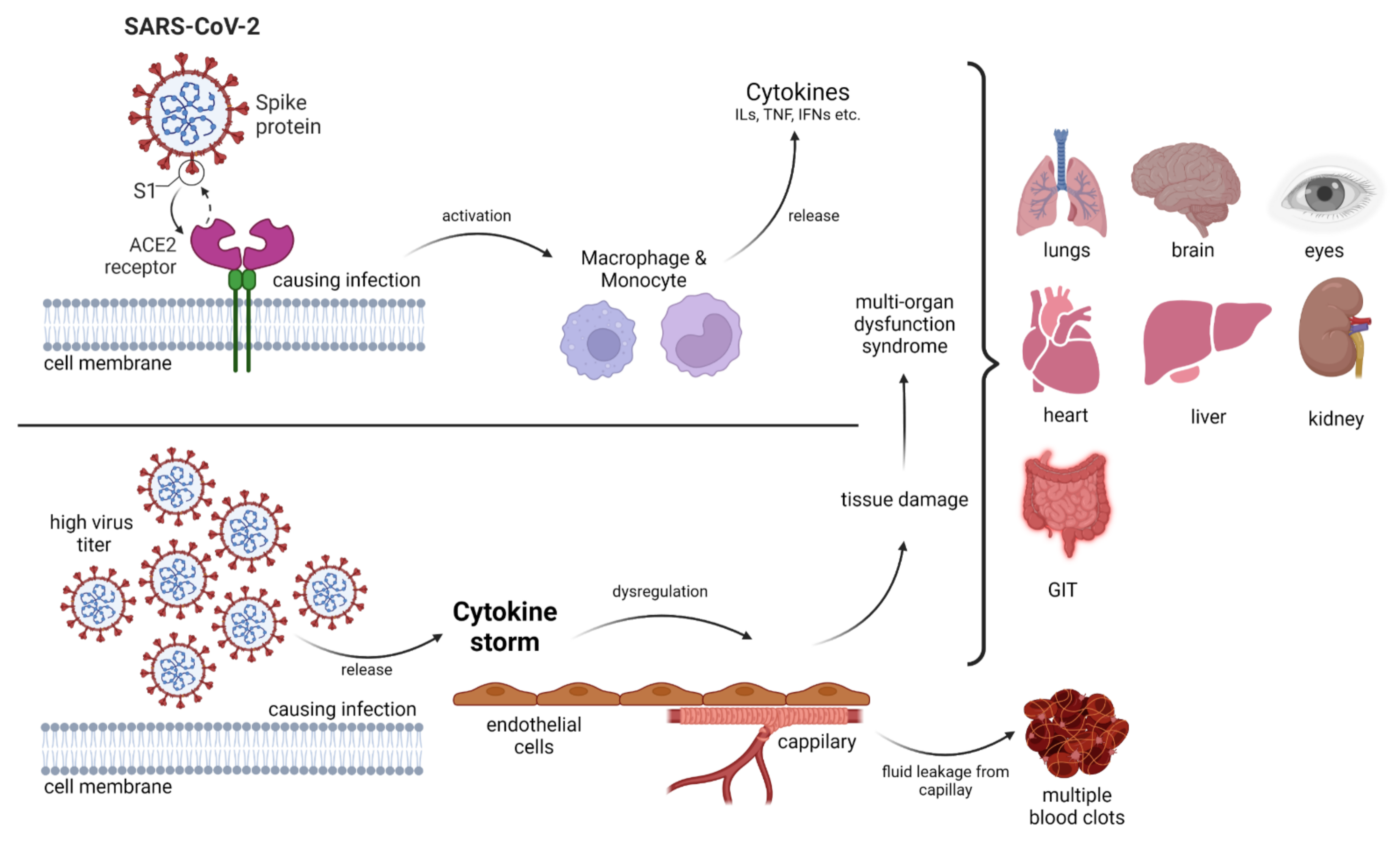
A novel single-stranded RNA (ssRNA)-containing virus causes coronavirus disease (COVID-19), also referred to as severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), which became a pandemic after the first case was identified in Wuhan, China in December 2019. With the spread of COVID-19, the pandemic poses a global challenge [11,12]. From a clinical point of view, the virus has various manifestations ranging from patients becoming critically ill with acute respiratory distress syndrome to asymptomatic infection. In the intensive care unit, multiorgan support therapy has been essential in almost every case of COVID-19 (Figure 1). The critical disease stage is typically observed at 7–10 days of clinical infection [11,13]. Hyperinflammatory outcomes (cytokine storm) are mainly associated with clinical impediments and mortality [14]. A possible treatment methodology in the form of vaccines is being employed for the prevention of SARS-CoV-2 infection, but there is no operative therapeutic treatment option available. Consequently, exploring new drug targets is necessary. One of the most important molecular targets is TLRs. The interaction of the SARS-CoV-2 spike glycoprotein with TLR and the enhanced expression of genes associated with TLR signaling could indicate the possible involvement of these tiny molecular machines and their inflammatory cascades [14].
Structurally, TLRs are type I transmembrane (TM) proteins with three distinct domains including an extracellular domain (ectodomain) that contains tandem copies of leucine-rich repeats, a single-pass TM as well as a cytoplasmic TIR downstream-signaling domain. TLRs experience either homodimerization or heterodimerization when encountering PAMPs and/or DAMPs, and adaptor proteins are employed; subsequently, a complex cellular event of downstream signal transduction is initiated, leading to the expression of inflammatory cytokines and interferons (IFN) that is observable at the molecular level [2]. The underlying TLR signaling cascades have been elucidated using structural, genetic, biochemical, and in silico methodologies [15].
Downstream signaling is made possible by the presence of cytosolic TIR domain-adaptor proteins such as TRIF (also known as TICAM1), TRAM (TICAM2), MyD88, and MAL [5,16]. The involvement of TLRs with TIR adaptors leads to the activation of cytosolic signaling complexes including IRAK and TRAF proteins. These entities are responsible for the activation of transcription factors such as IRF and NF-κB. This executes the synthesis of type I IFNs and proinflammatory cytokines [16]. IRF7 is essential for IFN-α synthesis, NF-κB is necessary for TNF and IL-6 induction, and IRF3 and NF-κB are required for IFN-β production [17].
By neutralizing internal and/or external threats administered by TLRs, the innate immune system makes defensive contributions to the survival of the host biological system. However, dysregulation and/or overactivation of this system leads to various disorders such as inflammation, cancer, and autoimmunity [18,19,20].
2. Structure of Coronavirus
SARS-CoV-2, a member of the β-coronavirus genus in the family Coronaviridae, has an envelope and positive-sense ssRNA genome of 29,891 nucleotides, encoding circular nucleocapsid proteins with 9860 amino acid residues [21]. The viral particle size ranges from 80 to 220 nm. Overall, 10 open reading frames (ORFs) have been identified in its genome to date (approximately 26–32 kb). The first ORF (almost 2/3 of the viral RNA) encodes polyproteins 1a (ORF1a) and 1b (ORF1b) [22]. Furthermore, these ORFs are cleaved by proteases into 16 nonstructural proteins (NSPs) that are responsible for genome replication and transcription [23]. Structural proteins (SPs) are encoded by the remaining ORFs [24,25]. The main SPs and NSPs of SARS-CoV-2 are summarized in Table 1 and Table 2, respectively. The name coronavirus is derived from the appearance under the electron microscope, in which the presence of crown-like spikes on the envelope resembles the corona of the sun [26]. SPs form the viral envelope that holds the RNA genome, while NSPs are expressed in host-infected cells but are not incorporated into virion infectious particles. These NSPs include various transcription factors and enzymes such as RNA-dependent RNA polymerase (RdRp) and hemagglutinin esterase (HE). Moreover, the virion employs enzymes such as RNA replicases and viral proteases to replicate itself [22,27,28,29].
Various SPs have been identified including the glycoprotein membrane (M), spike (S), small envelope (E), and nucleoprotein (N), and other accessory proteins. M-glycoprotein is the most abundant, spanning the membrane bilayer thrice [30]. S-glycoprotein (150 kDa) is a type-I TM protein on the outer surface of the virus and is responsible for the binding of the virus to host cell receptors (ACE2). The S protein amino acid sequence of SARS-CoV-2 exhibits 86% similarity to that of SARS-CoV [31]. The S protein consists of oligosaccharides bound to serine amino acids through o-glycosides. The three major segments of S protein are the ectodomain, TM, and intracellular regions. The intracellular domain comprises the membrane fusion subunit S2 (trimeric stalk) as well as a short tail part known as the receptor-binding S1 domain (RBD; three S1 heads) [32,33]. Protein–protein interaction (PPI) between the human ACE2 and SARS-CoV-2 S protein facilitates viral attachment as well as the cellular entry of coronaviruses; thus, small-molecule blockage of these PPIs is a more inspiring therapeutic approach than inhibition via antibodies [34]. The S1 subunit of the S protein enables ACE2-mediated virus attachment, whereas the S2 subunit facilitates membrane fusion. Specifically, asparagine, glutamine, serine, phenylalanine, and leucine residues present in the S protein boost ACE2 binding [35].
Moreover, N protein bound to nucleic acids is an important structural component of the virus, which is responsible for viral replication and cellular response to infection in the host cellular machinery [31] (Table 1). The N protein comprises a serine-rich linker region sandwiched between the N-terminal domain (NTD) and the C-terminal domain (CTD). These termini are crucial for viral entry and processing in host cells. The CTD regulates nucleocapsid formation and the NTD adheres to the viral genome in the form of orthorhombic crystals. Phosphorylation sites are also present in the linker region, which control its function [35]. In the case of SARS-CoV, the N protein enhances the activation of cyclooxygenase-2 (COX-2), resulting in the inflammation of pulmonary cells [36]. Moreover, the N protein interacts with the p42 proteasome subunit, which degrades the virion [37]. This also disables type-I IFN, which is responsible for suppressing the host immune responses produced by biological systems against viral infections [38]. The interaction of the N protein with heterogeneous nuclear ribonucleoproteins leads to increased viral RNA synthesis [39]. The N protein sequence of SARS-CoV-2 shows a 94.3% similarity to that of the SARS-CoV [31].
The smallest TM structural protein in coronaviruses is the E protein (Table 1), which comprises two different domains: the NTD (1–9 residues) as well as a hydrophobic domain (10–37 residues), with a chain at the terminal (38–76 residues) [40,41,42]. The E protein plays a crucial biological role, not only in the structural integrity of the virus, but also in host virulence [43]. The E protein sequence of SARS-CoV-2 shows a 96.1% similarity to that of SARS-CoV [31].
The M protein plays a crucial role in maintaining the shape of the viral envelope (Table 1). This function can be achieved by interacting with other viral proteins that exhibit PPIs [44]. The M protein is also known as the central organization of coronavirus proteins. The binding of E to M produces the virus envelope, and this interaction is sufficient for the synthesis and release of viruses [45,46]. The binding of M with S is an important event for the retention of the S protein in the endoplasmic reticulum–Golgi complex as well as its integration into new viruses [46,47]. Moreover, the interaction of N with M stabilizes the nucleocapsid (RNA–N protein complex) and the internal core of viruses, resulting in the completion of viral assembly [47,48]. The M protein amino acid sequence of SARS-CoV-2 exhibits a 96.4% similarity with that of SARS-CoV [31].

{kind=link}
{kind=link}
Table 1.
The structural proteins (SPs) of coronaviruses and their physiological significance.
| Sr. No. | SPs | PDB ID | Residues | Physiological Significance | Reference |
|---|---|---|---|---|---|
| 1 | E | 7K3G | 76–109 | Virus assembly, morphogenesis, viral–host interaction, membrane permeability | [49] |
| 2 | M | 8CTK | 220–260 | Virus assembly, protein interactions (M–M, M–S, M–N) | [50] |
| 3 | N | 6VY0, 6YUN | 422 | Abundant RNA-binding protein, virion genome packaging | [51] |
| 4 | S | 6VYB | 1273 | Main antigen component, triggers the host immune response | [52] |
Table 2.
The non-structural proteins (NSPs) of coronaviruses and their physiological significance.
| Sr. No. | NSPs | PDB ID | Residues | Physiological Significance | Reference |
|---|---|---|---|---|---|
| 1 | NSP1 | 7K3N | 180 | Protein synthesis, prevents antiviral activity of host cells, degrades host mRNA | [53,54,55] |
| 2 | NSP2 | 7MSW | 638 | Genome replication, disruption of intracellular host signaling | [56,57,58] |
| 3 | NSP3 (Papain-like protease, PLpro) | 7KAG, 6WEY, 6WUU, 7LG0 | 1945 | Integral to viral replication, post-translational processing of the two polyproteins, suppresses host protein synthesis | [22,58,59] |
| 4 | NSP4 | 3GZF | 500 | Protects new replicated virions, replication and assembly of viral structures in host cell | [60,61] |
| 5 | NSP5 (3C-like protease, 3CLpro) | 6LU7 | 306 | Protein cleavage capacity (conserved feature) | [62,63] |
| 6 | NSP6 | - | 290 | Induction of autophagosomes, inhibition of viral components to reach host lysosomes | [64,65,66] |
| 7 | NSP7 | 7JLT | 83 | Primase complex (NSP7-NSP8), hetero-oligomeric complex (NSP7-NSP8-RdRp), viral replication | [67,68,69] |
| 8 | NSP8 | 7JLT | 198 | Primase complex (NSP7-NSP8), hetero-oligomeric complex (NSP7-NSP8-RdRp), viral replication | [67,68,69] |
| 9 | NSP9 | 6WXD | 113 | RNA synthesis, carries viral RNA to the host cell, responsible for proliferation | [70,71,72] |
| 10 | NSP10 | 6ZPE | 139 | Cofactor activation for replicative enzymes, complex NSP10-NSP14, viral RNA proofreading | [73,74,75] |
| 11 | NSP11 | - | 13 | Cleavage product of PP1a by 3CLpro/MPro | [21,76] |
| 12 | NSP12 (RNA polymerase, RdRp) | 6YYT | 932 | RNA polymerase activity | [29,77,78,79,80] |
| 13 | NSP13 | 6JYT | 601 | Helicase activity | [29,81] |
| 14 | NSP14 | 7R2V | 527 | Viral RNA methylation, viral RNA proofreading, methyltransferase activity | [73,82,83,84] |
| 15 | NSP15 | 6WXC | 346 | Endoribonuclease activity | [81,85] |
| 16 | NSP16 | 6WVN | 298 | Viral replication, immune response evasion Viral RNA methylation, methyltransferase activity | [84,86,87] |
3. Overview of TLR Signaling
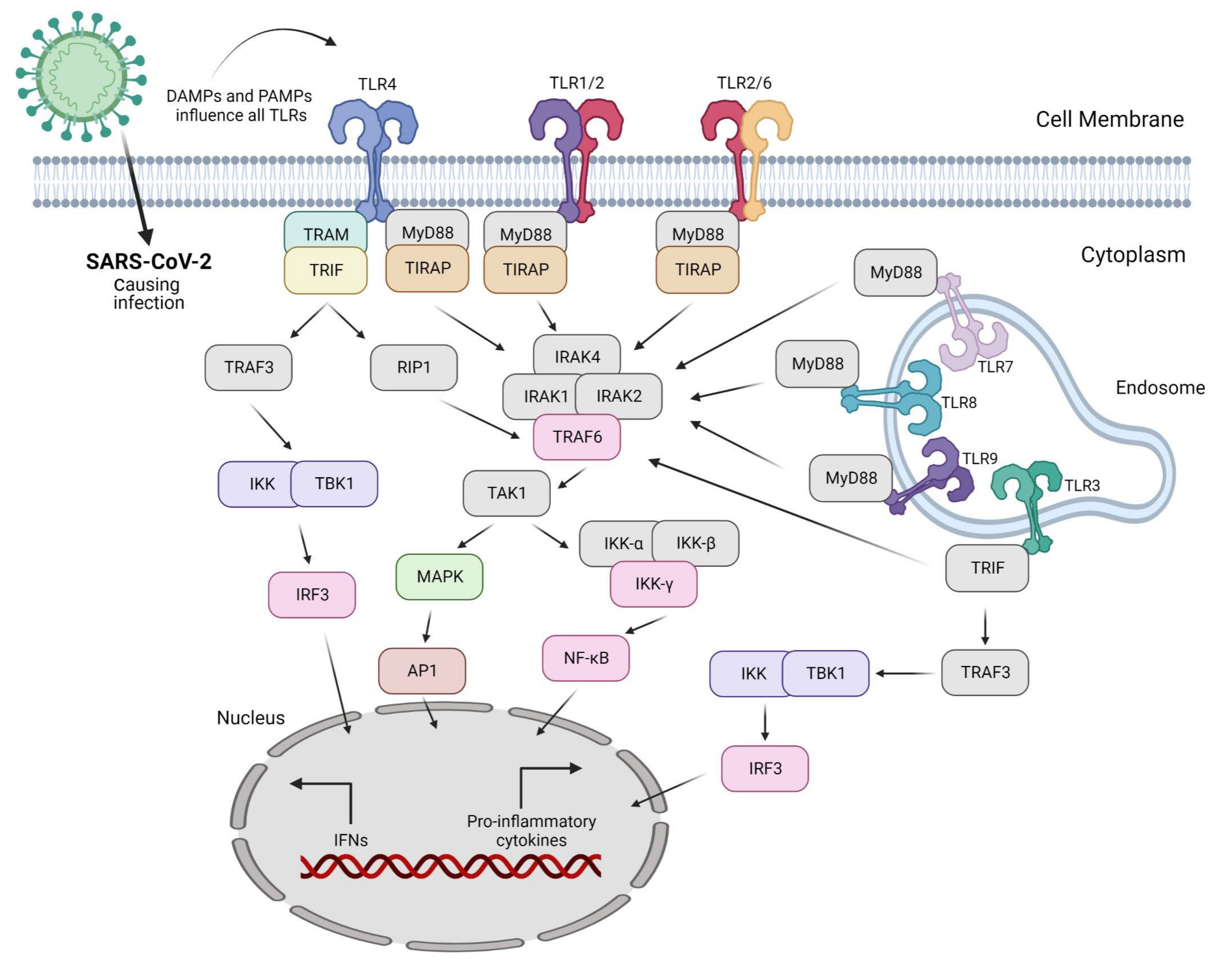
Invading pathogens stimulate the release of proinflammatory mediators in response to infection (Figure 1 and Figure 2). Signaling networks are necessary for the protection of the host against invading microorganisms. TLR signaling dysregulation plays a central role in the development and progression of infection. Inflammatory secretory molecules including chemokines, ILs, IFNs, and tumor necrosis factor-alpha (TNF-α) are part and parcel of TLR signaling, resulting in the modulation of cellular characteristics such as apoptosis, immune response, and proliferation [88,89,90]. Mitogen-activated protein kinases (MAPKs) and NF-κB are activated by TLRs. TLR3 and TLR4 are involved in the stimulation of IRF3. In contrast, IRF7 is triggered by TLR7–9 [91]. TLRs are stimulated by interactions with ligands to initiate an intracellular downstream signaling cascade, leading to activation of the host defense system [92].
