
Healthcare-Associated SARS-CoV-2 Reinfection after 3 Months with a Phylogenetically Distinct Omicron Variant: A Case Report

 , ,
, ,
Abstract
:1. Introduction
2. Results
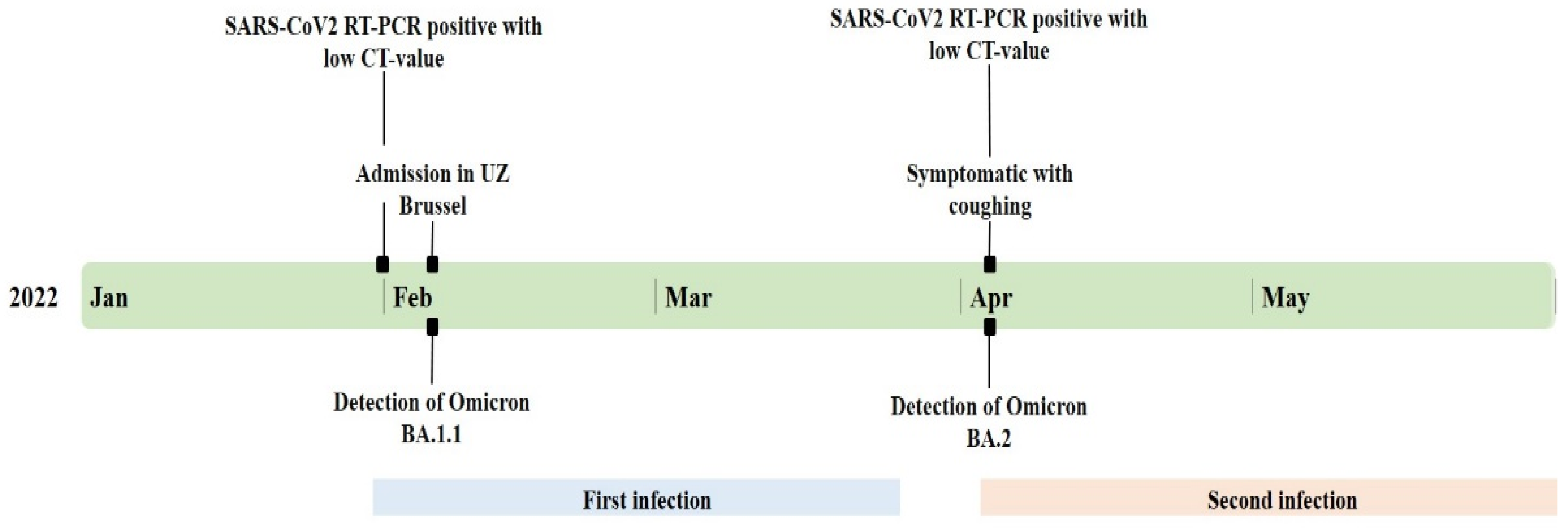
2.1. Case Description
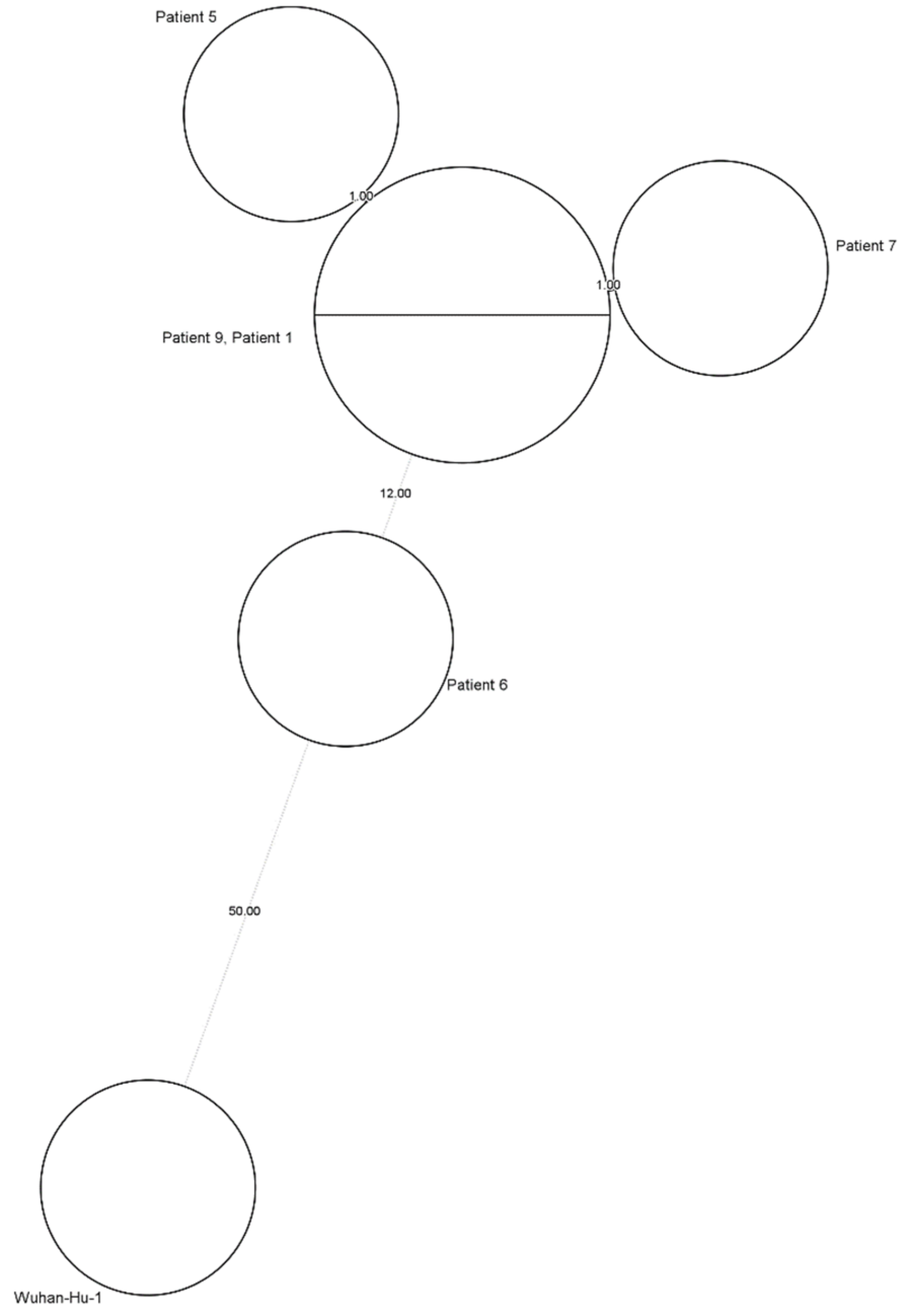
