
Impact of COVID-19 Lockdown on the Nasopharyngeal Microbiota of Children and Adults Self-Confined at Home

 , , , ,
, , , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Study Population
2.2. Inclusion and Exclusion Criteria
2.3. Microbiological Methods
2.4. DNA Extraction and 16S rRNA Sequencing
2.5. Bioinformatics Analysis
2.6. Statistical Analysis
3. Results
3.1. Epidemiological and Clinical Characteristics of Participants
3.2. Nasopharyngeal Microbiota of Children and Adults Dominated by Corynebacterium with Limited Relative Abundance of Common Pathobionts
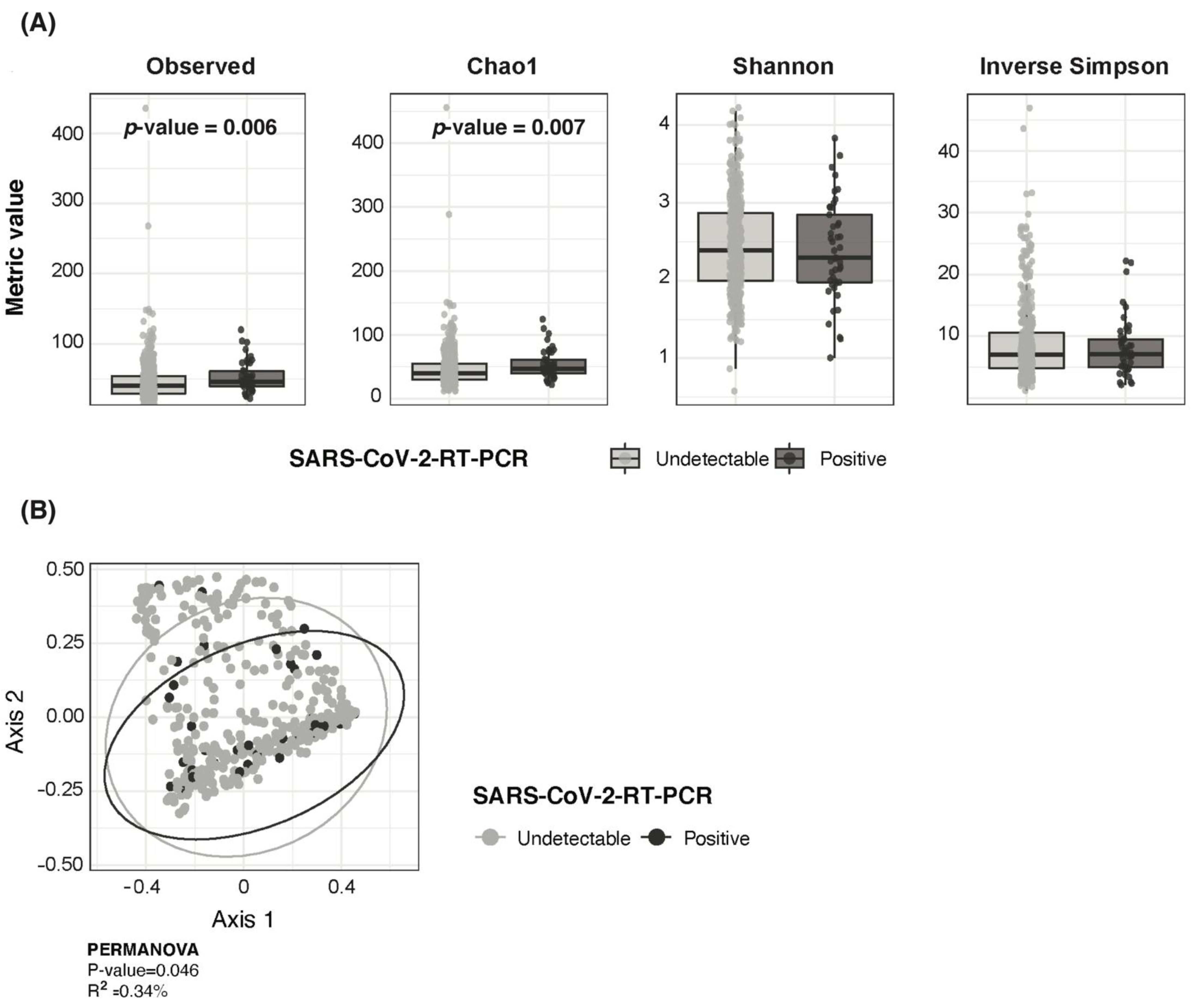
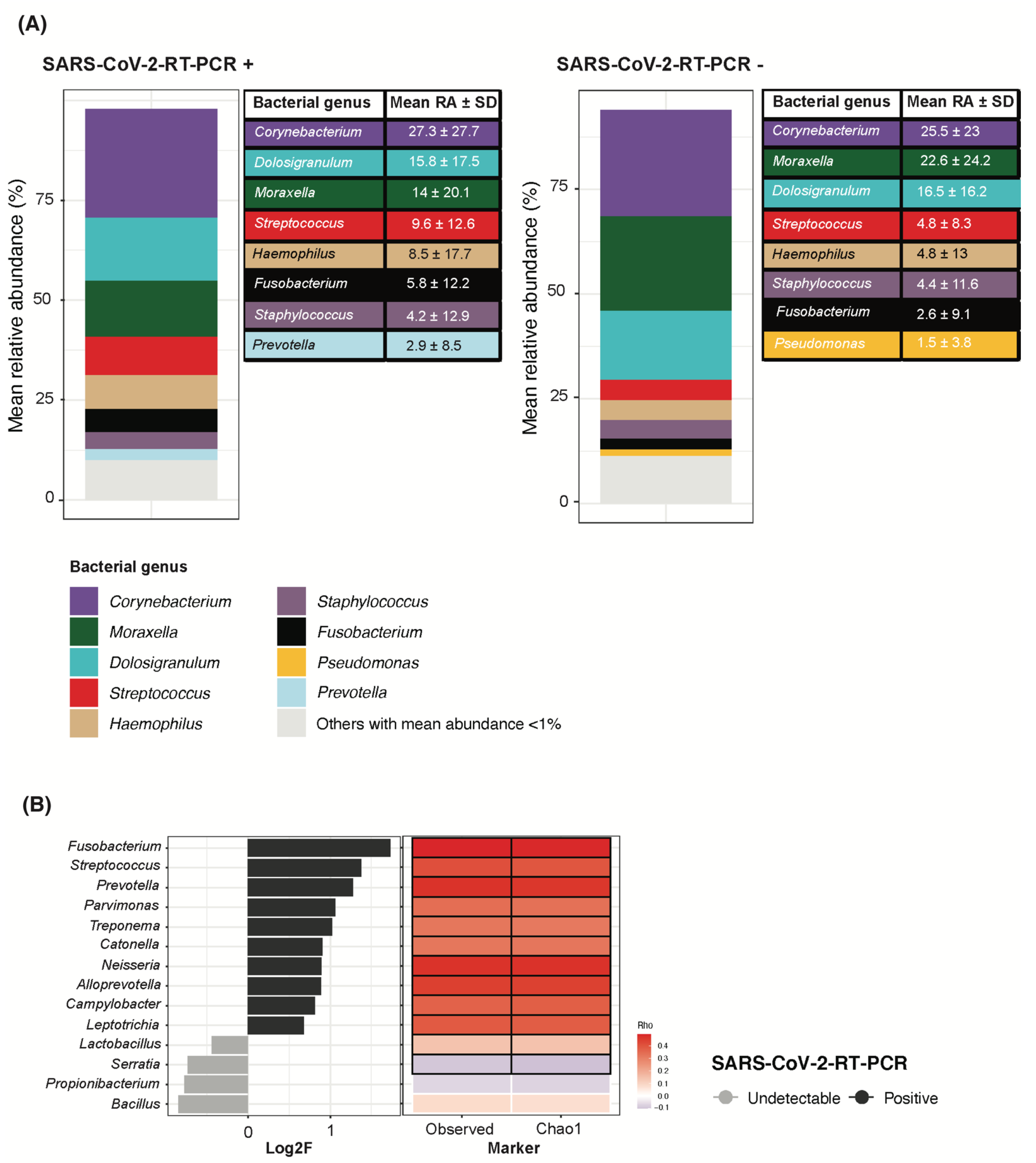
3.3. SARS-CoV-2 RNA Detection in Children Associated with Higher Bacterial Richness and Higher Fusobacterium, Streptococcus and Prevotella Abundance
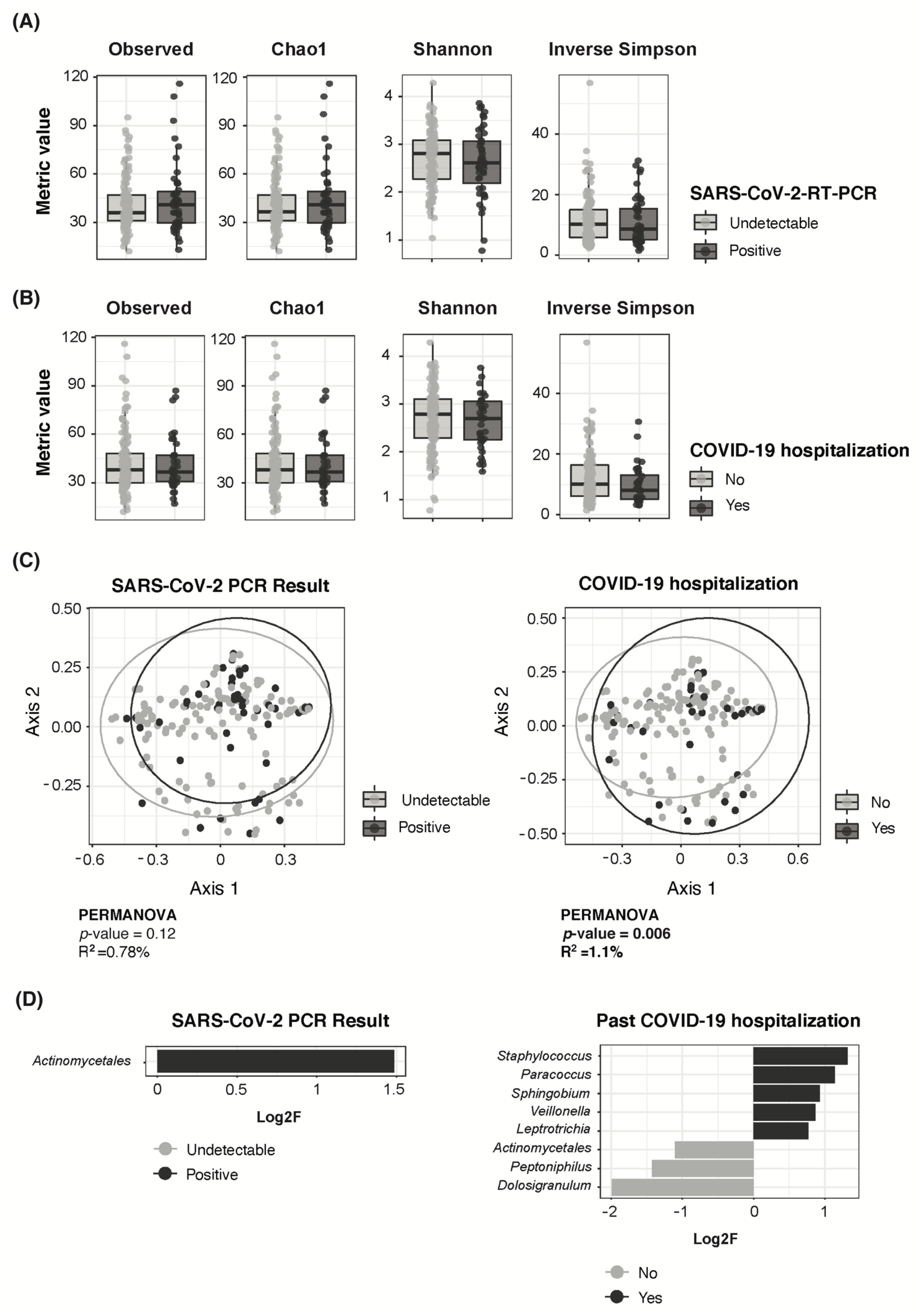
3.4. Adult COVID-19 Severity Associated to Higher Staphylococcus and Lower Dolosigranulum Abundance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Other Kids Corona Study Group Members
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Rosenwald, M.S. History’s Deadliest Pandemics: Plague, Smallpox, Flu, Covid-19. Washington Post. Available online: https://www.washingtonpost.com/graphics/2020/local/retropolis/coronavirus-deadliest-pandemics/ (accessed on 14 June 2022).