The nature of the ligand and downstream adaptor molecules directs the TLR signaling cascade (Table 3). Two distinct pathways play critical roles in TLR signaling: MyD88-dependent and -independent pathways [93] (Figure 2). The former pathway employs all TLRs (except for TLR3), resulting in the biosynthesis of inflammatory cytokines [94]. In contrast, the latter pathway (also referred to as the TRIF-dependent pathway) involves TLR3 and TLR4, resulting in the expression of IFN-I [95]. In other words, the interaction of PAMP and PRR leads to the biosynthesis of proinflammatory cytokines as well as IFN-1, which is a cellular indication of the immune response [96]. Several negative regulators that enhance the activation of the innate immune response are involved in TLR-dependent signaling cascades. Hence, the overactivation of TLRs can lead to the interruption of immune cell homeostasis, resulting in the risk of inflammatory disorders [97]. Consequently, inhibitors (antagonists) targeting these receptors and/or cascades can serve as novel therapeutics to treat such disorders [98].
4. Role of Antiviral Drugs Employing TLRs
When a pathogen such as a virus invades, an antiviral immune response is evident in the host cells. Various conserved molecular patterns of PAMPs have been identified. As discussed above, TLRs are the key constituents of the innate immune system, and multiple TLRs (TLR1–4, TLR6–9) identify viral ligands [17,117,118,119]. With respect to their functional importance, TLRs might be potentially employed to treat not only inflammatory disorders but also viral diseases. This can be explained by a deep insight into the positive and negative mediators of TLRs [97,120]. TLR agonists lack accessory molecules but can mimic natural ligands; hence, they exhibit a low molecular weight and have potential for expanded pharmacokinetics and pharmacodynamics in comparison with the parent molecule. Moreover, TLR antagonists help to deal with autoimmune and inflammatory disorders by defeating unnecessary inflammation, resulting in an antibody- or cell-mediated response that suppresses disease progression [97,121,122].
Different approaches are employed by viruses in which they weaken their recognition by masking and/or increasing the dysregulation of mediators. Viruses disturb TLR signaling through their own mechanisms. Thus, TLRs are largely involved in the molecular interaction between viruses and host cells [5]. Various PRRs are engaged in the response to viral infection, which is also the case for TLRs. A thorough understanding of this interaction has facilitated the development of various strategies to limit viral infection including antiviral immunity as well as therapeutics [5]. Moreover, viral infection activates TLRs to increase cytokine levels, resulting in an antiviral innate immune response. The interaction between viruses and TLRs at every step of the signaling pathway plays an important role in developing effective antiviral therapies as well as in identifying novel molecular targets for the advancement in antiviral drugs [123]. The regulation of invasion, replication, and immune responses is a significant factor in viral pathogenesis [117]. Viral glycoproteins and NSPs released in the extracellular region are responsible for the stimulation of TLR2 and TLR4 due to their presence on the cellular surface [117,124,125]. In contrast, TLR3, TLR7/8, and TLR9, which are present in the endosomal compartment, contain viral double-stranded RNA (dsRNA) [126], ssRNA [114], and CpG DNA (unmethylated) [116], respectively.
TLR agonists have a positive effect on antiviral immunity and exhibit significant resistance against experimental infections [127,128,129]. The TLR–virus interaction involves a complex mechanism that is associated with the type of TLR as well as the type of virus. Moreover, multiple PRRs are required to initiate an immune response to various viral infections. Moreover, significant differences in TLR signaling have been reported between mice and humans. Therefore, therapeutic manipulation of TLRs requires an understanding of human cellular immunity [130]. Some examples are presented below.
TLR2 activation enhances the innate immune response to viral infections and can be used to treat viral respiratory diseases. Using the shock-and-kill strategy, immune cell recognition is enhanced and latently infected cells are eliminated [112,131]. TLRs can be used to reverse HIV-1 latency and trigger innate immune responses. In an evaluation of the effectiveness of SMU-Z1 (a novel TLR1/2 agonist), in addition to enhancing latent HIV-1 transcription (ex vivo), the NF-κB and MAPK pathways were also targeted in cells [131]. Latency-reversing agents have been employed for HIV reactivation, resulting in enhanced immune activation [112]. Dual TLR2/7 agonists were synthesized and characterized based on their latency-reversing ability, which were found to effectively reactivate the latency. TLR2 components reactivate HIV by NF-κB stimulation and the secretion of IL-22 (thereby enhancing the antiviral state and inhibiting HIV infection), whereas TLR7 components induce the secretion of TNF-α [112]. The activation of TLR2 in vivo has been assessed against rhinovirus infection [132]. Airway epithelial cells promote an extended immune response characterized by IFN-λ expression, NF-κB activation, and lymphocyte recruitment, resulting in a reduction in viral-induced inflammation and continued antiviral innate immunity [132].
TLR3 (the first identified antiviral TLR) in humans confers protective immunity against vaccinia virus (VACV) infection. In contrast, TLR3 is responsible for the detrimental effects of VACV infection in mice and TLR4 has the same effect in humans [133,134]. The recognition of dsRNA by TLR3 is further evidence of the role of TLRs in the antiviral response [119,126,135]. TLR3 signaling can be activated by a synthetic dsRNA agonist (a potent immune stimulant), resulting in protective immunity against multiple viruses including coronaviruses [136,137,138,139]. Viral-origin ssRNA sequences (rich in GU- and AU-) are detected by TLR7 and TLR8, which are functionally similar and only differ with respect to their expression patterns [113,130]. TLR7/8 expression is evident in dendritic cells, monocytes, and macrophages [140]. Additional examples are listed in Table 4.
5. Possible Molecular Interactions of SARS-CoV-2 with TLRs
SARS-CoV-2 is not only associated with viral illness but also with disorders of immunopathology. DAMPs and viral components act as TLR ligands for their overactivation. TLR4 (membrane-bound) and TLR3/7/8 (endosomal) play significant roles in the production of cytokine storms. The ssRNA of SARS-CoV-2 is recognized by TLR7/8, and after replication, the viral dsRNA is recognized by TLR3, which leads to TRIF-mediated inflammatory signaling [152]. The MyD88-dependent pathway (leading to overactivation of TLRs), related to the TRIF pathway, provides a possible link between SARS-CoV-2 and TLRs [153]. The production of type I (IFN-α and IFN-β) and type III [IFN-λ (1/2/3)] IFNs by TLRs is a significant antiviral feature that can be exploited for systematized viral control as well as clearance [117,119,154]. Type I and III IFNs perform the same function (despite their structural differences) in cellular signaling, although type III IFN receptors are primarily localized to the epithelial surface (airway epithelial cells) [155]. Cytokines (type III IFN) bind to their receptors, and the signal cascade is initiated by the JAK/STAT pathway, leading to the formation of IFN-stimulated genes [156]. Activation of the JAK/STAT pathway induced by TLRs may lead to macrophage activation syndrome [157]. Virally infected cells are killed by activated dendritic cells, natural killer cells, and macrophages stimulated by IFN [158]. SARS-CoV-2 infection results in higher levels of chemokines and proinflammatory cytokines in the blood [159,160]. These biological conditions lead to host cell death and organ injury [161]. Hence, the synthesis of DAMPs amplifies inflammation by TLR binding via the MyD88-dependent pathway. Elevated TLR stimulation, signaling cascades, and NF-κB may influence the severity of COVID-19 [153]. The nutritional profile has a basic influence on immunity. Compounds with immunomodulatory, anti-inflammatory as well as antiviral characteristics can be helpful against SARS-CoV-2 infection [162]. Various studies have suggested encouraging results in the case of nutraceuticals [163,164]. Compounds including astaxanthin, curcumin, glycyrrhizin, hesperidin, lactoferrin, luteolin, quercetin as well as resveratrol may inhibit and counteract the symptoms of COVID-19 [165,166,167,168,169,170,171,172].
Accordingly, IFN has been dynamically explored as a therapeutic target for COVID-19. This is because the release of type III IFN in the lungs could be responsible for the observed immunopathology of COVID-19 [173,174]. In contrast, type I IFN in combination with antiviral drugs has exhibited the opposite results including reduced systemic inflammation and viral clearance [173,174,175]. The synthesis of proinflammatory cytokines is associated with MyD88-dependent pathways, whereas the activation of type I and III IFNs is linked with the TRIF-dependent pathway [176,177] (Table 3). SARS-CoV-1 dsRNA and ssRNA are not detected by TLR3 and TLR7 and show some protective dodging mechanisms; hence, the same strategy could be used by SARS-CoV-2 [178,179]. The stimulation of TLRs by SARS-CoV-2 is responsible for activation of the inflammasome and the subsequent release of IL-1β and IL-6. Moreover, enhanced inflammasome activation is linked to non-promising consequences in patients with COVID-19 [180]. TLR2 signaling is activated by SARS-CoV-2 infection (Table 5). Thus, blocking of the signaling has been proposed as a potential target for the treatment of COVID-19 [181] because the strong effect of proinflammatory cytokines leads to disease severity through the activation of TLR2 [182]. In the context of infection with β-coronaviruses, MyD88, the TLR adaptor, has been reported to be a significant factor in the release of a large number of inflammatory cytokines [183]. SARS-CoV-2 interacts with various TLRs, directly or indirectly. Multiple interacting residues have been reported in the literature considering PPI and the design of agonists/antagonists. The interacting residues are based on experimental as well as computation studies. Only TLRs involved in virus sensing and/or signaling are displayed (Table 5).
Multiple TLR (2, 4, 7, 9)-deficient macrophages were infected with the mouse hepatitis virus. TLR2 deficiency resulted in the inhibition of TNF and IL6 expression as well as inflammatory cytokine genes. In contrast, other TLR deficiencies had negligible effects on these genes [182]. In the case of SARS-CoV-2, an inhibitor of TLR2 caused a noteworthy reduction in cytokine and chemokine release. This study demonstrated the role of TLR2 in sensing viral invasion upstream of MyD88 [182]. The TRIF pathway activated by TLR3 showed a protective response against Middle East respiratory syndrome (MERS)-CoV and SARS-CoV infections [184] (Table 3 and Table 5). Mice lacking TLR3, TLR4, and TRIF adaptor are exceedingly vulnerable to SARS-CoV and enhanced pulmonary infection, resulting in a risk of mortality [185]. Moreover, a role of TLR4 has been identified in the pathology of SARS-CoV-2, characterized by excessive inflammation in patients and activation of the inflammasome [186,187]. TLR4 inhibition in animal models has been shown to decrease lung injury by alleviating NF-κB pathway stimulation [188]. Viral infection and subsequent inflammation results in the production of DAMPs that act as ligands for TLR4. Heat shock proteins released from virus-infected cells act as TLR4 agonists [189]. TLR5 has been proposed as a target against SARS-CoV-2 in the development of drugs and vaccines [190].
During cytokine storms, elevated levels of IL-6 in the serum have been observed in patients with COVID-19 (Figure 1). TLR7 (activated by viral components) stimulates the MyD88-dependent pathway, resulting in the release of ILs and TNF-α, particularly IL-6 [191,192]. Structurally and phylogenetically similar receptors but different TLR7/8 agonists synthesize different cytokines [193]. ssRNA fragments in SARS-CoV-2 induced by TLR7/8 have been detected [194]. Whole-genome sequencing of SARS-CoV-2 in comparison with other coronaviruses (MERS-CoV and SARS-CoV) has revealed that TLR7 could be significantly involved in COVID-19 as the viral genome contains more ssRNA motifs that can bind to TLR7 [195]. Moreover, the TLR7 agonists imiquimod and imidazoquinolinone (with a role in TLR7activation) are under investigation as potential therapeutics against COVID-19. These drugs have been observed to decrease systemic inflammation and innate immune activation due to their antiviral effects [196,197]. RNA and DNA rich in unmethylated CpG islands can be recognized by TLR9. Both viral and mitochondrial DNA enriched in the same sequence are associated with inflammatory responses involving TLR9-mediated signaling. Moreover, the activation of p53 [198] and mammalian target of rapamycin (mTOR) is being considered as a therapeutic target against SARS-CoV-2. mTOR blockers are also associated with the MyD88 and TLR9 pathways [199].
Table 5.
The interaction of Toll-like receptors (TLRs) with SARS-CoV-2 and other coronaviruses.
| Coronaviruses | TLRs | Interacting Residues of TLRs | References |
|---|---|---|---|
| SARS-CoV-2 | TLR2 | Tyr323, Phe325, Val 348, Phe349 | [182,200,201] |
| TLR3 | His39, His60, His108, Asn515, Asn517, His539, Asn541, Arg544, Ser571 | [111,202,203] | |
| TLR4 | Arg264, Glu266, Asp294, Tyr295, Tyr296, Thr319, Glu321, Lys341, Lys362, Gly363, Gly364, Arg382 | [188,204,205] | |
| TLR7/8 | Phe349, Tyr356, Gly379, Val381, Phe408, Asp555, Leu557, Gly584, Thr586 | [114,206,207,208] | |
| SARS-CoV | TLR3 | His39, His60, His108, Asn515, Asn517, His539, Asn541, Arg544, Ser571 | [136,185,203] |
| TLR4 | Arg264, Glu266, Asp294, Tyr295, Tyr296, Thr319, Glu321, Lys341, Lys362, Gly363, Gly364, Arg382 | [185,205] | |
| TLR7/8 | Phe349, Tyr356, Gly379, Val381, Phe408, Asp555, Leu557, Gly584, Thr586 | [207,208,209] | |
| MERS-CoV | TLR3 | His39, His60, His108, Asn515, Asn517, His539, Asn541, Arg544, Ser571 | [203,209] |
| TLR4 | Arg264, Glu266, Asp294, Tyr295, Tyr296, Thr319, Glu321, Lys341, Lys362, Gly363, Gly364, Arg382 | [205,210,211] | |
| TLR7/8 | Phe349, Tyr356, Gly379, Val381, Phe408, Asp555, Leu557, Gly584, Thr586 | [207,208,209,212,213] |
6. Promising Drug Targets in SARS-CoV-2
Possible and effective drug targets as well as therapeutic agents against SARS-CoV-2 have been suggested by various researchers worldwide [214]. For example, virulence factors, enzymes, host-specific receptors, and glycosylated-structural proteins have been identified in pathological conditions caused by the coronavirus [215]. Activators of transcription signaling pathways, proinflammatory cytokines, Janus kinase/signal transducers, and NSPs also play a crucial role in the pathology. Antiviral therapeutic strategies such as drug repurposing depend on chemical and molecular interactions between the host machinery and viral small molecules [215].