2.2. Healthcare-Associated Outbreak Analysis
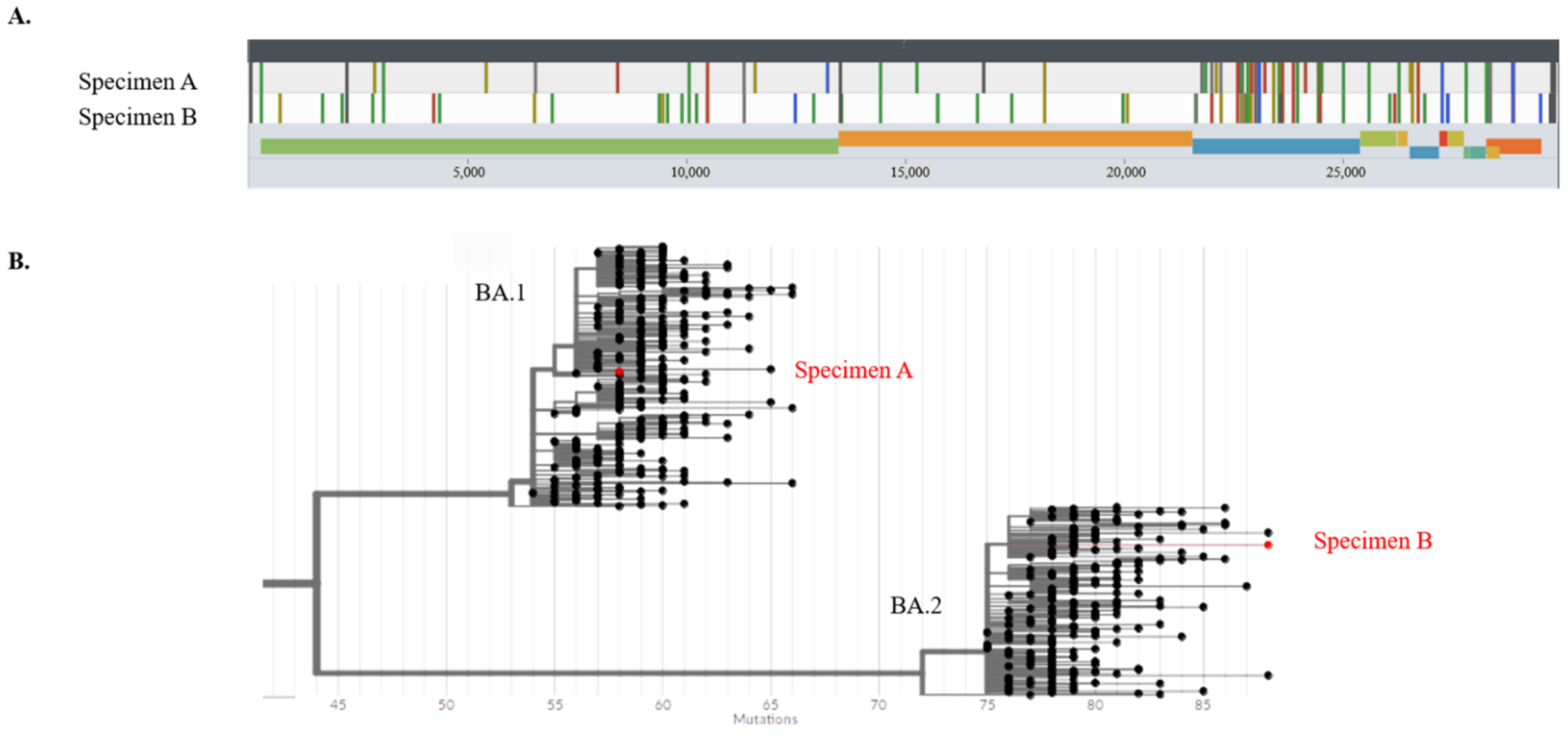
2.3. SARS-CoV-2 Strain Analysis
2.4. SARS-CoV-2 Serology
3. Materials and Methods
3.1. Healthcare-Associated Infections and Outbreak Analysis
3.2. SARS-CoV-2 Reverse-Transcriptase Semi-Quantitative Polymerase Chain Reaction (RT-qPCR)
3.3. SARS-CoV-2 Whole Genome Sequencing (WGS)
3.4. SARS-CoV-2 Serology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organisation: Coronavirus Disease (COVID-19) [Internet]. Available online: https://www.who.int/health-topics/coronavirus#tab=tab_1 (accessed on 22 June 2022).