- Government Decrees State of Emergency to Stop Spread of Coronavirus COVID-19 [Government/Council of Ministers]. La Moncloa. Available online: https://www.lamoncloa.gob.es/lang/en/gobierno/councilministers/Paginas/2020/20200314council-extr.aspx (accessed on 14 June 2022).
- Ahmed, F.; Zviedrite, N.; Uzicanin, A. Effectiveness of workplace social distancing measures in reducing influenza transmission: A systematic review. BMC Public Health 2018, 18, 518. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, R.C.; Chughta, A.A. Facemasks for the prevention of infection in healthcare and community settings. BMJ 2015, 350, h694. [Google Scholar] [CrossRef] [PubMed]
- Brueggemann, A.B.; Jansen van Rensburg, M.J.; Shaw, D.; McCarthy, N.D.; Jolley, K.A.; Maiden, M.C.; van der Linden, M.P.; Amin-Chowdhury, Z.; Bennett, D.E.; Borrow, R.; et al. Changes in the incidence of invasive disease due to Streptococcus pneumoniae, Haemophilus influenzae, and Neisseria meningitidis during the COVID-19 pandemic in 26 countries and territories in the Invasive Respiratory Infection Surveillance Initiative: A prospective analysis of surveillance data. Lancet. Digit. Health 2021, 3, e360–e370. [Google Scholar] [PubMed]
- Haak, B.W.; Brands, X.; Davids, M.; Peters-Sengers, H.; Kullberg, R.F.J.; van Houdt, R.; Hugenholtz, F.; Faber, D.R.; Zaaijer, H.L.; Scicluna, B.P.; et al. Bacterial and viral respiratory tract microbiota and host characteristics in adults with lower respiratory tract infections: A case-control study. Clin. Infect. Dis. 2021, 74, 776–784. [Google Scholar] [CrossRef]
- Lanaspa, M.; Bassat, Q.; Medeiros, M.M.; Muñoz-Almagro, C. Respiratory microbiota and lower respiratory tract disease. Expert Rev. Anti. Infect. Ther. 2017, 15, 703–711. [Google Scholar] [CrossRef]
- de Steenhuijsen Piters, W.A.A.; Sanders, E.A.M.; Bogaert, D. The role of the local microbial ecosystem in respiratory health and disease. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140294. [Google Scholar] [CrossRef] [Green Version]
- Henares, D.; Brotons, P.; de Sevilla, M.F.; Fernandez-Lopez, A.; Hernandez-Bou, S.; Perez-Argüello, A.; Mira, A.; Muñoz-Almagro, C.; Cabrera-Rubio, R. Differential nasopharyngeal microbiota composition in children according to respiratory health status. Microb. Genom. 2021, 7, 000661. [Google Scholar] [CrossRef]
- Henares, D.; Rocafort, M.; Brotons, P.; de Sevilla, M.F.; Mira, A.; Launes, C.; Cabrera-Rubio, R.; Muñoz-Almagro, C. Rapid Increase of Oral Bacteria in Nasopharyngeal Microbiota After Antibiotic Treatment in Children With Invasive Pneumococcal Disease. Front. Cell. Infect. Microbiol. 2021, 11, 929. [Google Scholar] [CrossRef]
- Man, W.H.; De Steenhuijsen Piters, W.A.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef]
- Brotons, P.; Launes, C.; Buetas, E.; Fumado, V.; Henares, D.; de Sevilla, M.F.; Redin, A.; Fuente-Soro, L.; Cuadras, D.; Mele, M.; et al. Susceptibility to Severe Acute Respiratory Syndrome Coronavirus 2 Infection Among Children and Adults: A Seroprevalence Study of Family Households in the Barcelona Metropolitan Region, Spain. Clin. Infect. Dis. 2021, 72, e970. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014, 42, D633–D642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. R Package Version 2.0–8 2013, 2, 254. [Google Scholar] [CrossRef]