Low-molecular-weight molecules from plants (phytochemicals) have been tested for their antiviral activity. Compounds extracted from plants have been shown to exhibit antiviral activity against SARS-CoV in Vero cells. Lycorine was identified as the active ingredient of Lindera aggregata, and it has been suggested that the plant extract and lycorine can be a good option for the development of novel antiviral drugs [7]. In plants, secondary metabolites are produced by metabolic pathways, which are also referred to as phytochemicals [216]. These metabolites have been screened for their efficacy against microbes and viruses. Various phytochemicals have been shown to inhibit viral infection and replication [217]. Bioactive phytochemicals can improve and strengthen host immunity. For example, less vulnerability to infections and assistance in the stoppage of viral infections through host immune function have been observed with the treatment of vitamins A and C [218]. Various in vitro, in vivo, and in silico models using marine-derived natural compounds exhibiting promising anti-SARS-CoV-2 efficacy have been previously summarized [219].
Various proteins including ACE-2, RdRp, 3CLpro, PLpro, RBD, and cathepsin L could be operative therapeutic targets [67,198,220,221,222,223]. Although several molecules have been suggested as drug candidates, currently, there are no accessible operative anti-CoV mediators. Molecular interactions between ACE2 and SARS viruses are determinants of the initial infection. Hence, renin–angiotensin–aldosterone system (RAAS) inhibitors may modify ACE2 expression, resulting in reduced SARS-CoV-2 virulence. ACE2 (type I transmembrane-metallocarboxypeptidase enzyme) controls the effects of RAAS and is a key receptor for both SARS-CoV-1 and SARS-CoV-2, which facilitates entry into human lung cells through the S protein of the coronavirus [224,225,226,227,228]. Considering the complexity of the pathogenesis of SARS-CoV-2, clinically approved drugs that stimulate ACE2 may serve as operative anti-SARS-CoV-2 therapeutics [229]. ACE inhibitors (captopril) stimulate the ACE2/angiotensin (1–7)/receptor axis [230]. In animal models, treatment with angiotensin receptor blockers was shown to enhance ACE2 expression [231,232]. The ACE2–RBD complex is proteolytically regulated by type-2 transmembrane cellular serine protease (TMPRSS2), which leads to ACE2 cleavage and S protein activation [233]. The RBD (S protein) of SARS-CoV-2 contains more ACE2-interacting residues (Tyr473, Gln474, Cys488, Tyr489, Val524, and Cys525) than that of SARS-CoV, and is involved in loop formation. These mutations are evident in the sequence (RBD) of SARS-CoV-2 [234,235,236]. Moreover, two binding hotspot residues (Lys31 and Lys353) have been reported to be more sensitive to S protein binding. Lys31 and Lys353 formed salt-bridge(s) with Glu35 and Asp38, respectively, surrounded by a hydrophobic region [237]. Additionally, other studies support the development of promising ACE2 inhibitors for SARS-CoV-2 [220,238,239].
In the case of glycosylated S protein, membrane fusion inhibitors for the S2 subunit and antibodies (monoclonal) targeting the S1 subunit could be operative therapeutic mediators to treat coronavirus infection. Vaccine development has also been promoted against coronaviruses. Small-molecule inhibitors (SMIs) might be suitable for inhaled and/or oral administration, exhibit less mutation and strain sensitivity, are less immunogenic, and convenient. Novel drug-like SMIs (DRI-C23041 and DRI-C91005) have been identified. These SMIs inhibit the interaction between S protein and human ACE2 [34]. Moreover, griffithsin, a compound derived from red algae, adheres to the SARS-CoV-2 glycosylated S protein as well as to HIV [240]. Furin, a serine endoprotease, cleaves S1–S2 and may be suitable as an anti-SARS-CoV-2 agent [241]. Emodin, a Rheum tangutica-derived compound, not only inhibits the interaction between the ACE2 receptor and SARS-CoV-2 but also blocks SARS-CoV ORF3a [242,243]. Moreover, the host protease is employed by SARS-CoV-2 for the priming of the S protein. Camostat mesylate, an inhibitor of proteases, helps in the infection of lung cell lines [244]. Similar to the S protein, other structural proteins as well as NSPs have been highlighted as potential targets for the development of antiviral drugs. In the RBD, 14 different potent residues of the S protein have been identified that interact significantly with ACE2, resulting in the stability of the complex [245], while 15 significant residues in the RBD of the S protein have been reported in the case of the Omicron SARS-CoV-2 variant [246]. Both of these studies analyzed (in silico) natural compounds for their anti-SARS-CoV-2 bioactivity [245,246]. Additionally, terpenes (natural compounds) have been suggested as anti-SARS-CoV2 binding agents between the RBD and ACE2 receptor. Terpenes showed a strong affinity for RBD and inhibited its interaction with ACE2 [245].
Toremifene, a nonsteroidal selective estrogen receptor modulator, was found to block the viral replication of coronaviruses such as MERS-CoV and SARS-CoV [247] and the Ebola virus [248] by targeting viral membrane proteins. Hence, it has also become a potential candidate inhibitor of SARS-CoV-2 replication [248]. Moreover, a team of researchers proposed a region (residues) in the M protein for the development of novel drugs and/or peptides to block dimer formation [249]. The interaction of the M protein (heterodimer) with the S and E proteins (via PPIs) has been proposed by computational analysis, and key amino acids for the M–E complex (W55, F96, F103) and M–S complex (Y71, Y75) have been identified [250].
A high percentage of E protein is expressed inside the infected cells, which is responsible for viral assembly, maturation, budding, and proliferation [35,40,214]. The percentage similarity of the SARS-CoV-2 E protein sequence with that of other coronaviruses (96.1%) [31] demonstrates the potential for repurposing and/or development of pan-anti-corona drug candidates. Small molecules (phytochemicals) such as belachinal, vibsanol B, and macaflavanone E have been evaluated for the inhibition of E protein activity by in silico analyses [251].
The N protein exhibits essential activities such as proliferation of the virus as another important component, similar to other SPs. This provides a promising area for developing effective therapeutics to inhibit viral proliferation. For example, glycogen synthase kinase-3 (GSK-3), also known as serine/threonine protein kinase, is an important component of N protein phosphorylation. GSK-3 inhibitors inhibit N protein phosphorylation and result in damaged proliferation (infected lung epithelial cells) in SARS-CoV-2 in a cell type-dependent manner [252]. Candidate inhibitors of the N protein have been suggested by a screening method on a biochip platform using a quantum-dot (QD) RNA oligonucleotide. The novel anti-SARS potential of catechin gallate and gallocatechin has been identified. These two molecules (0.05 µg/mL) presented more than 40% inhibition activity on a QD-based RNA oligonucleotide system [253]. Computational analysis has suggested that zidovudine triphosphate is a potent inhibitor of the N protein of SARS-CoV-2 [254]. Based on an in silico approach, another repurposing study shed light on vanganciclovir, which is approved for treating patients with HIV and shows activity against N protein as well as the main protease [254].
SARS-CoV-2 depends on proteases of the Golgi apparatus to synthesize NSP1–16 in the host cell [27]. NSP3 [papain-like protease (PLpro)] is a multidomain protein and the largest protein in the coronavirus genome. Several regions of the NSP3 gene are involved in viral replication. NSP3 contains a SARS-unique domain that can attach to G-quadruplexes, which are guanine-rich non-canonical nucleic acid structures that are essential for SARS-CoV replication. SARS-CoV-2 NSP3 shows structural similarities [255]. By developing a protease assay and screening a custom compound library, two molecules (dihydrotanshinone I and Ro 08-2750) were identified to significantly inhibit PLpro in protease. Additionally, the inhibition of viral replication was evaluated by an isopeptidase assay using cell culture [256]. Another protease, NSP5 (3CLpro), was identified as a primary target (similar to PLpro) for coronavirus drug discovery. Both of these targets are crucial and have conserved activity in the proteolytic processing of viral replicase polyproteins [257].
Coronaviruses encode two or three proteases that cleave replicase polyproteins. Many NSPs assemble into the replicase–transcriptase complex, which generates a reasonable environment for RNA synthesis and subsequent replication as well as the transcription of sub-genomic RNAs [258]. Replicase polyproteins 1a and 1ab are comprised of NSPs11 and 16, respectively [259]. MPro (the main protease), commonly known as NSP5, is employed for the cleavage of these polyproteins, exhibiting crucial events of viral assembly and maturation [259]. MPro is a dimer (306 residues) with two identical monomers, and is a significant target responsible for viral polyprotein cleavage 1ab at 11 (a major cleavage site), required for generating the NSP7–NSP8–NSP12 complex (viral replication complex) [260,261]. Residues interacting with two novel inhibitors against MPro have been identified: His41, Met49, Met165, Val186, Asp187, Arg188 as well as Gln189, exhibiting hydrophobic and H-bonding. Both inhibitors reside in the substrate-binding site and inhibit the enzymatic activity of MPro in SARS-CoV-2 [261].
Targeting highly conserved genes and/or proteins including RdRp (NSP12), MPro, and helicases is a promising antiviral drug development approach to inhibit the replication and proliferation of SARS-CoV-2 [262]. Hence, inhibitors targeting these enzymes may reduce the threat of mutation-mediated drug resistance and facilitate effective antiviral protection [262]. A conserved motif (Ser-Aps-Asp) in the RdRp domain was identified at the C-terminus. The binding and activity of RdRp were enhanced by the NSP7–NSP8 (cofactor) complex. This binding stabilizes the entire closed conformation, which is packed beside the thumb–finger interface. The binding residues between the RdRp and RNA complex and RdRp docking to develop inhibitors have been extensively studied [78,79,80].
The inhibition of RdRP is important as one of the key strategies for developing antiviral therapeutics. The selective inhibition of RdRp may not cause noteworthy side effects or toxicity in host cells [263]. Natural compounds and their derivatives have exhibited significant binding affinity to RdRp [264,265,266], with promising outcomes that require further investigation.
NSPs 7–16 are responsible for coronavirus RNA synthesis and processing, which generate two large replicase polyproteins by cleavage. SARS-CoV-2 possesses a large number of enzymes that are responsible for RNA synthesis as well as RNA processing. The genome that is expressed and replicated by enzymes is two to three times larger than that of any other RNA viruses. RdRp is an important drug target because of its vital role in generating viral RNA [77,78,79,80].
Coronaviruses possess three important virulence factors: NSP1, NSP3c, and ORF7a. These factors help in the escape of viruses from host innate immunity and may be potential drug targets [55,267]. NSP1 and NSP3c interact with the host 40S ribosomal subunit and adenosine diphosphate-ribose, respectively. This leads to the degradation of mRNA and the inhibition of type-I IFN synthesis by NSP1, while NSP3c assists viruses to counterattack the immune response of the host [55,268]. ORF7a is directly attached to bone marrow matrix antigen-2 (BST-2) and has the ability to stop its glycosylation. BST-2 plays a specific role in regulating the release of newly synthesized viruses [267,269].
7. Conclusions
SARS-CoV-2 is recognized by various TLRs. Surface (TLR2 and 4) and intracellular (TRL3, 7/8, and 9) factors have been reported to be involved in the perception of SARS-CoV-2 infection by the immune system. Multiple adaptors such as MyD88 and TRIF are recruited by TLRs to initiate downstream signaling pathways. Various protein targets from viruses and the host machinery have been suggested as potential drug targets against SARS-CoV-2. Protein targets from viruses include both structural and nonstructural proteins. Similarly, TLRs are functional protein targets during SARS-CoV-2 infection.
Author Contributions
Conceptualization, A.M. and R.H.P.; Writing—original draft preparation, A.M.; Writing—review and editing, A.M., R.H.P. and M.H.; Visualization, A.M. and M.H.; Supervision, S.C.; Funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Korea Drug Development Fund funded by the Ministry of Science and ICT, Ministry of Trade, Industry, and Energy, and Ministry of Health and Welfare (HN21C1058). This study was also supported by the National Research Foundation of Korea (NRF-2022M3A9G1014520, 2019M3D1A1078940, and 2019R1A6A1A11051471).