- Murchu, E.O.; Byrne, P.; Walsh, K.A.; Carty, P.G.; Connolly, M.; De Gascun, C.; Jordan, K.; Keoghan, M.; O’Brien, K.K.; O’Neill, M.; et al. Immune response following infection with SARS-CoV-2 and other coronaviruses: A rapid review. Rev. Med. Virol. 2021, 31, e2162. [Google Scholar] [CrossRef]
- World Health Organization. COVID-19 Natural Immunity; WHO: Geneva, Switzerland, 2021; pp. 11–14. [Google Scholar]
- Hall, V.J.; Foulkes, S.; Charlett, A.; Atti, A.; Monk, E.J.M.; Simmons, R.; Wellington, E.; Cole, M.J.; Saei, A.; Oguti, B.; et al. SARS-CoV-2 infection rates of antibody-positive compared with antibody-negative health-care workers in England: A large, multicentre, prospective cohort study (SIREN). Lancet 2021, 397, 1459–1469. [Google Scholar] [CrossRef]
- Michlmayr, D.; Hansen, C.H.; Gubbels, S.M.; Valentiner-Branth, P.; Bager, P.; Obel, N.; Drewes, B.; Møller, C.H.; Møller, F.T.; Legarth, R.; et al. Observed protection against SARS-CoV-2 reinfection following a primary infection: A Danish cohort study among unvaccinated using two years of nationwide PCR-test data. Lancet Reg. Health Eur. 2022, 20, 100452. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.W.; Hung, I.F.N.; Ip, J.D.; Chu, A.W.H.; Chan, W.M.; Tam, A.R.; Fong, C.H.; Yuan, S.; Tsoi, H.W.; Ng, A.C.; et al. Coronavirus Disease 2019 (COVID-19) Re-infection by a Phylogenetically Distinct Severe Acute Respiratory Syndrome Coronavirus 2 Strain Confirmed by Whole Genome Sequencing. Clin. Infect. Dis. 2021, 73, E2946–E2951. [Google Scholar] [CrossRef] [PubMed]
- Schwendimann, R.; Blatter, C.; Dhaini, S.; Simon, M.; Ausserhofer, D. The occurrence, types, consequences and preventability of in-hospital adverse events—A scoping review. BMC Health Serv. Res. 2018, 18, 521. [Google Scholar] [CrossRef]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.J.; Edwards, J.R.; Sievert, D.M. Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control. Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef] [Green Version]
- Graman, P.S.; Hall, C.B. Epidemiology and control of nosocomial viral infections. Infect. Dis. Clin. N. Am. 1989, 3, 815–842. [Google Scholar] [CrossRef]
- Hong, H.L.; Hong, S.B.; Ko, G.B.; Huh, J.W.; Sung, H.; Do, K.H.; Kim, S.-H.; Lee, S.-O.; Kim, M.-N.; Jeong, J.-Y.; et al. Viral infection is not uncommon in adult patients with severe hospital-acquired pneumonia. PLoS ONE 2014, 9, e95865. [Google Scholar] [CrossRef] [Green Version]
- Malavaud, S.; Malavaud, B.; Sandres, K.; Durand, D.; Marty, N.; Icart, J.; Rostaing, L.; Malavaud, B. Nosocomial outbreak of influenza virus A (H3N2) infection in a solid organ transplant department. Transplantation 2001, 72, 535–537. [Google Scholar] [CrossRef]
- Taylor, G.S.; Vipond, I.B.; Caul, E.O. Molecular epidemiology of outbreak of respiratory syncytial virus within bone marrow transplantation unit. J. Clin. Microbiol. 2001, 39, 801–803. [Google Scholar] [CrossRef] [Green Version]
- Biernat, M.M.; Zińczuk, A.; Biernat, P.; Bogucka-Fedorczuk, A.; Kwiatkowski, J.; Kalicińska, E.; Marciniak, D.; Simon, K.; Wróbel, T. Nosocomial outbreak of SARS-CoV-2 infection in a haematological unit—High mortality rate in infected patients with haematologic malignancies. J. Clin. Virol. 2020, 130, 104574. [Google Scholar] [CrossRef]
- Jung, J.; Lee, J.; Park, H.; Lim, Y.J.; Kim, E.O.; Park, M.S.; Kim, S.H. Nosocomial Outbreak by Delta Variant from a Fully Vaccinated Patient. J. Korean Med. Sci. 2022, 37, 1–8. [Google Scholar] [CrossRef]
- Cuypers, L.; Bode, J.; Beuselinck, K.; Laenen, L.; Dewaele, K.; Janssen, R.; Capron, A.; Lafort, Y.; Paridaens, H.; Bearzatto, B.; et al. Nationwide Harmonization Effort for Semi-Quantitative Reporting of SARS-CoV-2 PCR Test Results in Belgium. Viruses 2022, 14, 1294. [Google Scholar] [CrossRef]
- Khonyongwa, K.; Taori, S.; Soares, A.; Desai, N.; Sudhanva, M.; Bernal, W.; Schelenz, S.; Curran, L. Incidence and outcomes of healthcare-associated COVID-19 infections: Significance of delayed diagnosis and correlation with staff absence. J. Hosp. Infect. 2020, 106, 663–672. [Google Scholar] [CrossRef]
- Paltansing, S.; Sikkema, R.S.; de Man, S.J.; Koopmans, M.P.G.; Oude Munnink, B.B.; de Man, P. Transmission of SARS-CoV-2 among healthcare workers and patients in a teaching hospital in the Netherlands confirmed by whole-genome sequencing. J. Hosp. Infect. 2021, 110, 178–183. [Google Scholar] [CrossRef]
- Loman, N.; Rowe, W.; Rambaut, A. nCoV-2019 Novel Coronavirus Bioinformatics Protocol. Available online: https://artic.network/ncov-2019/ncov2019-bioinformatics-sop.html (accessed on 3 January 2022).