- Lin, H.; Das Peddada, S. Analysis of compositions of microbiomes with bias correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camelo-Castillo, A.; Henares, D.; Brotons, P.; Galiana, A.; Rodríguez, J.C.; Mira, A.; Muñoz-Almagro, C. Nasopharyngeal Microbiota in Children With Invasive Pneumococcal Disease: Identification of Bacteria With Potential Disease-Promoting and Protective Effects. Front. Microbiol. 2019, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.A.T.M.; de Steenhuijsen Piters, W.A.A.; van Houten, M.A.; Chu, M.L.J.N.; Biesbroek, G.; Kool, J.; Pernet, P.; de Groot, P.K.C.M.; Eijkemans, M.J.C.; Keijser, B.J.F.; et al. Maturation of the Infant Respiratory Microbiota, Environmental Drivers, and Health Consequences. A Prospective Cohort Study. Am. J. Respir. Crit. Care Med. 2017, 196, 1582–1590. [Google Scholar] [CrossRef]
- Bogaert, D.; Keijser, B.; Huse, S.; Rossen, J.; Veenhoven, R.; van Gils, E.; Bruin, J.; Montijn, R.; Bonten, M.; Sanders, E. Variability and Diversity of Nasopharyngeal Microbiota in Children: A Metagenomic Analysis. PLoS ONE 2011, 6, e17035. [Google Scholar] [CrossRef] [Green Version]
- Biesbroek, G.; Bosch, A.A.T.M.; Wang, X.; Keijser, B.J.F.; Veenhoven, R.H.; Sanders, E.A.M.; Bogaert, D. The impact of breastfeeding on nasopharyngeal microbial communities in infants. Am. J. Respir. Crit. Care Med. 2014, 190, 298–308. [Google Scholar] [CrossRef]
- Bosch, A.A.T.M.; Levin, E.; van Houten, M.A.; Hasrat, R.; Kalkman, G.; Biesbroek, G.; de Steenhuijsen Piters, W.A.A.; de Groot, P.K.C.M.; Pernet, P.; Keijser, B.J.F.; et al. Development of Upper Respiratory Tract Microbiota in Infancy is Affected by Mode of Delivery. EBioMedicine 2016, 9, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Kaul, D.; Rathnasinghe, R.; Ferres, M.; Tan, G.S.; Barrera, A.; Pickett, B.E.; Methe, B.A.; Das, S.R.; Budnik, I.; Halpin, R.A.; et al. Microbiome disturbance and resilience dynamics of the upper respiratory tract during influenza A virus infection. Nat. Commun. 2020, 11, 2537. [Google Scholar] [CrossRef]
- Dubourg, G.; Edouard, S.; Raoult, D. Relationship between nasopharyngeal microbiota and patient’s susceptibility to viral infection. Expert Rev. Anti. Infect. Ther. 2019, 17, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Buehrle, D.J.; Wagener, M.M.; Nguyen, M.H.; Clancy, C.J. Trends in Outpatient Antibiotic Prescriptions in the United States During the COVID-19 Pandemic in 2020. JAMA Netw. Open 2021, 4, e2126114. [Google Scholar] [CrossRef]
- Peñalva, G.; Benavente, R.S.; Pérez-Moreno, M.A.; Pérez-Pacheco, M.D.; Pérez-Milena, A.; Murcia, J.; Cisneros, J.M. Effect of the coronavirus disease 2019 pandemic on antibiotic use in primary care. Clin. Microbiol. Infect. 2021, 27, 1058. [Google Scholar] [CrossRef]
- Bomar, L.; Brugger, S.D.; Yost, B.H.; Davies, S.S.; Lemon, K.P. Corynebacterium accolens Releases Antipneumococcal Free Fatty Acids from Human Nostril and Skin Surface Triacylglycerols. MBio 2016, 7, e01725-15. [Google Scholar] [CrossRef] [Green Version]