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Iwasaki, A.; Medzhitov, R. Control of Adaptive Immunity by the Innate Immune System. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.C.; Choi, S. Recent Progress in the Development of Toll-like Receptor (TLR) Antagonists. Expert Opin. Ther. Pat. 2016, 26, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.; Cai, S.-Y.; Shao, J.-Z.; Chen, J. Toll-Like Receptors, Associated Biological Roles, and Signaling Networks in Non-Mammals. Front. Immunol. 2018, 9, 1523. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, H.; Zhao, C.; Zhang, H. Evolutionary History of the Toll-Like Receptor Gene Family across Vertebrates. Genome Biol. Evol. 2020, 12, 3615–3634. [Google Scholar] [CrossRef] [PubMed]
- Carty, M.; Bowie, A.G. Recent Insights into the Role of Toll-like Receptors in Viral Infection. Clin. Exp. Immunol. 2010, 161, 397–406. [Google Scholar] [CrossRef]
- Abe, T.; Kaname, Y.; Hamamoto, I.; Tsuda, Y.; Wen, X.; Taguwa, S.; Moriishi, K.; Takeuchi, O.; Kawai, T.; Kanto, T.; et al. Hepatitis C Virus Nonstructural Protein 5A Modulates the Toll-like Receptor-MyD88-Dependent Signaling Pathway in Macrophage Cell Lines. J. Virol. 2007, 81, 8953–8966. [Google Scholar] [CrossRef]
- Li, S.Y.; Chen, C.; Zhang, H.Q.; Guo, H.Y.; Wang, H.; Wang, L.; Zhang, X.; Hua, S.N.; Yu, J.; Xiao, P.G.; et al. Identification of Natural Compounds with Antiviral Activities against SARS-Associated Coronavirus. Antivir. Res. 2005, 67, 18. [Google Scholar] [CrossRef]
- Maloney, G.; Schröder, M.; Bowie, A.G. Vaccinia Virus Protein A52R Activates P38 Mitogen-Activated Protein Kinase and Potentiates Lipopolysaccharide-Induced Interleukin-10. J. Biol. Chem. 2005, 280, 30838–30844. [Google Scholar] [CrossRef]
- Stack, J.; Haga, I.R.; Schröder, M.; Bartlett, N.W.; Maloney, G.; Reading, P.C.; Fitzgerald, K.A.; Smith, G.L.; Bowie, A.G. Vaccinia Virus Protein A46R Targets Multiple Toll-like-Interleukin-1 Receptor Adaptors and Contributes to Virulence. J. Exp. Med. 2005, 201, 1007–1018. [Google Scholar] [CrossRef]
- Blasius, A.L.; Beutler, B. Intracellular Toll-like Receptors. Immunity 2010, 32, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Onofrio, L.; Caraglia, M.; Facchini, G.; Margherita, V.; de Placido, S.; Buonerba, C. Toll-like Receptors and COVID-19: A Two-Faced Story with an Exciting Ending. Future Sci. OA 2020, 6, FSO605. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.; Davidson, S.M.; Yellon, D.M. The Cytokine Storm of COVID-19: A Spotlight on Prevention and Protection. Expert Opin. Ther. Targets 2020, 24, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, D.; Bhandari, R.; Kuhad, A. TLR4 as a Therapeutic Target for Respiratory and Neurological Complications of SARS-CoV-2. Expert Opin. Ther. Targets 2021, 25, 491–508. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like Receptor Signaling Pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Bowie, A.G. The Family of Five: TIR-Domain-Containing Adaptors in Toll-like Receptor Signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Innate Immunity to Virus Infection. Immunol. Rev. 2009, 227, 75–86. [Google Scholar] [CrossRef]
- Jiménez-Dalmaroni, M.J.; Gerswhin, M.E.; Adamopoulos, I.E. The Critical Role of Toll-like Receptors-From Microbial Recognition to Autoimmunity: A Comprehensive Review. Autoimmun. Rev. 2016, 15, 1–8. [Google Scholar] [CrossRef]
- Singh, N.; Baby, D.; Rajguru, J.; Patil, P.; Thakkannavar, S.; Pujari, V. Inflammation and Cancer. Ann. Afr. Med. 2019, 18, 121–126. [Google Scholar] [CrossRef]
- Wu, Y.W.; Tang, W.; Zuo, J.P. Toll-like Receptors: Potential Targets for Lupus Treatment. Acta Pharmacol. Sin. 2015, 36, 1395–1407. [Google Scholar] [CrossRef]
- Chan, J.F.W.; Kok, K.H.; Zhu, Z.; Chu, H.; To, K.K.W.; Yuan, S.; Yuen, K.Y. Genomic Characterization of the 2019 Novel Human-Pathogenic Coronavirus Isolated from a Patient with Atypical Pneumonia after Visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Snijder, E.J.; Decroly, E.; Ziebuhr, J. The Nonstructural Proteins Directing Coronavirus RNA Synthesis and Processing. Adv. Virus Res. 2016, 96, 59. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.T.; Lai, M.M.C. Viral and Cellular Proteins Involved in Coronavirus Replication. In Coronavirus Replication and Reverse Genetics; Springer: Berlin/Heidelberg, Germany, 2005; Volume 287, p. 95. [Google Scholar] [CrossRef]
- Guo, Y.R.; Cao, Q.D.; Hong, Z.S.; Tan, Y.Y.; Chen, S.D.; Jin, H.J.; Tan, K.S.; Wang, D.Y.; Yan, Y. The Origin, Transmission and Clinical Therapies on Coronavirus Disease 2019 (COVID-19) Outbreak-an Update on the Status. Mil. Med. Res. 2020, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Du, J.; Su, H.; Zhang, J.; Zhu, G.; Zhang, S.; Wu, Z.; Jin, Q. Identification of Diverse Bat Alphacoronaviruses and Betacoronaviruses in China Provides New Insights into the Evolution and Origin of Coronavirus-Related Diseases. Front. Microbiol. 2019, 10, 1900. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E. Epidemiology, Virology, and Clinical Features of Severe Acute Respiratory Syndrome -Coronavirus-2 (SARS-CoV-2; Coronavirus Disease-19). Clin. Exp. Pediatr. 2020, 63, 119–124. [Google Scholar] [CrossRef]
- Gasmalbari, E.; Abbadi, O.S. Non-Structural Proteins of SARS-CoV-2 as Potential Sources for Vaccine Synthesis. Infect. Dis. Trop. Med. 2020, 6, e667. [Google Scholar]
- Gorla, U.S.; Rao, G.S.N.K. SARS-CoV-2: The Prominent Role of Non-Structural Proteins (NSPS) in COVID-19. Indian J. Pharm. Educ. Res. 2020, 54, S381–S389. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. The Proteins of Severe Acute Respiratory Syndrome Coronavirus-2 (SARS CoV-2 or n-COV19), the Cause of COVID-19. Protein J. 2020, 39, 198–216. [Google Scholar] [CrossRef]
- Mousavizadeh, L.; Ghasemi, S. Genotype and Phenotype of COVID-19: Their Roles in Pathogenesis. J. Microbiol. Immunol. Infect. 2021, 54, 159–163. [Google Scholar] [CrossRef]
- Mohammed, M.E.A. The Percentages of SARS-CoV-2 Protein Similarity and Identity with SARS-CoV and BatCoV RaTG13 Proteins Can Be Used as Indicators of Virus Origin. J. Proteins Proteom. 2021, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.J.; Jiang, S. The Spike Protein of SARS-CoV--a Target for Vaccine and Therapeutic Development. Nat. Rev. Microbiol. 2009, 7, 226–236. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Cao, D.; Zhang, Y.; Ma, J.; Qi, J.; Wang, Q.; Lu, G.; Wu, Y.; Yan, J.; Shi, Y.; et al. Cryo-EM Structures of MERS-CoV and SARS-CoV Spike Glycoproteins Reveal the Dynamic Receptor Binding Domains. Nat. Commun. 2017, 8, 15092. [Google Scholar] [CrossRef] [PubMed]
- Bojadzic, D.; Alcazar, O.; Chen, J.; Chuang, S.T.; Condor Capcha, J.M.; Shehadeh, L.A.; Buchwald, P. Small-Molecule Inhibitors of the Coronavirus Spike: ACE2 Protein-Protein Interaction as Blockers of Viral Attachment and Entry for SARS-CoV-2. ACS Infect. Dis. 2021, 7, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- Satarker, S.; Nampoothiri, M. Structural Proteins in Severe Acute Respiratory Syndrome Coronavirus-2. Arch. Med. Res. 2020, 51, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Hao, Q.; Mu, Y.; Timani, K.A.; Ye, L.; Zhu, Y.; Wu, J. Nucleocapsid Protein of SARS-CoV Activates the Expression of Cyclooxygenase-2 by Binding Directly to Regulatory Elements for Nuclear Factor-Kappa B and CCAAT/Enhancer Binding Protein. Int. J. Biochem. Cell Biol. 2006, 38, 1417–1428. [Google Scholar] [CrossRef]
- Wang, Q.; Li, C.; Zhang, Q.; Wang, T.; Li, J.; Guan, W.; Yu, J.; Liang, M.; Li, D. Interactions of SARS Coronavirus Nucleocapsid Protein with the Host Cell Proteasome Subunit P42. Virol. J. 2010, 7, 99. [Google Scholar] [CrossRef]
- Lu, X.; Pan, J.; Tao, J.; Guo, D. SARS-CoV Nucleocapsid Protein Antagonizes IFN-β Response by Targeting Initial Step of IFN-β Induction Pathway, and Its C-Terminal Region Is Critical for the Antagonism. Virus Genes 2011, 42, 37. [Google Scholar] [CrossRef]
- Luo, H.; Chen, Q.; Chen, J.; Chen, K.; Shen, X.; Jiang, H. The Nucleocapsid Protein of SARS Coronavirus Has a High Binding Affinity to the Human Cellular Heterogeneous Nuclear Ribonucleoprotein A1. FEBS Lett. 2005, 579, 2623. [Google Scholar] [CrossRef]
- Ruch, T.R.; Machamer, C.E. The Hydrophobic Domain of Infectious Bronchitis Virus E Protein Alters the Host Secretory Pathway and Is Important for Release of Infectious Virus. J. Virol. 2011, 85, 675–685. [Google Scholar] [CrossRef]
- Kuo, L.; Hurst, K.R.; Masters, P.S. Exceptional Flexibility in the Sequence Requirements for Coronavirus Small Envelope Protein Function. J. Virol. 2007, 81, 2249–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdiá-Báguena, C.; Nieto-Torres, J.L.; Alcaraz, A.; Dediego, M.L.; Enjuanes, L.; Aguilella, V.M. Analysis of SARS-CoV E Protein Ion Channel Activity by Tuning the Protein and Lipid Charge. Biochim. Biophys. Acta 2013, 1828, 2026–2031. [Google Scholar] [CrossRef] [PubMed]
- Venkatagopalan, P.; Daskalova, S.M.; Lopez, L.A.; Dolezal, K.A.; Hogue, B.G. Coronavirus Envelope (E) Protein Remains at the Site of Assembly. Virology 2015, 478, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Schoeman, D.; Fielding, B.C. Coronavirus Envelope Protein: Current Knowledge. Virol. J. 2019, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Corse, E.; Machamer, C.E. Infectious Bronchitis Virus E Protein Is Targeted to the Golgi Complex and Directs Release of Virus-like Particles. J. Virol. 2000, 74, 4319–4326. [Google Scholar] [CrossRef] [PubMed]
- Mortola, E.; Roy, P. Efficient Assembly and Release of SARS Coronavirus-like Particles by a Heterologous Expression System. FEBS Lett 2004, 576, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Fehr, A.R.; Perlman, S. Coronaviruses: An Overview of Their Replication and Pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar] [CrossRef]
- Narayanan, K.; Maeda, A.; Maeda, J.; Makino, S. Characterization of the Coronavirus M Protein and Nucleocapsid Interaction in Infected Cells. J. Virol. 2000, 74, 8127–8134. [Google Scholar] [CrossRef]
- Liu, D.X.; Yuan, Q.; Liao, Y. Coronavirus Envelope Protein: A Small Membrane Protein with Multiple Functions. Cell Mol. Life Sci 2007, 64, 2043–2048. [Google Scholar] [CrossRef]
- Arndt, A.L.; Larson, B.J.; Hogue, B.G. A Conserved Domain in the Coronavirus Membrane Protein Tail Is Important for Virus Assembly. J. Virol. 2010, 84, 11418–11428. [Google Scholar] [CrossRef]
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 Nucleocapsid Protein Is Dynamic, Disordered, and Phase Separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.F.; Xu, W.; Liu, S. wen Structural and Functional Properties of SARS-CoV-2 Spike Protein: Potential Antivirus Drug Development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Züst, R.; Cervantes-Barragán, L.; Kuri, T.; Blakqori, G.; Weber, F.; Ludewig, B.; Thiel, V. Coronavirus Non-Structural Protein 1 Is a Major Pathogenicity Factor: Implications for the Rational Design of Coronavirus Vaccines. PLoS Pathog. 2007, 3, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Afsar, M.; Narayan, R.; Akhtar, M.N.; Das, D.; Rahil, H.; Nagaraj, S.K.; Eswarappa, S.M.; Tripathi, S.; Hussain, T. Drug Targeting Nsp1-Ribosomal Complex Shows Antiviral Activity against SARS-CoV-2. Elife 2022, 11, e74877. [Google Scholar] [CrossRef]
- Kamitani, W.; Narayanan, K.; Huang, C.; Lokugamage, K.; Ikegami, T.; Ito, N.; Kubo, H.; Makino, S. Severe Acute Respiratory Syndrome Coronavirus Nsp1 Protein Suppresses Host Gene Expression by Promoting Host MRNA Degradation. Proc. Natl. Acad. Sci. USA 2006, 103, 12885–12890. [Google Scholar] [CrossRef]
- Ma, J.; Chen, Y.; Wu, W.; Chen, Z. Structure and Function of N-Terminal Zinc Finger Domain of SARS-CoV-2 NSP2. Virol. Sin. 2021, 36, 1104–1112. [Google Scholar] [CrossRef]
- Cornillez-Ty, C.T.; Liao, L.; Yates, J.R.; Kuhn, P.; Buchmeier, M.J. Severe Acute Respiratory Syndrome Coronavirus Nonstructural Protein 2 Interacts with a Host Protein Complex Involved in Mitochondrial Biogenesis and Intracellular Signaling. J. Virol. 2009, 83, 10314–10318. [Google Scholar] [CrossRef]
- Angeletti, S.; Benvenuto, D.; Bianchi, M.; Giovanetti, M.; Pascarella, S.; Ciccozzi, M. COVID-2019: The Role of the Nsp2 and Nsp3 in Its Pathogenesis. J Med Virol 2020, 92, 584–588. [Google Scholar] [CrossRef]
- Khan, M.T.; Zeb, M.T.; Ahsan, H.; Ahmed, A.; Ali, A.; Akhtar, K.; Malik, S.I.; Cui, Z.; Ali, S.; Khan, A.S.; et al. SARS-CoV-2 Nucleocapsid and Nsp3 Binding: An in Silico Study. Arch. Microbiol. 2021, 203, 59–66. [Google Scholar] [CrossRef]
- Alsaadi, E.A.J.; Jones, I.M. Membrane Binding Proteins of Coronaviruses. Future Virol. 2019, 14, 275–286. [Google Scholar] [CrossRef]
- Oostra, M.; te Lintelo, E.G.; Deijs, M.; Verheije, M.H.; Rottier, P.J.M.; de Haan, C.A.M. Localization and Membrane Topology of Coronavirus Nonstructural Protein 4: Involvement of the Early Secretory Pathway in Replication. J. Virol. 2007, 81, 12323–12336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmy, Y.A.; Fawzy, M.; Elaswad, A.; Sobieh, A.; Kenney, S.P.; Shehata, A.A. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J. Clin. Med. 2020, 9, 1225. [Google Scholar] [CrossRef] [PubMed]
- Stobart, C.C.; Sexton, N.R.; Munjal, H.; Lu, X.; Molland, K.L.; Tomar, S.; Mesecar, A.D.; Denison, M.R. Chimeric Exchange of Coronavirus Nsp5 Proteases (3CLpro) Identifies Common and Divergent Regulatory Determinants of Protease Activity. J. Virol. 2013, 87, 12611. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, D.; Angeletti, S.; Giovanetti, M.; Bianchi, M.; Pascarella, S.; Cauda, R.; Ciccozzi, M.; Cassone, A. Evolutionary Analysis of SARS-CoV-2: How Mutation of Non-Structural Protein 6 (NSP6) Could Affect Viral Autophagy. J. Infect. 2020, 81, e24–e27. [Google Scholar] [CrossRef] [PubMed]