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef]
- Aksamentov, I.; Roemer, C.; Hodcroft, E.; Neher, R. Nextclade: Clade assignment, mutation calling and quality control for viral genomes. J. Open Source Softw. 2021, 6, 3773. [Google Scholar] [CrossRef]
- Turakhia, Y.; Thornlow, B.; Hinrichs, A.S.; De Maio, N.; Gozashti, L.; Lanfear, R.; Haussler, D.; Corbett-Detig, R. Ultrafast Sample placement on Existing tRees (UShER) enables real-time phylogenetics for the SARS-CoV-2 pandemic. Nat. Genet. 2021, 53, 809–816. [Google Scholar] [CrossRef]
- Severe Acute Respiratory Syndrome Coronavirus 2 Isolate Wuhan-Hu-1, co—Nucleotide—NCBI. Available online: https://www.ncbi.nlm.nih.gov/nuccore/1798174254 (accessed on 26 July 2021).
- Kasson, P.M.; Kamerlin, S.C.L.; Kasson, P.M. Reinfection of Severe Acute Respiratory Syndrome Coronavirus 2 in an Immunocompromised patient: A case report. Clin Infect Dis. 2011, 52, 1–2. Available online: https://github.com/kassonlab/covid19-epi (accessed on 8 June 2022).
- Prado-Vivar, B.; Becerra-Wong, M.; Guadalupe, J.J.; Marquez, S.; Gutierrez, B.; Rojas-Silva, P.; Grunauer, M.; Trueba, G.; Barragan, G.; Cardenas, P. COVID-19 Re-Infection by a Phylogenetically Distinct SARS-CoV-2 Variant, First Confirmed Event in South America. SSRN Electron. J. 2020. [Google Scholar] [CrossRef]
- Van Elslande, J.; Vermeersch, P.; Vandervoort, K.; Wawina-Bokalanga, T.; Vanmechelen, B.; Wollants, E.; Laenen, L.; André, E.; Van Ranst, M.; Lagrou, K.; et al. Symptomatic Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Reinfection by a Phylogenetically Distinct Strain. Clin. Infect. Dis. 2021, 73, 354–356. [Google Scholar] [CrossRef]
- Callow, K.A.; Parry, H.F.; Sergeant, M.; Tyrrell, D.A.J. The time course of the immune response to experimental coronavirus infection of man. Epidemiol. Infect. 1990, 105, 435–446. [Google Scholar] [CrossRef] [Green Version]
- Edridge, A.W.D.; Kaczorowska, J.; Hoste, A.C.R.; Bakker, M.; Klein, M.; Loens, K.; Jebbink, M.F.; Matser, A.; Kinsella, C.M.; Rueda, P.; et al. Seasonal coronavirus protective immunity is short-lasting. Nat Med. 2020, 26, 1691–1693. [Google Scholar] [CrossRef]
- Galanti, M.; Shaman, J. Direct observation of repeated infections with endemic coronaviruses. J. Infect. Dis. 2021, 223, 409–415. [Google Scholar] [CrossRef]
- Goldman, J.D.; Wang, K.; Roltgen, K.; Nielsen, S.C.A.; Roach, J.C.; Naccache, S.N.; Yang, F.; Wirz, O.F.; Yost, K.E.; Lee, J.-Y.; et al. Reinfection with SARS-CoV-2 and Failure of Humoral Immunity: A case report. medRxiv Prepr. Serv. Health Sci. 2020, 98104, 1–21. Available online: http://www.ncbi.nlm.nih.gov/pubmed/32995830%0Ahttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC7523175 (accessed on 8 June 2022).