- Brugger, S.D.; Eslami, S.M.; Pettigrew, M.M.; Escapa, I.F.; Henke, M.T.; Kong, Y.; Lemon, K.P.; D’orazio, S.E.F. Dolosigranulum pigrum Cooperation and Competition in Human Nasal Microbiota. mSphere 2020, 5, e00852-20. [Google Scholar] [CrossRef]
- Cremers, A.J.; Zomer, A.L.; Gritzfeld, J.F.; Ferwerda, G.; van Hijum, S.A.; Ferreira, D.M.; Shak, J.R.; Klugman, K.P.; Boekhorst, J.; Timmerman, H.M.; et al. The adult nasopharyngeal microbiome as a determinant of pneumococcal acquisition. Microbiome 2014, 2, 44. [Google Scholar] [CrossRef] [Green Version]
- De Boeck, I.; Wittouck, S.; Wuyts, S.; Oerlemans, E.F.M.; van den Broek, M.F.L.; Vandenheuvel, D.; Vanderveken, O.; Lebeer, S. Comparing the healthy nose and nasopharynx microbiota reveals continuity as well as niche-specificity. Front. Microbiol. 2017, 8, 2372. [Google Scholar] [CrossRef] [Green Version]
- Burchill, E.; Lymberopoulos, E.; Menozzi, E.; Budhdeo, S.; McIlroy, J.R.; Macnaughtan, J.; Sharma, N. The Unique Impact of COVID-19 on Human Gut Microbiome Research. Front. Med. 2021, 8, 652464. [Google Scholar] [CrossRef]
- Xu, R.; Liu, P.; Zhang, T.; Wu, Q.; Zeng, M.; Ma, Y.; Jin, X.; Xu, J.; Zhang, Z.; Zhang, C. Progressive deterioration of the upper respiratory tract and the gut microbiomes in children during the early infection stages of COVID-19. J. Genet. Genom. 2021, 48, 803–814. [Google Scholar] [CrossRef]
- Nardelli, C.; Gentile, I.; Setaro, M.; Di Domenico, C.; Pinchera, B.; Buonomo, A.R.; Zappulo, E.; Scotto, R.; Scaglione, G.L.; Castaldo, G.; et al. Nasopharyngeal Microbiome Signature in COVID-19 Positive Patients: Can We Definitively Get a Role to Fusobacterium periodonticum? Front. Cell. Infect. Microbiol. 2021, 11, 18. [Google Scholar] [CrossRef]
- Wolff, L.; Martiny, D.; Deyi, V.Y.M.; Maillart, E.; Clevenbergh, P.; Dauby, N. COVID-19–Associated Fusobacterium nucleatum Bacteremia, Belgium. Emerg. Infect. Dis. 2021, 27, 975. [Google Scholar] [CrossRef]
- Aykac, K.; Ozsurekci, Y.; Cura Yayla, B.C.; Evren, K.; Lacinel Gurlevik, S.; Oygar, P.D.; Yucel, M.; Karakoc, A.E.; Alp, A.; Cengiz, A.B.; et al. Pneumococcal carriage in children with COVID-19. Hum. Vaccines Immunother. 2021, 17, 1628–1634. [Google Scholar] [CrossRef]
- Engen, P.A.; Naqib, A.; Jennings, C.; Green, S.J.; Landay, A.; Keshavarzian, A.; Voigt, R.M. Nasopharyngeal Microbiota in SARS-CoV-2 Positive and Negative Patients. Biol. Proced. Online 2021, 23, 10. [Google Scholar] [CrossRef]
- Merenstein, C.; Liang, G.; Whiteside, S.A.; Cobián-Güemes, A.G.; Merlino, M.S.; Taylor, L.J.; Glascock, A.; Bittinger, K.; Tanes, C.; Graham-Wooten, J.; et al. Signatures of COVID-19 severity and immune response in the respiratory tract microbiome. medRxiv, 2021; Preprint. [Google Scholar] [CrossRef]
- Mostafa, H.H.; Fissel, J.A.; Fanelli, B.; Bergman, Y.; Gniazdowski, V.; Dadlani, M.; Carroll, K.C.; Colwell, R.R.; Simner, P.J. Metagenomic Next-Generation Sequencing of Nasopharyngeal Specimens Collected from Confirmed and Suspect COVID-19 Patients. MBio 2020, 11, e01969-20. [Google Scholar] [CrossRef]
- De Maio, F.; Posteraro, B.; Ponziani, F.R.; Cattani, P.; Gasbarrini, A.; Sanguinetti, M. Nasopharyngeal Microbiota Profiling of SARS-CoV-2 Infected Patients. Biol. Proced. Online 2020, 22, 18. [Google Scholar] [CrossRef]