- Cottam, E.M.; Whelband, M.C.; Wileman, T. Coronavirus NSP6 Restricts Autophagosome Expansion. Autophagy 2014, 10, 1426–1441. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Y.; Huang, Z.; Xu, W.; Hu, W.; Yi, L.; Liu, Z.; Chan, H.; Zeng, J.; Liu, X.; et al. SARS-CoV-2 Non-Structural Protein 6 Triggers NLRP3-Dependent Pyroptosis by Targeting ATP6AP1. Cell Death Differ. 2022, 29, 1240–1254. [Google Scholar] [CrossRef]
- Biswal, M.; Diggs, S.; Xu, D.; Khudaverdyan, N.; Lu, J.; Fang, J.; Blaha, G.; Hai, R.; Song, J. Two Conserved Oligomer Interfaces of NSP7 and NSP8 Underpin the Dynamic Assembly of SARS-CoV-2 RdRP. Nucleic Acids Res. 2021, 49, 5956–5966. [Google Scholar] [CrossRef]
- Krichel, B.; Falke, S.; Hilgenfeld, R.; Redecke, L.; Uetrecht, C. Processing of the SARS-CoV Pp1a/Ab Nsp7-10 Region. Biochem. J. 2020, 477, 1009–1019. [Google Scholar] [CrossRef]
- Te Velthuis, A.J.W.; van den Worm, S.H.E.; Snijder, E.J. The SARS-Coronavirus Nsp7+nsp8 Complex Is a Unique Multimeric RNA Polymerase Capable of Both de Novo Initiation and Primer Extension. Nucleic Acids Res. 2012, 40, 1737–1747. [Google Scholar] [CrossRef]
- Zeng, Z.; Deng, F.; Shi, K.; Ye, G.; Wang, G.; Fang, L.; Xiao, S.; Fu, Z.; Peng, G. Dimerization of Coronavirus Nsp9 with Diverse Modes Enhances Its Nucleic Acid Binding Affinity. J. Virol. 2018, 92, e00692-18. [Google Scholar] [CrossRef]
- De Araújo, O.J.; Pinheiro, S.; Zamora, W.J.; Alves, C.N.; Lameira, J.; Lima, A.H. Structural, Energetic and Lipophilic Analysis of SARS-CoV-2 Non-Structural Protein 9 (NSP9). Sci. Rep. 2021, 11, 23003. [Google Scholar] [CrossRef]
- Littler, D.R.; Gully, B.S.; Colson, R.N.; Rossjohn, J. Crystal Structure of the SARS-CoV-2 Non-Structural Protein 9, Nsp9. iScience 2020, 23, 101258. [Google Scholar] [CrossRef] [PubMed]
- Rona, G.; Zeke, A.; Miwatani-Minter, B.; de Vries, M.; Kaur, R.; Schinlever, A.; Garcia, S.F.; Goldberg, H.V.; Wang, H.; Hinds, T.R.; et al. The NSP14/NSP10 RNA Repair Complex as a Pan-Coronavirus Therapeutic Target. Cell Death Differ. 2021, 29, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wu, L.; Shaw, N.; Gao, Y.; Wang, J.; Sun, Y.; Lou, Z.; Yan, L.; Zhang, R.; Rao, Z. Structural Basis and Functional Analysis of the SARS Coronavirus Nsp14-Nsp10 Complex. Proc. Natl. Acad. Sci. USA 2015, 112, 9436–9441. [Google Scholar] [CrossRef]
- Bouvet, M.; Lugari, A.; Posthuma, C.C.; Zevenhoven, J.C.; Bernard, S.; Betzi, S.; Imbert, I.; Canard, B.; Guillemot, J.C.; Lécine, P.; et al. Coronavirus Nsp10, a Critical Co-Factor for Activation of Multiple Replicative Enzymes. J. Biol. Chem. 2014, 289, 25783–25796. [Google Scholar] [CrossRef]
- Gadhave, K.; Kumar, P.; Kumar, A.; Bhardwaj, T.; Garg, N.; Giri, R. Conformational Dynamics of 13 Amino Acids Long NSP11 of SARS-CoV-2 under Membrane Mimetics and Different Solvent Conditions. Microb. Pathog. 2021, 158, 105041. [Google Scholar] [CrossRef]
- Elfiky, A.A. SARS-CoV-2 RNA Dependent RNA Polymerase (RdRp) Targeting: An in Silico Perspective. J. Biomol. Struct. Dyn. 2021, 39, 3204–3212. [Google Scholar] [CrossRef]
- Khan, M.T.; Irfan, M.; Ahsan, H.; Ahmed, A.; Kaushik, A.C.; Khan, A.S.; Chinnasamy, S.; Ali, A.; Wei, D.-Q. Structures of SARS-CoV-2 RNA-Binding Proteins and Therapeutic Targets. Intervirology 2021, 64, 55–68. [Google Scholar] [CrossRef]
- Mishra, A.; Rathore, A.S. RNA Dependent RNA Polymerase (RdRp) as a Drug Target for SARS-CoV2. J. Biomol. Struct. Dyn. 2022, 40, 6039–6051. [Google Scholar] [CrossRef]
- Yin, W.; Mao, C.; Luan, X.; Shen, D.D.; Shen, Q.; Su, H.; Wang, X.; Zhou, F.; Zhao, W.; Gao, M.; et al. Structural Basis for Inhibition of the RNA-Dependent RNA Polymerase from SARS-CoV-2 by Remdesivir. Science 2020, 368, 1499. [Google Scholar] [CrossRef]
- Wang, H.; Xue, S.; Yang, H.; Chen, C. Recent Progress in the Discovery of Inhibitors Targeting Coronavirus Proteases. Virol. Sin. 2016, 31, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, X.; Kong, F.; Hou, Y.J.; Wang, Q. Crucial Mutation in the Exoribonuclease Domain of Nsp14 of PEDV Leads to High Genetic Instability during Viral Replication. Cell Biosci. 2021, 11, 106. [Google Scholar] [CrossRef]
- Tahir, M. Coronavirus Genomic Nsp14-ExoN, Structure, Role, Mechanism, and Potential Application as a Drug Target. J. Med. Virol. 2021, 93, 4258–4264. [Google Scholar] [CrossRef]
- Krafcikova, P.; Silhan, J.; Nencka, R.; Boura, E. Structural Analysis of the SARS-CoV-2 Methyltransferase Complex Involved in RNA Cap Creation Bound to Sinefungin. Nat. Commun. 2020, 11, 3717. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. A Biochemical Perspective of the Nonstructural Proteins (NSPs) and the Spike Protein of SARS CoV-2. Protein J. 2021, 40, 260–295. [Google Scholar] [CrossRef] [PubMed]
- Vithani, N.; Ward, M.D.; Zimmerman, M.I.; Novak, B.; Borowsky, J.H.; Singh, S.; Bowman, G.R. SARS-CoV-2 Nsp16 Activation Mechanism and a Cryptic Pocket with Pan-Coronavirus Antiviral Potential. Biophys. J. 2021, 120, 2880–2889. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Lemus, M.; Minasov, G.; Shuvalova, L.; Inniss, N.L.; Kiryukhina, O.; Brunzelle, J.; Satchell, K.J.F. High-Resolution Structures of the SARS-CoV-2 2′-O-Methyltransferase Reveal Strategies for Structure-Based Inhibitor Design. Sci. Signal. 2020, 13, eabe1202. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef]
- Balka, K.R.; de Nardo, D. Understanding Early TLR Signaling through the Myddosome. J. Leukoc. Biol. 2019, 105, 339–351. [Google Scholar] [CrossRef]
- Satoh, T.; Akira, S. Toll-Like Receptor Signaling and Its Inducible Proteins. Microbiol. Spectr. 2016, 4, 4–6. [Google Scholar] [CrossRef]
- Barton, G.M.; Kagan, J.C. A Cell Biological View of Toll-like Receptor Function: Regulation through Compartmentalization. Nat. Rev. Immunol. 2009, 9, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Song, E.; Bai, B.; Vanhoutte, P.M. Toll-like Receptors Mediating Vascular Malfunction: Lessons from Receptor Subtypes. Pharmacol. Ther. 2016, 158, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Kagan, J.C. Signaling Organelles of the Innate Immune System. Cell 2012, 151, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Sato, S.; Mori, K.; Hoshino, K.; Takeuchi, O.; Takeda, K.; Akira, S. Cutting Edge: A Novel Toll/IL-1 Receptor Domain-Containing Adapter That Preferentially Activates the IFN-Beta Promoter in the Toll-like Receptor Signaling. J. Immunol. 2002, 169, 6668–6672. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The Role of Pattern-Recognition Receptors in Innate Immunity: Update on Toll-like Receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, C. Innate Recognition of Microbial-Derived Signals in Immunity and Inflammation. Sci. China Life Sci. 2016, 59, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like Receptors Activation, Signaling, and Targeting: An Overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef]
- Gao, W.; Xiong, Y.; Li, Q.; Yang, H. Inhibition of Toll-like Receptor Signaling as a Promising Therapy for Inflammatory Diseases: A Journey from Molecular to Nano Therapeutics. Front. Physiol. 2017, 8, 508. [Google Scholar] [CrossRef]
- Lushpa, V.A.; Goncharuk, M.V.; Lin, C.; Zalevsky, A.O.; Talyzina, I.A.; Luginina, A.P.; Vakhrameev, D.D.; Shevtsov, M.B.; Goncharuk, S.A.; Arseniev, A.S.; et al. Modulation of Toll-like Receptor 1 Intracellular Domain Structure and Activity by Zn2+ Ions. Commun. Biol. 2021, 4, 1003. [Google Scholar] [CrossRef]
- Takeuchi, O.; Sato, S.; Horiuchi, T.; Hoshino, K.; Takeda, K.; Dong, Z.; Modlin, R.L.; Akira, S. Cutting Edge: Role of Toll-Like Receptor 1 in Mediating Immune Response to Microbial Lipoproteins. J. Immunol. 2002, 169, 10–14. [Google Scholar] [CrossRef]
- Ahmed, S.; Moawad, M.; Elhefny, R.; Chest, M.A.-E.J. Is Toll like Receptor 4 a Common Pathway Hypothesis for Development of Lung Cancer and Idiopathic Pulmonary Fibrosis? Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The Role of TLR2 Ininfection and Immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zainol, M.I.B.; Kawasaki, T.; Monwan, W.; Murase, M.; Sueyoshi, T.; Kawai, T. Innate Immune Responses through Toll-like Receptor 3 Require Human-Antigen-R-Mediated Atp6v0d2 MRNA Stabilization. Sci. Rep. 2019, 9, 20406. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like Receptor 3 (TLR3) Regulation Mechanisms and Roles in Antiviral Innate Immune Responses. J. Zhejiang Univ. Sci. B 2021, 22, 609. [Google Scholar] [CrossRef]
- Molteni, M.; Gemma, S.; Rossetti, C. The Role of Toll-Like Receptor 4 in Infectious and Noninfectious Inflammation. Mediators Inflamm. 2016, 2016, 6978936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liang, X.; Bao, X.; Xiao, W.; Chen, G. Toll-like Receptor 4 (TLR4) Inhibitors: Current Research and Prospective. Eur. J. Med. Chem. 2022, 235, 114291. [Google Scholar] [CrossRef]
- Caballero, I.; Boyd, J.; Alminanã, C.; Sánchez-López, J.A.; Basatvat, S.; Montazeri, M.; Maslehat Lay, N.; Elliott, S.; Spiller, D.G.; White, M.R.H.; et al. Understanding the Dynamics of Toll-like Receptor 5 Response to Flagellin and Its Regulation by Estradiol. Sci. Rep. 2017, 7, 40981. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Sun, X.H.; Liu, X.; Xiao, Y.; Zhang, J.; Wang, T.; Chen, H.; Zhan, Y.Q.; Yu, M.; et al. Toll-like Receptor 5-Mediated Signaling Enhances Liver Regeneration in Mice. Mil. Med. Res. 2021, 8, 16. [Google Scholar] [CrossRef]
- Kim, J.H.; Kordahi, M.C.; Chac, D.; William DePaolo, R. Toll-like Receptor-6 Signaling Prevents Inflammation and Impacts Composition of the Microbiota during Inflammation-Induced Colorectal Cancer. Cancer Prev. Res. 2020, 13, 25–40. [Google Scholar] [CrossRef]
- De Almeida, L.A.; Macedo, G.C.; Marinho, F.A.V.; Gomes, M.T.R.; Corsetti, P.P.; Silva, A.M.; Cassataro, J.; Giambartolomei, G.H.; Oliveira, S.C. Toll-like Receptor 6 Plays an Important Role in Host Innate Resistance to Brucella Abortus Infection in Mice. Infect. Immun. 2013, 81, 1654–1662. [Google Scholar] [CrossRef]
- Bortolotti, D.; Gentili, V.; Rizzo, S.; Schiuma, G.; Beltrami, S.; Strazzabosco, G.; Fernandez, M.; Caccuri, F.; Caruso, A.; Rizzo, R. TLR3 and TLR7 RNA Sensor Activation during SARS-CoV-2 Infection. Microorganisms 2021, 9, 1820. [Google Scholar] [CrossRef]
- Macedo, A.B.; Novis, C.L.; de Assis, C.M.; Sorensen, E.S.; Moszczynski, P.; Huang, S.H.; Ren, Y.; Spivak, A.M.; Jones, R.B.; Planelles, V.; et al. Dual TLR2 and TLR7 Agonists as HIV Latency-Reversing Agents. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Marcken, M.; Dhaliwal, K.; Danielsen, A.C.; Gautron, A.S.; Dominguez-Villar, M. TLR7 and TLR8 Activate Distinct Pathways in Monocytes during RNA Virus Infection. Sci. Signal. 2019, 12, eaaw1347. [Google Scholar] [CrossRef] [PubMed]
- Salvi, V.; Nguyen, H.O.; Sozio, F.; Schioppa, T.; Gaudenzi, C.; Laffranchi, M.; Scapini, P.; Passari, M.; Barbazza, I.; Tiberio, L.; et al. SARS-CoV-2-Associated SsRNAs Activate Inflammation and Immunity via TLR7/8. JCI Insight 2021, 6, e150542. [Google Scholar] [CrossRef] [PubMed]
- Costa, T.J.; Potje, S.R.; Fraga-Silva, T.F.C.; da Silva-Neto, J.A.; Barros, P.R.; Rodrigues, D.; Machado, M.R.; Martins, R.B.; Santos-Eichler, R.A.; Benatti, M.N.; et al. Mitochondrial DNA and TLR9 Activation Contribute to SARS-CoV-2-Induced Endothelial Cell Damage. Vascul. Pharmacol. 2022, 142, 106946. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.S.; Lukason, D.P.; Feliu, M.; Ward, R.A.; Lord, A.K.; Reedy, J.L.; Ramirez-Ortiz, Z.G.; Tam, J.M.; Kasperkovitz, P.V.; Negoro, P.E.; et al. CD82 Controls CpG-Dependent TLR9 Signaling. FASEB J. 2019, 33, 12500–12514. [Google Scholar] [CrossRef]
- Lester, S.N.; Li, K. Toll-Like Receptors in Antiviral Innate Immunity. J. Mol. Biol. 2014, 426, 1246. [Google Scholar] [CrossRef]
- Gaglia, M.M. Anti-Viral and pro-Inflammatory Functions of Toll-like Receptors during Gamma-Herpesvirus Infections. Virol. J. 2021, 18, 218. [Google Scholar] [CrossRef]
- Shah, M.; Anwar, M.A.; Kim, J.H.; Choi, S. Advances in Antiviral Therapies Targeting Toll-like Receptors. Expert Opin. Investig. Drugs 2016, 25, 437–453. [Google Scholar] [CrossRef]
- Vijay, K. Toll-like Receptors in Immunity and Inflammatory Diseases: Past, Present, and Future. Int. Immunopharmacol. 2018, 59, 391. [Google Scholar] [CrossRef]
- Federico, S.; Pozzetti, L.; Papa, A.; Carullo, G.; Gemma, S.; Butini, S.; Campiani, G.; Relitti, N. Modulation of the Innate Immune Response by Targeting Toll-like Receptors: A Perspective on Their Agonists and Antagonists. J. Med. Chem. 2020, 63, 13466–13513. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Hennessy, E.J.; Parker, A.E. Targeting Toll-like Receptors: Emerging Therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef]
- Zheng, W.; Xu, Q.; Zhang, Y.; Xiaofei, E.; Gao, W.; Zhang, M.; Zhai, W.; Rajkumar, R.S.; Liu, Z. Toll-like Receptor-Mediated Innate Immunity against Herpesviridae Infection: A Current Perspective on Viral Infection Signaling Pathways. Virol. J. 2020, 17, 192. [Google Scholar] [CrossRef]
- Boehme, K.W.; Compton, T. Innate Sensing of Viruses by Toll-Like Receptors. J. Virol. 2004, 78, 7867. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Mansell, A.; Ussher, J.E.; Brooks, A.E.S.; Manning, K.; Wang, C.J.H.; Taylor, J.A. Rotavirus NSP4 Triggers Secretion of Proinflammatory Cytokines from Macrophages via Toll-Like Receptor 2. J. Virol. 2013, 87, 11160–11167. [Google Scholar] [CrossRef]