- Bok, K.; Prevots, D.R.; Binder, A.M.; Parra, G.I.; Strollo, S.; Fahle, G.A.; Behrle-Yardley, A.; Johnson, J.A.; Levenson, E.A.; Sosnovtsev, S.V.; et al. Epidemiology of norovirus infection among immunocompromised patients at a tertiary care research hospital, 2010–2013. Open Forum. Infect. Dis. 2016, 3, ofw169. [Google Scholar] [CrossRef] [Green Version]
- Lumby, C.K.; Zhao, L.; Oporto, M.; Best, T.; Tutill, H.; Shah, D.; Veys, P.; Williams, R.; Worth, A.; Illingworth, C.J.R.; et al. Favipiravir and zanamivir cleared infection with influenza B in a severely immunocompromised child. Clin. Infect. Dis. 2020, 71, E191–E194. [Google Scholar] [CrossRef]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.-H.; Boucau, J.; Bowman, K.; et al. Persistence and Evolution of SARS-CoV-2 in an Immunocompromised Host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef]
- Baang, J.H.; Smith, C.; Mirabelli, C.; Valesano, A.L.; Manthei, D.M.; Bachman, M.A.; Wobus, C.E.; Adams, M.; Washer, L.; Martin, E.T.; et al. Prolonged Severe Acute Respiratory Syndrome Coronavirus 2 Replication in an Immunocompromised Patient. J. Infect Dis. 2021, 223, 23–27. [Google Scholar] [CrossRef]
- Meredith, L.W.; Hamilton, W.L.; Warne, B.; Houldcroft, C.J.; Hosmillo, M.; Jahun, A.S.; Curran, M.D.; Parmar, S.; Caller, L.G.; Caddy, S.L.; et al. Rapid implementation of SARS-CoV-2 sequencing to investigate cases of health-care associated COVID-19: A prospective genomic surveillance study. Lancet Infect. Dis. 2020, 20, 1263–1271. [Google Scholar] [CrossRef]
- Thielen, B.K.; Bye, E.; Wang, X.; Maroushek, S.; Friedlander, H.; Bistodeau, S.; Christensen, J.; Reisdorf, E.; Shilts, M.H.; Martin, K.; et al. Summer Outbreak of Severe RSV-B Disease, Minnesota, 2017 Associated with Emergence of a Genetically Distinct Viral Lineage. J. Infect. Dis. 2020, 222, 288–297. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.; Motta, F.; Isailovic, N.; Clementi, M.; Criscuolo, E.; Clementi, N.; Tonutti, A.; Rodolfi, S.; Barone, E.; Colapietro, F.; et al. Dose-Dependent Impairment of the Immune Response to the Moderna-1273 mRNA Vaccine by Mycophenolate Mofetil in Patients with Rheumatic and Autoimmune Liver Diseases. Vaccines 2022, 10, 801. [Google Scholar] [CrossRef] [PubMed]
- Boekel, L.; Wolbink, G.J. Rituximab during the COVID-19 pandemic: Time to discuss treatment options with patients. Lancet Rheumatol. 2022, 4, e154–e155. [Google Scholar] [CrossRef]
- Ong, D.; Fragkou, P.C.; Schweitzer, V.A.; Chemaly, R.F.; Moschopoulos, C.D.; Skevaki, C.; European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Study Group for Respiratory Viruses (ESGREV). How to interpret and use COVID-19 serology and immunology tests. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2021, 27, 981–986. [Google Scholar] [CrossRef]
- Bitoun, S.; Henry, J.; Desjardins, D.; Vauloup-Fellous, C.; Dib, N.; Belkhir, R.; Mouna, L.; Joly, C.; Bitu, M.; Ly, B.; et al. Rituximab Impairs B Cell Response but Not T Cell Response to COVID-19 Vaccine in Autoimmune Diseases. Arthritis Rheumatol. 2022, 74, 927–933. [Google Scholar] [CrossRef]