- Pettigrew, M.M.; Laufer, A.S.; Gent, J.F.; Kong, Y.; Fennie, K.P.; Metlay, J.P. Upper Respiratory Tract Microbial Communities, Acute Otitis Media Pathogens, and Antibiotic Use in Healthy and Sick Children. Appl. Environ. Microbiol. 2012, 78, 6262. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Infants (n = 470) | SARS-CoV-2 RNA Detection | p-Value Pos vs. Neg | |||
|---|---|---|---|---|---|
| Positive (n = 45) | Negative (n = 425) | ||||
| Gender, female | 226 (48.1%) | 22 (48.9%) | 204 (48%) | 1 a | |
| Median age, years (IQR) | 4.4 (2.6, 7.5) | 4.7 (2.4, 8.5) | 4.4 (2.7, 7.4) | 0.619 b | |
| Days since adult’s infection (IQR) | 52.5 (42, 61) | 49 (36, 59) | 53 (43, 61) | 0.181 b | |
| Median body temperature, °C (IQR) | 36 (35.7, 36.3) | 36 (35.6, 36.2) | 36 (35.7, 36.3) | 0.371 b | |
| Active respiratory symptoms | 21 (n = 468) (4.5%) | 3 (n = 41) (6.8%) | 18 (n = 424) (4.3%) | 0.688 a | |
| Antibiotic use (last 3 months) | 56 (n = 215) (26%) | 7 (n = 24) (29.2%) | 49 (n = 191) (25.7%) | 0.923 a | |
| Probiotic use (last 3 months) | 17 (n = 7.9%) | 1 (n = 24) (4.2%) | 16 (n = 191) (8.4%) | 0.750 a | |
| Other respiratory viruses | Overall | 115 (24.5%) | 20 (44.4%) | 95 (22.4%) | 0.002 a |
| Rhinovirus/Enterovirus | 89 (18.9%) | 18 (40%) | 71 (16.7%) | <0.001 a | |
| Adenovirus | 13 (2.8%) | 1 (2.2%) | 12 (2.8%) | 1 a | |
| Bocavirus | 29 (6.2%) | 6 (13.3%) | 23 (5.4%) | 0.076 a | |
| Coronavirus | 1 (0.21%) | 0 (0%) | 1 (0.24%) | 1 c | |
| Metapneumovirus | 0 (0%) | 0 (0%) | 0 (0%) | - | |
| VRS (type A and B) | 1 (0.21%) | 0 (0%) | 1 (0.24%) * | 1 c | |
| Influenza virus (A and B) | 1 (0.21%) | 0 (0%) | 1 (0.24%) ** | 1 c | |
| Parainfluenza virus 1 | 6 (1.28%) | 3 (6.7%) | 3 (0.70%) | 0.013 c | |
| Parainfluenza virus 2 | 4 (0.85%) | 1 (2.2%) | 3 (0.70%) | 0.332 c | |
| Parainfluenza virus 3 | 1 (0.21%) | 0 (0%) | 1 (0.24%) | 1 c | |
| Parainfluenza virus 4 | 0 | 0 (0%) | 0 (0%) (n = 424) | - | |
| All Adults (n = 173) | SARS-CoV-2 RNA Detection | p-Value Pos vs. Neg | |||
|---|---|---|---|---|---|
| Positive (n = 47) | Negative (n = 126) | ||||
| Gender, female | 63 (36.4%) | 30 (63.8%) | 80 (63.5%) | 1 a | |
| Median age, years (IQR) | 39.9 (35.9, 44.4) | 40 (36.2, 45.8) | 39.9 (35.9, 43.9) | 0.639 b | |
| Days since first infection (IQR) | 53 (44, 61) | 50 (43.5, 56.5) | 53.5 (46, 61) | 0.150 b | |
| Median body temperature, °C (IQR) | 36 (35.6, 36.2) | 36.1 (35.6, 36.3) | 35.9 (35.5, 36.2) | 0.298 b | |
| Active respiratory symptoms | 20 (n = 170) (11.8%) | 10 (n = 45) (22.2%) | 10 (n = 125) (8%) | 0.023 a | |
| Antibiotic use (last 3 months) | 54 (n = 138) (39.1%) | 13 (n = 41) (31.7%) | 41 (n = 97) (42.3%) | 0.332 b | |
| Probiotic use (last 3 months) | 15 (n = 138) (10.9%) | 4 (n = 40) (10%) | 11 (n = 98) (11.2%) | 1 b | |
| Other respiratory viruses | Overall | 10 (n = 172) (5.8%) | 7 (14.9%) | 3 (n = 125) (2.4%) | 0.006 a |
| Rhinovirus/Enterovirus | 9 (n = 172) (5.2%) | 6 (12.8%) | 3 (n = 125) (2.4%) | 0.019 a | |
| Adenovirus | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Bocavirus | 2 (n = 172) (1.2%) | 1 (2.1%) | 1 (n = 125) (0.8%) | 0.473 c | |