- Dela Justina, V.; Giachini, F.R.; Priviero, F.; Webb, R.C. COVID-19 and Hypertension: Is There a Role for DsRNA and Activation of Toll-like Receptor 3? Vascul. Pharmacol. 2021, 140, 106861. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.F.; Tang, L.H.; Ooi, E.E. A TLR3 Ligand That Exhibits Potent Inhibition of Influenza Virus Replication and Has Strong Adjuvant Activity Has the Potential for Dual Applications in an Influenza Pandemic. Vaccine 2009, 27, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.L.; Meng, T.C.; Tomai, M.A. The Antiviral Activity of Toll-like Receptor 7 and 7/8 Agonists. Drug News Perspect. 2008, 21, 69–87. [Google Scholar] [CrossRef]
- Rose, W.A.; McGowin, C.L.; Pyles, R.B. FSL-1, a Bacterial-Derived Toll-like Receptor 2/6 Agonist, Enhances Resistance to Experimental HSV-2 Infection. Virol. J. 2009, 6, 195. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral Evasion and Subversion of Pattern-Recognition Receptor Signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef]
- Duan, S.; Xu, X.; Wang, J.; Huang, L.; Peng, J.; Yu, T.; Zhou, Y.; Cheng, K.; Liu, S. TLR1/2 Agonist Enhances Reversal of HIV-1 Latency and Promotes NK Cell-Induced Suppression of HIV-1-Infected Autologous CD4 + T Cells. J. Virol. 2021, 95, e00816-21. [Google Scholar] [CrossRef]
- Girkin, J.; Loo, S.L.; Esneau, C.; Maltby, S.; Mercuri, F.; Chua, B.; Reid, A.T.; Veerati, P.C.; Grainge, C.L.; Wark, P.A.B.; et al. TLR2-Mediated Innate Immune Priming Boosts Lung Anti-Viral Immunity. Eur. Respir. J. 2021, 58, 2001584. [Google Scholar] [CrossRef] [PubMed]
- Howell, M.D.; Gallo, R.L.; Boguniewicz, M.; Jones, J.F.; Wong, C.; Streib, J.E.; Leung, D.Y.M. Cytokine Milieu of Atopic Dermatitis Skin Subverts the Innate Immune Response to Vaccinia Virus. Immunity 2006, 24, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Hutchens, M.A.; Luker, K.E.; Sonstein, J.; Núñez, G.; Curtis, J.L.; Luker, G.D. Protective Effect of Toll-like Receptor 4 in Pulmonary Vaccinia Infection. PLoS Pathog. 2008, 4, e1000153. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of Double-Stranded RNA and Activation of NF-KappaB by Toll-like Receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Mazaleuskaya, L.; Veltrop, R.; Ikpeze, N.; Martin-Garcia, J.; Navas-Martin, S. Protective Role of Toll-like Receptor 3-Induced Type I Interferon in Murine Coronavirus Infection of Macrophages. Viruses 2012, 4, 901–923. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, X.; Liu, M.; Hu, Q.; Song, L.; Ye, L.; Zhou, D.; Ho, W. A Critical Function of Toll-like Receptor-3 in the Induction of Anti-Human Immunodeficiency Virus Activities in Macrophages. Immunology 2010, 131, 40–49. [Google Scholar] [CrossRef]
- Isogawa, M.; Robek, M.D.; Furuichi, Y.; Chisari, F.v. Toll-like Receptor Signaling Inhibits Hepatitis B Virus Replication in Vivo. J. Virol. 2005, 79, 7269–7272. [Google Scholar] [CrossRef]
- Guillot, L.; le Goffic, R.; Bloch, S.; Escriou, N.; Akira, S.; Chignard, M.; Si-Tahar, M. Involvement of Toll-like Receptor 3 in the Immune Response of Lung Epithelial Cells to Double-Stranded RNA and Influenza A Virus. J. Biol. Chem. 2005, 280, 5571–5580. [Google Scholar] [CrossRef]
- Zhang, M.; Yan, Z.; Wang, J.; Yao, X. Toll-like Receptors 7 and 8 Expression Correlates with the Expression of Immune Biomarkers and Positively Predicts the Clinical Outcome of Patients with Melanoma. Onco Targets Ther. 2017, 10, 4339. [Google Scholar] [CrossRef]
- Drake, M.G.; Evans, S.E.; Dickey, B.F.; Fryer, A.D.; Jacoby, D.B. Toll-like Receptor-2/6 and Toll-like Receptor-9 Agonists Suppress Viral Replication but Not Airway Hyperreactivity in Guinea Pigs. Am J Respir Cell Mol. Biol. 2013, 48, 790–796. [Google Scholar] [CrossRef]
- Proud, P.C.; Tsitoura, D.; Watson, R.J.; Chua, B.Y.; Aram, M.J.; Bewley, K.R.; Cavell, B.E.; Cobb, R.; Dowall, S.; Fotheringham, S.A.; et al. Prophylactic Intranasal Administration of a TLR2/6 Agonist Reduces Upper Respiratory Tract Viral Shedding in a SARS-CoV-2 Challenge Ferret Model. EBioMedicine 2021, 63, 103153. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.F.; Tang, L.H.; Ooi, E.E.; Subbarao, K. Activation of the Innate Immune System Provides Broad-Spectrum Protection against Influenza A Viruses with Pandemic Potential in Mice. Virology 2010, 406, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Christopher, M.; Wong, J. Use of Toll-Like Receptor 3 Agonists Against Respiratory Viral Infections. Antiinflamm. Antiallergy Agents Med. Chem. 2011, 10, 327. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.E.; Cho, H.; Jung, H.G.; Lee, W.; Seo, H.Y.; Lee, S.H.; Ahn, D.G.; Kim, S.J.; Yu, J.W.; et al. Antiviral Efficacy of Orally Delivered Neoagarohexaose, a Nonconventional TLR4 Agonist, against Norovirus Infection in Mice. Biomaterials 2020, 263, 120391. [Google Scholar] [CrossRef] [PubMed]
- Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. [Google Scholar] [CrossRef]
- Georgel, A.F.; Cayet, D.; Pizzorno, A.; Rosa-Calatrava, M.; Paget, C.; Sencio, V.; Dubuisson, J.; Trottein, F.; Sirard, J.C.; Carnoy, C. Toll-like Receptor 5 Agonist Flagellin Reduces Influenza A Virus Replication Independently of Type I Interferon and Interleukin 22 and Improves Antiviral Efficacy of Oseltamivir. Antiviral Res. 2019, 168, 28–35. [Google Scholar] [CrossRef]
- Melin, N.; Sánchez-Taltavull, D.; Fahrner, R.; Keogh, A.; Dosch, M.; Büchi, I.; Zimmer, Y.; Medová, M.; Beldi, G.; Aebersold, D.M.; et al. Synergistic Effect of the TLR5 Agonist CBLB502 and Its Downstream Effector IL-22 against Liver Injury. Cell Death Dis. 2021, 12, 366. [Google Scholar] [CrossRef]
- SenGupta, D.; Brinson, C.; DeJesus, E.; Mills, A.; Shalit, P.; Guo, S.; Cai, Y.; Wallin, J.J.; Zhang, L.; Humeniuk, R.; et al. The TLR7 Agonist Vesatolimod Induced a Modest Delay in Viral Rebound in HIV Controllers after Cessation of Antiretroviral Therapy. Sci. Transl. Med. 2021, 13, eabg3071. [Google Scholar] [CrossRef]
- Vanwalscappel, B.; Tada, T.; Landau, N.R. Toll-like Receptor Agonist R848 Blocks Zika Virus Replication by Inducing the Antiviral Protein Viperin. Virology 2018, 522, 199–208. [Google Scholar] [CrossRef]
- Amin, O.E.; Colbeck, E.J.; Daffis, S.; Khan, S.; Ramakrishnan, D.; Pattabiraman, D.; Chu, R.; Micolochick Steuer, H.; Lehar, S.; Peiser, L.; et al. Therapeutic Potential of TLR8 Agonist GS-9688 (Selgantolimod) in Chronic Hepatitis B: Remodeling of Antiviral and Regulatory Mediators. Hepatology 2021, 74, 55–71. [Google Scholar] [CrossRef]
- Manik, M.; Singh, R.K. Role of Toll-like Receptors in Modulation of Cytokine Storm Signaling in SARS-CoV-2-induced COVID-19. J. Med. Virol. 2022, 94, 869. [Google Scholar] [CrossRef] [PubMed]
- Mabrey, F.L.; Morrell, E.D.; Wurfel, M.M. TLRs in COVID-19: How They Drive Immunopathology and the Rationale for Modulation. Innate Immun. 2021, 27, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Sanchez, D.J.; Aliyari, R.; Lu, S.; Cheng, G. Systematic Identification of Type I and Type II Interferon-Induced Antiviral Factors. Proc. Natl. Acad. Sci. USA 2012, 109, 4239–4244. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- Zanoni, I.; Granucci, F.; Broggi, A. Interferon (IFN)-λ Takes the Helm: Immunomodulatory Roles of Type III IFNs. Front. Immunol. 2017, 8, 1661. [Google Scholar] [CrossRef]
- Lotfi, M.; Rezaei, N. SARS-CoV-2: A Comprehensive Review from Pathogenicity of the Virus to Clinical Consequences. J. Med. Virol. 2020, 92, 1864. [Google Scholar] [CrossRef]
- Teijaro, J.R. Type I Interferons in Viral Control and Immune Regulation. Curr. Opin. Virol. 2016, 16, 31–40. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Al-Ahmed, S.H.; Muhammad, J.; Khan, A.; Sule, A.A.; Tirupathi, R.; Al Mutair, A.; Alhumaid, S.; Al-Omari, A.; Dhawan, M.; et al. Role of Inflammatory Cytokines in COVID-19 Patients: A Review on Molecular Mechanisms, Immune Functions, Immunopathology and Immunomodulatory Drugs to Counter Cytokine Storm. Vaccines 2021, 9, 436. [Google Scholar] [CrossRef]
- Darif, D.; Hammi, I.; Kihel, A.; el Idrissi Saik, I.; Guessous, F.; Akarid, K. The Pro-Inflammatory Cytokines in COVID-19 Pathogenesis: What Goes Wrong? Microb. Pathog. 2021, 153, 104799. [Google Scholar] [CrossRef]
- Yapasert, R.; Khaw-On, P.; Banjerdpongchai, R. Coronavirus Infection-Associated Cell Death Signaling and Potential Therapeutic Targets. Molecules 2021, 26, 7459. [Google Scholar] [CrossRef]
- Alesci, A.; Aragona, M.; Cicero, N.; Lauriano, E.R. Can Nutraceuticals Assist Treatment and Improve COVID-19 Symptoms? Nat. Prod. Res. 2022, 36, 2672–2691. [Google Scholar] [CrossRef] [PubMed]
- Domi, E.; Hoxha, M.; Kolovani, E.; Tricarico, D.; Zappacosta, B. The Importance of Nutraceuticals in COVID-19: What’s the Role of Resveratrol? Molecules 2022, 27, 2376. [Google Scholar] [CrossRef] [PubMed]
- Parisi, G.F.; Carota, G.; Castruccio Castracani, C.; Spampinato, M.; Manti, S.; Papale, M.; di Rosa, M.; Barbagallo, I.; Leonardi, S. Nutraceuticals in the Prevention of Viral Infections, Including COVID-19, among the Pediatric Population: A Review of the Literature. Int. J. Mol. Sci. 2021, 22, 52465. [Google Scholar] [CrossRef]
- Ramdani, L.H.; Bachari, K. Potential Therapeutic Effects of Resveratrol against SARS-CoV-2. Acta Virol. 2020, 64, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Maffioli, P.; D’Angelo, A.; di Pierro, F. A Role for Quercetin in Coronavirus Disease 2019 (COVID-19). Phytother. Res. 2021, 35, 1230–1236. [Google Scholar] [CrossRef]
- Fan, W.; Qian, S.; Qian, P.; Li, X. Antiviral Activity of Luteolin against Japanese Encephalitis Virus. Virus Res. 2016, 220, 112–116. [Google Scholar] [CrossRef]
- Hao, L.; Shan, Q.; Wei, J.; Ma, F.; Sun, P. Lactoferrin: Major Physiological Functions and Applications. Curr. Protein Pept. Sci. 2019, 20, 139–144. [Google Scholar] [CrossRef]
- Haggag, Y.A.; El-Ashmawy, N.E.; Okasha, K.M. Is Hesperidin Essential for Prophylaxis and Treatment of COVID-19 Infection? Med. Hypotheses 2020, 144, 109957. [Google Scholar] [CrossRef]
- Luo, P.; Liu, D.; Li, J. Pharmacological Perspective: Glycyrrhizin May Be an Efficacious Therapeutic Agent for COVID-19. Int. J. Antimicrob. Agents 2020, 55, 105995. [Google Scholar] [CrossRef]
- Babaei, F.; Nassiri-Asl, M.; Hosseinzadeh, H. Curcumin (a Constituent of Turmeric): New Treatment Option against COVID-19. Food Sci. Nutr. 2020, 8, 5215–5227. [Google Scholar] [CrossRef]
- Pasandi Pour, A.; Farahbakhsh, H. Lawsonia Inermis L. Leaves Aqueous Extract as a Natural Antioxidant and Antibacterial Product. Nat. Prod. Res. 2020, 34, 3399–3403. [Google Scholar] [CrossRef] [PubMed]
- Davoudi-Monfared, E.; Rahmani, H.; Khalili, H.; Hajiabdolbaghi, M.; Salehi, M.; Abbasian, L.; Kazemzadeh, H.; Yekaninejad, M.S. A Randomized Clinical Trial of the Efficacy and Safety of Interferon β-1a in Treatment of Severe COVID-19. Antimicrob. Agents Chemother. 2020, 64, e01061-20. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, V.; Shannon, C.P.; Wei, X.S.; Xiang, X.; Wang, X.; Wang, Z.H.; Tebbutt, S.J.; Kollmann, T.R.; Fish, E.N. Interferon-A2b Treatment for COVID-19. Front. Immunol. 2020, 11, 1061. [Google Scholar] [CrossRef] [PubMed]
- Hung, I.F.N.; Lung, K.C.; Tso, E.Y.K.; Liu, R.; Chung, T.W.H.; Chu, M.Y.; Ng, Y.Y.; Lo, J.; Chan, J.; Tam, A.R.; et al. Triple Combination of Interferon Beta-1b, Lopinavir-Ritonavir, and Ribavirin in the Treatment of Patients Admitted to Hospital with COVID-19: An Open-Label, Randomised, Phase 2 Trial. Lancet 2020, 395, 1695–1704. [Google Scholar] [CrossRef]
- Barbalat, R.; Lau, L.; Locksley, R.M.; Barton, G.M. Toll-like Receptor 2 on Inflammatory Monocytes Induces Type I Interferon in Response to Viral but Not Bacterial Ligands. Nat. Immunol. 2009, 10, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef]
- Menachery, V.D.; Debbink, K.; Baric, R.S. Coronavirus Non-Structural Protein 16: Evasion, Attenuation, and Possible Treatments. Virus Res. 2014, 194, 191–199. [Google Scholar] [CrossRef]
- Bouvet, M.; Debarnot, C.; Imbert, I.; Selisko, B.; Snijder, E.J.; Canard, B.; Decroly, E. In Vitro Reconstitution of SARS-Coronavirus MRNA Cap Methylation. PLoS Pathog. 2010, 6, e1000863. [Google Scholar] [CrossRef]
- De Rivero Vaccari, J.C.; Dietrich, W.D.; Keane, R.W.; de Rivero Vaccari, J.P. The Inflammasome in Times of COVID-19. Front. Immunol. 2020, 11, 583373. [Google Scholar] [CrossRef]
- Sariol, A.; Perlman, S. SARS-CoV-2 Takes Its Toll. Nat. Immunol. 2021, 22, 801–802. [Google Scholar] [CrossRef]
- Zheng, M.; Karki, R.; Williams, E.P.; Yang, D.; Fitzpatrick, E.; Vogel, P.; Jonsson, C.B.; Kanneganti, T.D. TLR2 Senses the SARS-CoV-2 Envelope Protein to Produce Inflammatory Cytokines. Nat. Immunol. 2021, 22, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhao, J.; Perlman, S. Autocrine Interferon Priming in Macrophages but Not Dendritic Cells Results in Enhanced Cytokine and Chemokine Production after Coronavirus Infection. mBio 2010, 1, e00219-10. [Google Scholar] [CrossRef] [PubMed]
- Khanmohammadi, S.; Rezaei, N. Role of Toll-like Receptors in the Pathogenesis of COVID-19. J. Med. Virol. 2021, 93, 2735. [Google Scholar] [CrossRef] [PubMed]
- Totura, A.L.; Whitmore, A.; Agnihothram, S.; Schäfer, A.; Katze, M.G.; Heise, M.T.; Baric, R.S. Toll-Like Receptor 3 Signaling via TRIF Contributes to a Protective Innate Immune Response to Severe Acute Respiratory Syndrome Coronavirus Infection. mBio 2015, 6, e00638-15. [Google Scholar] [CrossRef]