- Jyssum, I.; Kared, H.; Tran, T.T.; Tveter, A.T.; Provan, S.A.; Sexton, J.; Jørgensen, K.K.; Jahnsen, J.; Kro, G.B.; Warren, D.J.; et al. Humoral and cellular immune responses to two and three doses of SARS-CoV-2 vaccines in rituximab-treated patients with rheumatoid arthritis: A prospective, cohort study. Lancet Rheumatol. 2022, 4, e177–e187. [Google Scholar] [CrossRef]
- Levavi, H.; Lancman, G.; Gabrilove, J. Impact of rituximab on COVID-19 outcomes. Ann. Hematol. 2021, 100, 2805–2812. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BA.1 Like Characteristic Base Variations | BA.2 Like Characteristic Base Variations | ||||
|---|---|---|---|---|---|
| A | B | A | B | ||
| nuc:T5386G | + | − | ORF1ab:S135R | − | + |
| del: 6513:3 | + | − | ORF1ab:T842I | − | + |
| ORF1ab:A2710T | + | − | ORF1ab:G1307S | − | + |
| ORF1ab:L3674F | − | − | nuc:C4321T | − | + |
| ORF1ab:SGF3675- | − | − | ORF1ab:L3027F | − | + |
| ORF1ab:I3758V | + | − | nuc:A9424G | − | + |
| nuc:T13195C | + | − | ORF1ab:T3090I | − | + |
| nuc:C15240T | + | − | ORF1ab:L3201F | − | + |
| S:A67V | + | − | ORF1ab:SGF3675- | − | + |
| del:21765:6 | + | − | nuc:C10198T | − | + |
| S:T95I | + | − | nuc:G10447A | − | + |
| del:21987:9 | + | + | nuc:C12880T | − | + |
| del:22194:3 | + | − | nuc:C15714T | − | + |
| nuc:22205 + GAGCCAGAA | + | − | ORF1ab:R5716C | − | − |
| S:S371L | + | − | ORF1ab:T6564I | − | − |
| S:G446S | + | − | nuc:A20055G | − | + |
| S:Q493R | + | + | S:T19I | − | + |
| S:G496S | + | − | del:21633:9 | − | + |
| S:T547K | + | − | nuc:T22200G | − | + |
| S:N856K | + | − | S:S371F | − | + |
| S:L981F | + | − | S:T376A | − | + |
| nuc:C25000T | + | + | S:D405N | − | + |
| nuc:C25584T | + | + | S:R408S | − | + |
| M:D3G | + | − | S:Q493R | + | + |
| N:RG203KR | + | + | nuc:C25000T | + | + |
| nuc:C25584T | + | + | |||
| nuc:C26060T | − | + | |||
| nuc:C26858T | − | + | |||
| ORF6:D61L | − | + | |||
| N:RG203KR | + | + | |||
| N:S413R | − | + | |||
| Day * | Anti-Nucleocapsid IgG | Anti-Spike Protein IgG |
|---|---|---|
| 0 | Negative | Negative |
| 14 | Negative | Negative |
| 37 | Negative | Negative |
| 157 | Negative | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callebaut, K.; Stoefs, A.; Stylemans, D.; Soetens, O.; Crombé, F.; Vancutsem, E.; Imamura, H.; Wybo, I.; De Geyter, D.; Piérard, D.; et al. Healthcare-Associated SARS-CoV-2 Reinfection after 3 Months with a Phylogenetically Distinct Omicron Variant: A Case Report. Viruses 2022, 14, 1852. https://doi.org/10.3390/v14091852
Callebaut K, Stoefs A, Stylemans D, Soetens O, Crombé F, Vancutsem E, Imamura H, Wybo I, De Geyter D, Piérard D, et al. Healthcare-Associated SARS-CoV-2 Reinfection after 3 Months with a Phylogenetically Distinct Omicron Variant: A Case Report. Viruses. 2022; 14(9):1852. https://doi.org/10.3390/v14091852
Chicago/Turabian StyleCallebaut, Kim, Anke Stoefs, Dimitri Stylemans, Oriane Soetens, Florence Crombé, Ellen Vancutsem, Hideo Imamura, Ingrid Wybo, Deborah De Geyter, Denis Piérard, and et al. 2022. "Healthcare-Associated SARS-CoV-2 Reinfection after 3 Months with a Phylogenetically Distinct Omicron Variant: A Case Report" Viruses 14, no. 9: 1852. https://doi.org/10.3390/v14091852