| Coronavirus | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Metapneumovirus | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| VRS (type A and B) | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Influenza virus (A and B) | 1 (n = 172) (0.58%) * | 1 (2.1%) | 0 (n = 125) (0%) | - | |
| Parainfluenza virus 1 | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Parainfluenza virus 2 | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Parainfluenza virus 3 | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| Parainfluenza virus 4 | 0 (n = 172) (0%) | 0 (0%) | 0 (n = 125) (0%) | - | |
| COVID-19 inpatient history | p-value Yes vs. No | ||||
| Yes (n = 36) | No (n = 137) | ||||
| Gender, female | 11 (30.6%) | 99 (72.3%) | <0.001 a | ||
| Median age, years (IQR) | 44.2 (36.9, 48.4) | 39 (35.6, 42.9) | <0.001 b | ||
| Days since first infection (IQR) | 51 (41.8, 56.8) | 53 (46, 61) | 0.255 b | ||
| Median body temperature, °C (IQR) | 36 (35.6, 36.3) | 36 (35.6, 36.2) | 0.804 b | ||
| Active respiratory symptoms | 9 (n = 35) (25.7%) | 11 (n = 135) (8.1%) | 0.551 a | ||
| Antibiotic use (last 3 months) | 29 (n = 34) (85.3%) | 25 (n = 104) (24%) | <0.001 a | ||
| Probiotic use (last 3 months) | 6 (n = 33) (18.2%) | 9 (n = 105) (8.6%) | 0.22 a | ||
| Actual SARS-CoV-2 PCR (positive) | 8 (22.2%) | 39 (28.5%) | 0.589 a | ||
| Other respiratory viruses | Overall | 1 (n = 35) (2.9%) | 9 (6.60%) | 0.665 a | |
| Rhinovirus/Enterovirus | 1 (n = 35) (2.9%) | 8 (5.84%) | 0.778 a | ||
| Adenovirus | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Bocavirus | 0 (n = 35) (0%) | 2 (1.46%) | 1 c | ||
| Coronavirus | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Metapneumovirus | 0 (n = 35) (0%) | 0 (0%) | - | ||
| VRS (type A and B) | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Influenza virus (A and B) | 0 (n = 35) (0%) | 1 (0.73%) * | 1 c | ||
| Parainfluenza virus 1 | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Parainfluenza virus 2 | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Parainfluenza virus 3 | 0 (n = 35) (0%) | 0 (0%) | - | ||
| Parainfluenza virus 4 | 0 (n = 35) (0%) | 0 (0%) | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocafort, M.; Henares, D.; Brotons, P.; Launes, C.; Fernandez de Sevilla, M.; Fumado, V.; Barrabeig, I.; Arias, S.; Redin, A.; Ponomarenko, J.; et al. Impact of COVID-19 Lockdown on the Nasopharyngeal Microbiota of Children and Adults Self-Confined at Home. Viruses 2022, 14, 1521. https://doi.org/10.3390/v14071521
Rocafort M, Henares D, Brotons P, Launes C, Fernandez de Sevilla M, Fumado V, Barrabeig I, Arias S, Redin A, Ponomarenko J, et al. Impact of COVID-19 Lockdown on the Nasopharyngeal Microbiota of Children and Adults Self-Confined at Home. Viruses. 2022; 14(7):1521. https://doi.org/10.3390/v14071521
Chicago/Turabian StyleRocafort, Muntsa, Desiree Henares, Pedro Brotons, Cristian Launes, Mariona Fernandez de Sevilla, Victoria Fumado, Irene Barrabeig, Sara Arias, Alba Redin, Julia Ponomarenko, and et al. 2022. "Impact of COVID-19 Lockdown on the Nasopharyngeal Microbiota of Children and Adults Self-Confined at Home" Viruses 14, no. 7: 1521. https://doi.org/10.3390/v14071521