- Cicco, S.; Cicco, G.; Racanelli, V.; Vacca, A. Neutrophil Extracellular Traps (NETs) and Damage-Associated Molecular Patterns (DAMPs): Two Potential Targets for COVID-19 Treatment. Mediators Inflamm. 2020, 2020, 7527953. [Google Scholar] [CrossRef]
- Khadke, S.; Ahmed, N.; Ahmed, N.; Ratts, R.; Raju, S.; Gallogly, M.; de Lima, M.; Sohail, M.R. Harnessing the Immune System to Overcome Cytokine Storm and Reduce Viral Load in COVID-19: A Review of the Phases of Illness and Therapeutic Agents. Virol. J. 2020, 17, 154. [Google Scholar] [CrossRef]
- Cao, C.; Yin, C.; Shou, S.; Wang, J.; Yu, L.; Li, X.; Chai, Y. Ulinastatin Protects Against LPS-Induced Acute Lung Injury By Attenuating TLR4/NF-ΚB Pathway Activation and Reducing Inflammatory Mediators. Shock 2018, 50, 595–605. [Google Scholar] [CrossRef]
- Ohashi, K.; Burkart, V.; Flohé, S.; Kolb, H. Cutting Edge: Heat Shock Protein 60 Is a Putative Endogenous Ligand of the Toll-like Receptor-4 Complex. J. Immunol. 2000, 164, 558–561. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Bhattacharya, M.; Sharma, G.; Lee, S.S.; Agoramoorthy, G. Consider TLR5 for New Therapeutic Development against COVID-19. J. Med. Virol. 2020, 92, 2314–2315. [Google Scholar] [CrossRef]
- Magro, G. SARS-CoV-2 and COVID-19: Is Interleukin-6 (IL-6) the “culprit Lesion” of ARDS Onset? What Is There besides Tocilizumab? SGP130Fc. Cytokine X 2020, 2, 100029. [Google Scholar] [CrossRef]
- Su, H.; Lei, C.T.; Zhang, C. Interleukin-6 Signaling Pathway and Its Role in Kidney Disease: An Update. Front. Immunol. 2017, 8, 405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, T.K.; Mickelson, D.J.; Fink, J.; Solberg, J.C.; Inglefield, J.R.; Hook, D.; Gupta, S.K.; Gibson, S.; Alkan, S.S. Toll-like Receptor (TLR) 2-9 Agonists-Induced Cytokines and Chemokines: I. Comparison with T Cell Receptor-Induced Responses. Cell Immunol. 2006, 243, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Eutimio, M.A.; López-Macías, C.; Pastelin-Palacios, R. Bioinformatic Analysis and Identification of Single-Stranded RNA Sequences Recognized by TLR7/8 in the SARS-CoV-2, SARS-CoV, and MERS-CoV Genomes. Microbes Infect. 2020, 22, 226–229. [Google Scholar] [CrossRef]
- Van der Made, C.I.; Simons, A.; Schuurs-Hoeijmakers, J.; van den Heuvel, G.; Mantere, T.; Kersten, S.; van Deuren, R.C.; Steehouwer, M.; van Reijmersdal, S.V.; Jaeger, M.; et al. Presence of Genetic Variants Among Young Men with Severe COVID-19. JAMA 2020, 324, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, A.; Alexandris, N.; Konstantinou, E.; Mesiakaris, K.; Zanidis, C.; Farsalinos, K.; Poulas, K. Imiquimod-A Toll like Receptor 7 Agonist-Is an Ideal Option for Management of COVID 19. Environ. Res. 2020, 188, 109858. [Google Scholar] [CrossRef] [PubMed]
- Jangra, S.; de Vrieze, J.; Choi, A.; Rathnasinghe, R.; Laghlali, G.; Uvyn, A.; van Herck, S.; Nuhn, L.; Deswarte, K.; Zhong, Z.; et al. Sterilizing Immunity against SARS-CoV-2 Infection in Mice by a Single-Shot and Lipid Amphiphile Imidazoquinoline TLR7/8 Agonist-Adjuvanted Recombinant Spike Protein Vaccine. Angew. Chem. Int. Ed. Engl. 2021, 60, 9467–9473. [Google Scholar] [CrossRef]
- Ma-Lauer, Y.; Carbajo-Lozoya, J.; Hein, M.Y.; Müller, M.A.; Deng, W.; Lei, J.; Meyer, B.; Kusov, Y.; von Brunn, B.; Bairad, D.R.; et al. P53 Down-Regulates SARS Coronavirus Replication and Is Targeted by the SARS-Unique Domain and PLpro via E3 Ubiquitin Ligase RCHY1. Proc. Natl. Acad. Sci. USA 2016, 113, E5192–E5201. [Google Scholar] [CrossRef]
- Fekete, T.; Ágics, B.; Bencze, D.; Bene, K.; Szántó, A.; Tarr, T.; Veréb, Z.; Bácsi, A.; Pázmándi, K. Regulation of RLR-Mediated Antiviral Responses of Human Dendritic Cells by MTOR. Front. Immunol. 2020, 11, 2326. [Google Scholar] [CrossRef]
- Khan, S.; Shafiei, M.S.; Longoria, C.; Schoggins, J.W.; Savani, R.C.; Zaki, H. SARS-CoV-2 Spike Protein Induces Inflammation via TLR2-Dependent Activation of the NF-ΚB Pathway. Elife 2021, 10. [Google Scholar] [CrossRef]
- Durai, P.; Shin, H.-J.; Achek, A.; Kwon, H.-K.; Govindaraj, R.G.; Panneerselvam, S.; Yesudhas, D.; Choi, J.; No, K.T.; Choi, S. Toll-like Receptor 2 Antagonists Identified through Virtual Screening and Experimental Validation. FEBS J. 2017, 284, 2264–2283. [Google Scholar] [CrossRef]
- Menezes, M.C.S.; Veiga, A.D.M.; Martins de Lima, T.; Kunimi Kubo Ariga, S.; Vieira Barbeiro, H.; de Lucena Moreira, C.; Pinto, A.A.S.; Brandao, R.A.; Marchini, J.F.; Alencar, J.C.; et al. Lower Peripheral Blood Toll-like Receptor 3 Expression Is Associated with an Unfavorable Outcome in Severe COVID-19 Patients. Sci. Rep. 2021, 11, 15223. [Google Scholar] [CrossRef] [PubMed]
- Gosu, V.; Son, S.; Shin, D.; Song, K.-D. Insights into the Dynamic Nature of the DsRNA-Bound TLR3 Complex. Sci. Rep. 2019, 9, 3652. [Google Scholar] [CrossRef] [PubMed]
- Aboudounya, M.M.; Heads, R.J. COVID-19 and Toll-Like Receptor 4 (TLR4): SARS-CoV-2 May Bind and Activate TLR4 to Increase ACE2 Expression, Facilitating Entry and Causing Hyperinflammation. Mediators Inflamm. 2021, 2021, 8874339. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, M.; Choi, J.; Jang, A.; Choi, S.; Yeon, J.; Jang, M.; Lee, Y.; Son, K.; Shin, S.Y.; Jeong, M.S.; et al. Molecular Mechanism Underlying the TLR4 Antagonistic and Antiseptic Activities of Papiliocin, an Insect Innate Immune Response Molecule. Proc. Natl. Acad. Sci. USA 2022, 119, e2115669119. [Google Scholar] [CrossRef]
- Dyavar, S.R.; Singh, R.; Emani, R.; Pawar, G.P.; Chaudhari, V.D.; Podany, A.T.; Avedissian, S.N.; Fletcher, C.V.; Salunke, D.B. Role of Toll-like Receptor 7/8 Pathways in Regulation of Interferon Response and Inflammatory Mediators during SARS-CoV2 Infection and Potential Therapeutic Options. Biomed. Pharmacother. 2021, 141, 111794. [Google Scholar] [CrossRef]
- Gentile, F.; Deriu, M.; Licandro, G.; Prunotto, A.; Danani, A.; Tuszynski, J. Structure Based Modeling of Small Molecules Binding to the TLR7 by Atomistic Level Simulations. Molecules 2015, 20, 8316–8340. [Google Scholar] [CrossRef]
- Zhang, Z.; Ohto, U.; Shibata, T.; Krayukhina, E.; Taoka, M.; Yamauchi, Y.; Tanji, H.; Isobe, T.; Uchiyama, S.; Miyake, K.; et al. Structural Analysis Reveals That Toll-like Receptor 7 Is a Dual Receptor for Guanosine and Single-Stranded RNA. Immunity 2016, 45, 737–748. [Google Scholar] [CrossRef]
- Yang, C.W.; Chen, M.F. Low Compositions of Human Toll-like Receptor 7/8-Stimulating RNA Motifs in the MERS-CoV, SARS-CoV and SARS-CoV-2 Genomes Imply a Substantial Ability to Evade Human Innate Immunity. PeerJ 2021, 9, e11008. [Google Scholar] [CrossRef]
- Mubarak, A.; Alturaiki, W.; Hemida, M.G. Middle East Respiratory Syndrome Coronavirus (MERS-CoV): Infection, Immunological Response, and Vaccine Development. J. Immunol. Res. 2019, 2019, 6491738. [Google Scholar] [CrossRef]
- Durai, P.; Batool, M.; Shah, M.; Choi, S. Middle East Respiratory Syndrome Coronavirus: Transmission, Virology and Therapeutic Targeting to Aid in Outbreak Control. Exp. Mol. Med. 2015, 47, e181. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fehr, A.R.; Zheng, J.; Wohlford-Lenane, C.; Abrahante, J.E.; Mack, M.; Sompallae, R.; McCray, P.B.; Meyerholz, D.K.; Perlman, S. IFN-I Response Timing Relative to Virus Replication Determines MERS Coronavirus Infection Outcomes. J. Clin. Investig. 2019, 129, 3625–3639. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Barragan, L.; Züst, R.; Weber, F.; Spiegel, M.; Lang, K.S.; Akira, S.; Thiel, V.; Ludewig, B. Control of Coronavirus Infection through Plasmacytoid Dendritic-Cell-Derived Type I Interferon. Blood 2007, 109, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Gralinski, L.E.; Menachery, V.D. Return of the Coronavirus: 2019-NCoV. Viruses 2020, 12, 135. [Google Scholar] [CrossRef]
- Wondmkun, Y.T.; Mohammed, O.A. A Review on Novel Drug Targets and Future Directions for COVID-19 Treatment. Biologics 2020, 14, 77–82. [Google Scholar] [CrossRef]
- Hussein, R.A.; El-Anssary, A.A. Plants Secondary Metabolites: The Key Drivers of the Pharmacological Actions of Medicinal Plants. Herb. Med. 2018. [Google Scholar] [CrossRef]
- Pour, P.M.; Fakhri, S.; Asgary, S.; Farzaei, M.H.; Echeverría, J. The Signaling Pathways, and Therapeutic Targets of Antiviral Agents: Focusing on the Antiviral Approaches and Clinical Perspectives of Anthocyanins in the Management of Viral Diseases. Front. Pharmacol. 2019, 10, 1207. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Kärenlampi, S.O.; Heinonen, I.M.; Mykkänen, H.M.; Törronen, A.R. Content of the Flavonols Quercetin, Myricetin, and Kaempferol in 25 Edible Berries. J. Agric. Food Chem. 1999, 47, 2274–2279. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, R.; Shohag, S.; Hossain, E.; Shah, M.; Shuvo, S.; Khan, H.; Chowdhury, A.R.; Bulbul, I.J.; Hossain, S.; et al. Multifaceted Role of Natural Sources for COVID-19 Pandemic as Marine Drugs. Environ. Sci. Pollut. Res. Int. 2022, 29, 46527–46550. [Google Scholar] [CrossRef]
- Hwang, S.S.; Lim, J.; Yu, Z.; Kong, P.; Sefik, E.; Xu, H.; Harman, C.C.D.; Kim, L.K.; Lee, G.R.; Li, H.B.; et al. Cryo-EM Structure of the 2019-NCoV Spike in the Prefusion Conformation. Science 2020, 367, 1255–1260. [Google Scholar] [CrossRef]
- Mody, V.; Ho, J.; Wills, S.; Mawri, A.; Lawson, L.; Ebert, M.C.C.J.C.; Fortin, G.M.; Rayalam, S.; Taval, S. Identification of 3-Chymotrypsin like Protease (3CLPro) Inhibitors as Potential Anti-SARS-CoV-2 Agents. Commun. Biol. 2021, 4, 93. [Google Scholar] [CrossRef]
- Mamedov, T.; Yuksel, D.; Ilgın, M.; Gürbüzaslan, I.; Gulec, B.; Mammadova, G.; Ozdarendeli, A.; Yetiskin, H.; Kaplan, B.; Islam Pavel, S.T.; et al. Production and Characterization of Nucleocapsid and Rbd Cocktail Antigens of SARS–CoV-2 in Nicotiana Benthamiana Plant as a Vaccine Candidate against COVID-19. Vaccines 2021, 9, 1337. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.M.; Yang, W.L.; Yang, F.Y.; Zhang, L.; Huang, W.J.; Hou, W.; Fan, C.F.; Jin, R.H.; Feng, Y.M.; Wang, Y.C.; et al. Cathepsin L Plays a Key Role in SARS-CoV-2 Infection in Humans and Humanized Mice and Is a Promising Target for New Drug Development. Signal Transduct. Target. Ther. 2021, 6, 134. [Google Scholar] [CrossRef] [PubMed]
- Hippisley-Cox, J.; Young, D.; Coupland, C.; Channon, K.M.; Tan, P.S.; Harrison, D.A.; Rowan, K.; Aveyard, P.; Pavord, I.D.; Watkinson, P.J. Risk of Severe COVID-19 Disease with ACE Inhibitors and Angiotensin Receptor Blockers: Cohort Study Including 8.3 million People. Heart 2020, 106, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Vaduganathan, M.; Vardeny, O.; Michel, T.; McMurray, J.J.V.; Pfeffer, M.A.; Solomon, S.D. Renin–Angiotensin–Aldosterone System Inhibitors in Patients with COVID-19. N. Engl. J. Med. 2020, 382, 1653–1659. [Google Scholar] [CrossRef]
- Bombardini, T.; Picano, E. Angiotensin-Converting Enzyme 2 as the Molecular Bridge Between Epidemiologic and Clinical Features of COVID-19. Can. J. Cardiol. 2020, 36, 784.e1–784.e2. [Google Scholar] [CrossRef]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.M.; Rottier, P.J.M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef]
- Zhang, H.; Penninger, J.M.; Li, Y.; Zhong, N.; Slutsky, A.S. Angiotensin-Converting Enzyme 2 (ACE2) as a SARS-CoV-2 Receptor: Molecular Mechanisms and Potential Therapeutic Target. Intensive Care Med. 2020, 46, 586–590. [Google Scholar] [CrossRef]
- Chatterjee, B.; Thakur, S.S. ACE2 as a Potential Therapeutic Target for Pandemic COVID-19. RSC Adv. 2020, 10, 39808–39813. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, Z.; Li, Y.; Huang, W.; Zhou, M.; Zhang, X.; Jiang, W. Angiotensin-Converting Enzyme Inhibition Attenuates Lipopolysaccharide-Induced Lung Injury by Regulating the Balance between Angiotensin-Converting Enzyme and Angiotensin-Converting Enzyme 2 and Inhibiting Mitogen-Activated Protein Kinase Activation. Shock 2015, 43, 395–404. [Google Scholar] [CrossRef]
- Soler, M.J.; Ye, M.; Wysocki, J.; William, J.; Lloveras, J.; Batlle, D. Localization of ACE2 in the Renal Vasculature: Amplification by Angiotensin II Type 1 Receptor Blockade Using Telmisartan. Am. J. Physiol. Renal Physiol. 2009, 296, 398–405. [Google Scholar] [CrossRef]
- Wang, X.; Ye, Y.; Gong, H.; Wu, J.; Yuan, J.; Wang, S.; Yin, P.; Ding, Z.; Kang, L.; Jiang, Q.; et al. The Effects of Different Angiotensin II Type 1 Receptor Blockers on the Regulation of the ACE-AngII-AT1 and ACE2-Ang(1–7)-Mas Axes in Pressure Overload-Induced Cardiac Remodeling in Male Mice. J. Mol. Cell Cardiol. 2016, 97, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I.; Pawara, R.; Surana, S.; Patel, H. The Repurposed ACE2 Inhibitors: SARS-CoV-2 Entry Blockers of COVID-19. Top. Curr. Chem. 2021, 379, 40. [Google Scholar] [CrossRef] [PubMed]
- Pant, S.; Singh, M.; Ravichandiran, V.; Murty, U.S.N.; Srivastava, H.K. Peptide-like and Small-Molecule Inhibitors against COVID-19. J. Biomol. Struct. Dyn. 2021, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Veeramachaneni, G.K.; Thunuguntla, V.B.S.C.; Bobbillapati, J.; Bondili, J.S. Structural and Simulation Analysis of Hotspot Residues Interactions of SARS-CoV 2 with Human ACE2 Receptor. J. Biomol. Struct. Dyn. 2021, 39, 4015–4025. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 Spike Receptor-Binding Domain Bound to the ACE2 Receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor Recognition by the Novel Coronavirus from Wuhan: An Analysis Based on Decade-Long Structural Studies of SARS Coronavirus. J. Virol. 2020, 94, 127–147. [Google Scholar] [CrossRef]
- Han, D.P.; Penn-Nicholson, A.; Cho, M.W. Identification of Critical Determinants on ACE2 for SARS-CoV Entry and Development of a Potent Entry Inhibitor. Virology 2006, 350, 15–25. [Google Scholar] [CrossRef]
- Jani, V.; Koulgi, S.; Uppuladinne, V.N.M.; Sonavane, U.; Joshi, R. Computational Drug Repurposing Studies on the ACE2-Spike (RBD) Interface of SARS-CoV-2. ChemRxiv 2020. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.S.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-Spectrum in Vitro Activity and in Vivo Efficacy of the Antiviral Protein Griffithsin against Emerging Viruses of the Family Coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef]
- Mille, J.K.; Whittaker, G.R. Host Cell Entry of Middle East Respiratory Syndrome Coronavirus after Two-Step, Furin-Mediated Activation of the Spike Protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef]
- Ho, T.Y.; Wu, S.L.; Chen, J.C.; Li, C.C.; Hsiang, C.Y. Emodin Blocks the SARS Coronavirus Spike Protein and Angiotensin-Converting Enzyme 2 Interaction. Antiviral Res. 2007, 74, 92. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Wang, K.; Yu, W.; Sun, B.; Schwarz, W. Emodin Inhibits Current through SARS-Associated Coronavirus 3a Protein. Antiviral Res. 2011, 90, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hofmann-Winkler, H.; Smith, J.C.; Krüger, N.; Arora, P.; Sørensen, L.K.; Søgaard, O.S.; Hasselstrøm, J.B.; Winkler, M.; Hempel, T.; et al. Camostat Mesylate Inhibits SARS-CoV-2 Activation by TMPRSS2-Related Proteases and Its Metabolite GBPA Exerts Antiviral Activity. EBioMedicine 2021, 65, 103255. [Google Scholar] [CrossRef]
- Muhseen, Z.T.; Hameed, A.R.; Al-Hasani, H.M.H.; Tahir ul Qamar, M.; Li, G. Promising Terpenes as SARS-CoV-2 Spike Receptor-Binding Domain (RBD) Attachment Inhibitors to the Human ACE2 Receptor: Integrated Computational Approach. J. Mol. Liq. 2020, 320, 114493. [Google Scholar] [CrossRef] [PubMed]
- Hakami, A.R. Targeting the RBD of Omicron Variant (B.1.1.529) with Medicinal Phytocompounds to Abrogate the Binding of Spike Glycoprotein with the HACE2 Using Computational Molecular Search and Simulation Approach. Biology 2022, 11, 258. [Google Scholar] [CrossRef]
- Dyall, J.; Coleman, C.M.; Hart, B.J.; Venkataraman, T.; Holbrook, M.R.; Kindrachuk, J.; Johnson, R.F.; Olinger, G.G.; Jahrling, P.B.; Laidlaw, M.; et al. Repurposing of Clinically Developed Drugs for Treatment of Middle East Respiratory Syndrome Coronavirus Infection. Antimicrob. Agents Chemother. 2014, 58, 4885–4893. [Google Scholar] [CrossRef]
- Zhao, Y.; Ren, J.; Harlos, K.; Jones, D.M.; Zeltina, A.; Bowden, T.A.; Padilla-Parra, S.; Fry, E.E.; Stuart, D.I. Toremifene Interacts with and Destabilizes the Ebola Virus Glycoprotein. Nature 2016, 535, 169–172. [Google Scholar] [CrossRef]
- Marques-Pereira, C.; Pires, M.N.; Gouveia, R.P.; Pereira, N.N.; Caniceiro, A.B.; Rosário-Ferreira, N.; Moreira, I.S. SARS-CoV-2 Membrane Protein: From Genomic Data to Structural New Insights. Int. J. Mol. Sci. 2022, 23, 2986. [Google Scholar] [CrossRef]
- Kumar, P.; Kumar, A.; Garg, N.; Giri, R. An Insight into SARS-CoV-2 Membrane Protein Interaction with Spike, Envelope, and Nucleocapsid Proteins. J. Biomol. Struct. Dyn. 2021, 1–10. [Google Scholar] [CrossRef]
- Gupta, M.K.; Vemula, S.; Donde, R.; Gouda, G.; Behera, L.; Vadde, R. In-Silico Approaches to Detect Inhibitors of the Human Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Ion Channel. J. Biomol. Struct. Dyn. 2021, 39, 2617–2627. [Google Scholar] [CrossRef]
- Liu, X.; Verma, A.; Garcia, G.; Ramage, H.; Lucas, A.; Myers, R.L.; Michaelson, J.J.; Coryell, W.; Kumar, A.; Charney, A.W.; et al. Targeting the Coronavirus Nucleocapsid Protein through GSK-3 Inhibition. Proc. Natl. Acad. Sci. USA 2021, 118, e2113401118. [Google Scholar] [CrossRef] [PubMed]
- Roh, C. A Facile Inhibitor Screening of SARS Coronavirus N Protein Using Nanoparticle-Based RNA Oligonucleotide. Int. J. Nanomed. 2012, 7, 2173. [Google Scholar] [CrossRef]
- Kwarteng, A.; Asiedu, E.; Sakyi, S.A.; Asiedu, S.O. Targeting the SARS-CoV2 Nucleocapsid Protein for Potential Therapeutics Using Immuno-Informatics and Structure-Based Drug Discovery Techniques. Biomed. Pharmacother. 2020, 132, 110914. [Google Scholar] [CrossRef]
- Lavigne, M.; Helynck, O.; Rigolet, P.; Boudria-Souilah, R.; Nowakowski, M.; Baron, B.; Brülé, S.; Hoos, S.; Raynal, B.; Guittat, L.; et al. SARS-CoV-2 Nsp3 Unique Domain SUD Interacts with Guanine Quadruplexes and G4-Ligands Inhibit This Interaction. Nucleic Acids Res 2021, 49, 7695–7712. [Google Scholar] [CrossRef]
- Lim, C.T.; Tan, K.W.; Wu, M.; Ulferts, R.; Armstrong, L.A.; Ozono, E.; Drury, L.S.; Milligan, J.C.; Zeisner, T.U.; Zeng, J.; et al. Identifying SARS-CoV-2 Antiviral Compounds by Screening for Small Molecule Inhibitors of Nsp3 Papain-like Protease. Biochem. J. 2021, 478, 2517. [Google Scholar] [CrossRef] [PubMed]
- Roe, M.K.; Junod, N.A.; Young, A.R.; Beachboard, D.C.; Stobart, C.C. Targeting Novel Structural and Functional Features of Coronavirus Protease Nsp5 (3CL pro, M pro) in the Age of COVID-19. J. Gen. Virol. 2021, 102, 001558. [Google Scholar] [CrossRef]
- Malik, Y.A. Properties of Coronavirus and SARS-CoV-2. Malays. J. Pathol. 2020, 42, 3–11. [Google Scholar]
- Lee, J.; Worrall, L.J.; Vuckovic, M.; Rosell, F.I.; Gentile, F.; Ton, A.T.; Caveney, N.A.; Ban, F.; Cherkasov, A.; Paetzel, M.; et al. Crystallographic Structure of Wild-Type SARS-CoV-2 Main Protease Acyl-Enzyme Intermediate with Physiological C-Terminal Autoprocessing Site. Nat. Commun. 2020, 11, 5877. [Google Scholar] [CrossRef]
- Milligan, J.C.; Zeisner, T.U.; Papageorgiou, G.; Joshi, D.; Soudy, C.; Ulferts, R.; Wu, M.; Lim, C.T.; Tan, K.W.; Weissmann, F.; et al. Identifying SARS-CoV-2 Antiviral Compounds by Screening for Small Molecule Inhibitors of Nsp5 Main Protease. Biochem. J. 2021, 478, 2499–2515. [Google Scholar] [CrossRef]
- Dai, W.; Zhang, B.; Jiang, X.M.; Su, H.; Li, J.; Zhao, Y.; Xie, X.; Jin, Z.; Peng, J.; Liu, F.; et al. Structure-Based Design of Antiviral Drug Candidates Targeting the SARS-CoV-2 Main Protease. Science 2020, 368, 1331–1335. [Google Scholar] [CrossRef] [PubMed]
- Naidu, S.A.G.; Tripathi, Y.B.; Shree, P.; Clemens, R.A.; Naidu, A.S. Phytonutrient Inhibitors of SARS-CoV-2/NSP5-Encoded Main Protease (Mpro) Autocleavage Enzyme Critical for COVID-19 Pathogenesis. J. Diet. Suppl. 2021. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.K.; Gadthula, S.; Chen, X.; Choo, H.; Olgen, S.; Barnard, D.L.; Sidwell, R.W. Antiviral Activity of Nucleoside Analogues against SARS-Coronavirus (SARS-CoV). Antivir. Chem. Chemother. 2006, 17, 285–289. [Google Scholar] [CrossRef]
- Gandhi, Y.; Mishra, S.K.; Rawat, H.; Grewal, J.; Kumar, R.; Shakya, S.K.; Jain, V.K.; Babu, G.; Singh, A.; Singh, R.; et al. Phytomedicines Explored under in Vitro and in Silico Studies against Coronavirus: An Opportunity to Develop Traditional Medicines. S. Afr. J. Bot. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Pompei, R.; Laconi, S.; Ingianni, A. Antiviral Properties of Glycyrrhizic Acid and Its Semisynthetic Derivatives. Mini Rev. Med. Chem. 2009, 9, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Hoever, G.; Baltina, L.; Michaelis, M.; Kondratenko, R.; Baltina, L.; Tolstikov, G.A.; Doerr, H.W.; Cinatl, J. Antiviral Activity of Glycyrrhizic Acid Derivatives against SARS-Coronavirus. J. Med. Chem. 2005, 48, 1256–1259. [Google Scholar] [CrossRef]
- Taylor, J.K.; Coleman, C.M.; Postel, S.; Sisk, J.M.; Bernbaum, J.G.; Venkataraman, T.; Sundberg, E.J.; Frieman, M.B. Severe Acute Respiratory Syndrome Coronavirus ORF7a Inhibits Bone Marrow Stromal Antigen 2 Virion Tethering through a Novel Mechanism of Glycosylation Interference. J. Virol. 2015, 89, 11820–11833. [Google Scholar] [CrossRef]
- Narayanan, K.; Huang, C.; Lokugamage, K.; Kamitani, W.; Ikegami, T.; Tseng, C.-T.K.; Makino, S. Severe Acute Respiratory Syndrome Coronavirus Nsp1 Suppresses Host Gene Expression, Including That of Type I Interferon, in Infected Cells. J. Virol. 2008, 82, 4471–4479. [Google Scholar] [CrossRef]
- Forni, D.; Cagliani, R.; Mozzi, A.; Pozzoli, U.; Al-Daghri, N.; Clerici, M.; Sironi, M. Extensive Positive Selection Drives the Evolution of Nonstructural Proteins in Lineage C Betacoronaviruses. J. Virol. 2016, 90, 3627–3639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
An overview of the SARS-CoV-2 infection pathway. During viral infection, immune cells are activated and release several cytokines as required for the biological system. A high virus titer is associated with a cytokine storm, and such dysregulation in the body of the patient may lead to multi-organ dysfunction syndrome. GIT—gastrointestinal tract.
Figure 1.
An overview of the SARS-CoV-2 infection pathway. During viral infection, immune cells are activated and release several cytokines as required for the biological system. A high virus titer is associated with a cytokine storm, and such dysregulation in the body of the patient may lead to multi-organ dysfunction syndrome. GIT—gastrointestinal tract.

Figure 2.
SARS-CoV-2 causes infection in the lungs mainly via DAMPs and PAMPs produced as a result of the action of nearly all Toll-like receptors (TLRs). Only TLRs involved in virus sensing and/or signaling are displayed here.
Figure 2.
SARS-CoV-2 causes infection in the lungs mainly via DAMPs and PAMPs produced as a result of the action of nearly all Toll-like receptors (TLRs). Only TLRs involved in virus sensing and/or signaling are displayed here.

Table 3.
Toll-like receptors (TLRs) and their physiological significance.
| TLRs | Ligand Recognition | Form | Localization | Adaptor Molecules | Negative Adaptors | Response | Reference |
|---|---|---|---|---|---|---|---|
| TLR1 | Triacyl lipopeptides, soluble factors | Heterodimer | Cell surface | MyD88, Mal | - | NF-κB activation and proinflammatory cytokines | [99,100] |
| TLR2 | Hsp70, lipopeptide, HCV, Nonstructural protein 3 | Heterodimer | Cell surface | MyD88, Mal | - | NF-κB activation and proinflammatory cytokines | [101,102] |
| TLR3 | dsRNA | Homodimers | Endosomal membrane | TRIF | SARM negatively regulates TRIF | IRF activation, production of type 1 IFNs and proinflammatory cytokines | [103,104] |
| TLR4 | Lipopolysaccharide, Taxol, S protein of SARS-CoV-2 | Homodimers | Cell surface | MyD88, Mal, TRIF, TRAM | SARM negatively regulates TRIF and TRAM to consequently reduce inflammation | Activation of NF-κB, pro-inflammatory cytokines, and IFN-inducible genes | [105,106] |
| TLR5 | Flagellin | Homodimers | Cell surface | MyD88 | - | Activation of NF-κB and proinflammatory cytokines | [107,108] |
| TLR6 | Diacyl lipopeptides, lipoteichoic acid, fungal zymosan | Heterodimer | Cell surface | MyD88, Mal/TIRAP | - | Activation of NF-κB and proinflammatory cytokines | [109,110] |
| TLR7 | SARS-CoV-2 ssRNA, imadozoquinoline | Homodimers | Endosomal membrane | MyD88 | - | IRF activation, production of Type 1 IFNs and proinflammatory cytokines | [111,112] |
| TLR8 | SARS-CoV-2 ssRNA | Endosomal membrane | MyD88 | - | IRF activation, production of type 1 IFNs and proinflammatory cytokines | [113,114] | |
| TLR9 | Unmethylated CPG-containing ssDNA, hemozoin from the malaria parasite | Homodimers | Endosomal membrane | MyD88 | - | IRF activation, production of type 1 IFNs and proinflammatory cytokines | [115,116] |
Table 4.
Reported antiviral agonists employing Toll-like receptors (TLRs).
| Drugs | TLRs | Viruses | Significance | References |
|---|---|---|---|---|
| Pam2CSK4 | TLR2 | Parainfluenza | Reduced virus replication | [141] |
| INNA-051 | TLR2 | SARS-CoV-2 | Reduces viral RNA load | [142] |
| PIKA | TLR3 | Influenza A | Reduces virus load | [143] |
| Poly ICLC | TLR3 | HIV | Release of IFN-α/β/γ | [144] |
| NA6 | TLR4 | Norovirus | Induction of IFN-β | [145] |
| MPL | TLR4 | VZV | Stimulate cytokines | [146] |
| Flagellin | TLR5 | Influenza A | Reduces virus replication | [147] |
| CBLB502 | TLR5 | ConA | Activation of NF-κB | [148] |
| Pam2CSK4 | TLR6 | Parainfluenza | Reduces virus replication | [141] |
| INNA-051 | TLR6 | SARS-CoV-2 | Reduces viral RNA load | [142] |
| GS-9620 | TLR7 | HIV | Reactivates latency | [112] |
| Vesatolimod | TLR7 | HIV | Modest delay in viral rebound | [149] |
| R848 | TLR7/8 | Zika | Activation of NF-κB | [150] |
| GS-9688 | TLR8 | HBV | Activation of dendritic and natural killer cells | [151] |
| ODN2395 | TLR9 | Parainfluenza | Reduces viral replication | [141] |
CBLB502—Entolimod; ConA—Concanavalin A; GS-9688—Selgantolimod; R848—Resiquimod; NA6—neoagarohexaose; VZV—Varicella-Zoster virus.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Manan, A.; Pirzada, R.H.; Haseeb, M.; Choi, S. Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach. Int. J. Mol. Sci. 2022, 23, 10716. https://doi.org/10.3390/ijms231810716
AMA Style
Manan A, Pirzada RH, Haseeb M, Choi S. Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach. International Journal of Molecular Sciences. 2022; 23(18):10716. https://doi.org/10.3390/ijms231810716
Chicago/Turabian StyleManan, Abdul, Rameez Hassan Pirzada, Muhammad Haseeb, and Sangdun Choi. 2022. "Toll-like Receptor Mediation in SARS-CoV-2: A Therapeutic Approach" International Journal of Molecular Sciences 23, no. 18: 10716. https://doi.org/10.3390/ijms231810716
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.