

Neuropeptides, New Ligands of SARS-CoV-2 Nucleoprotein, a Potential Link between Replication, Inflammation and Neurotransmission
 , and
, and
Abstract
:1. Introduction
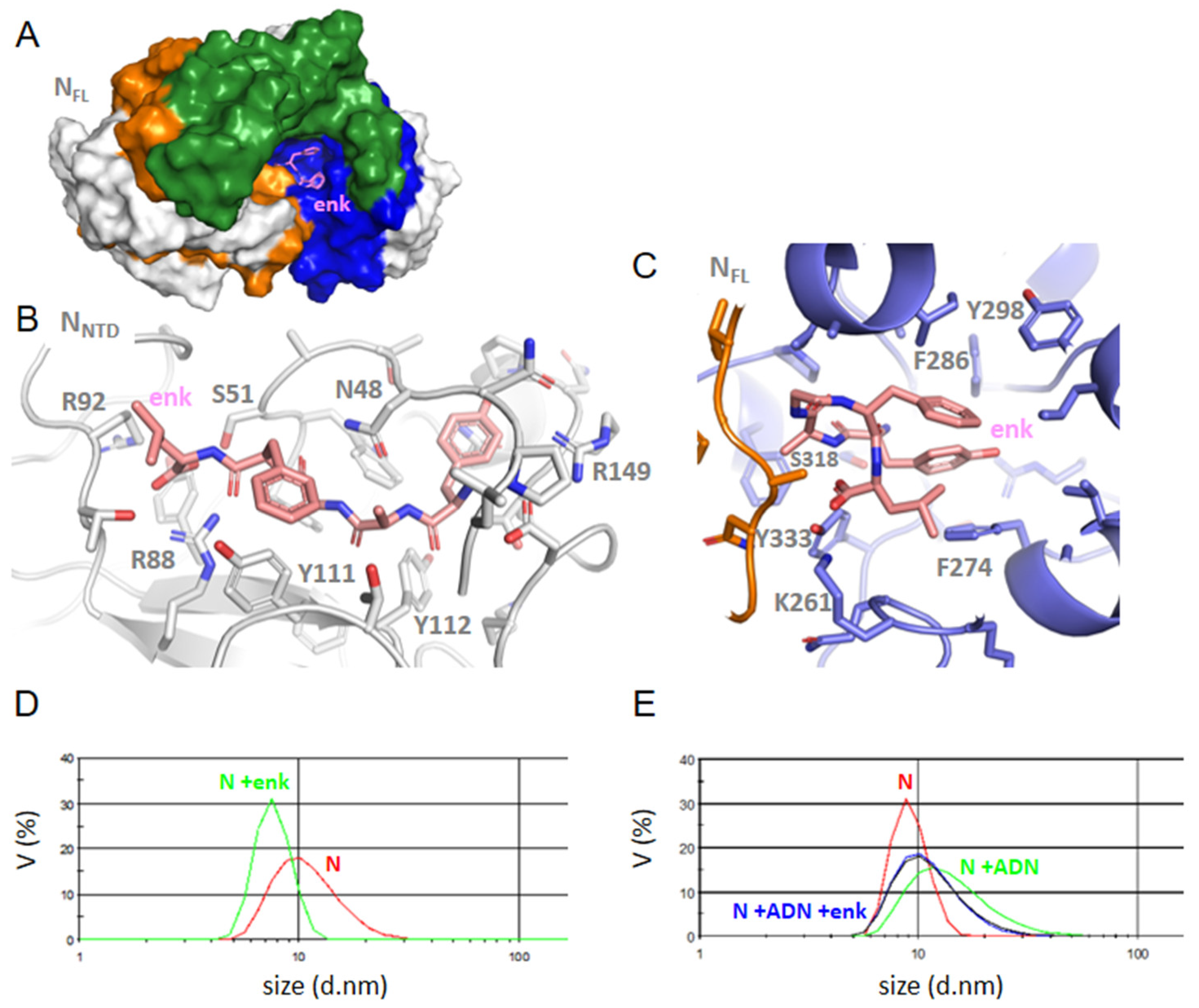
2. Results
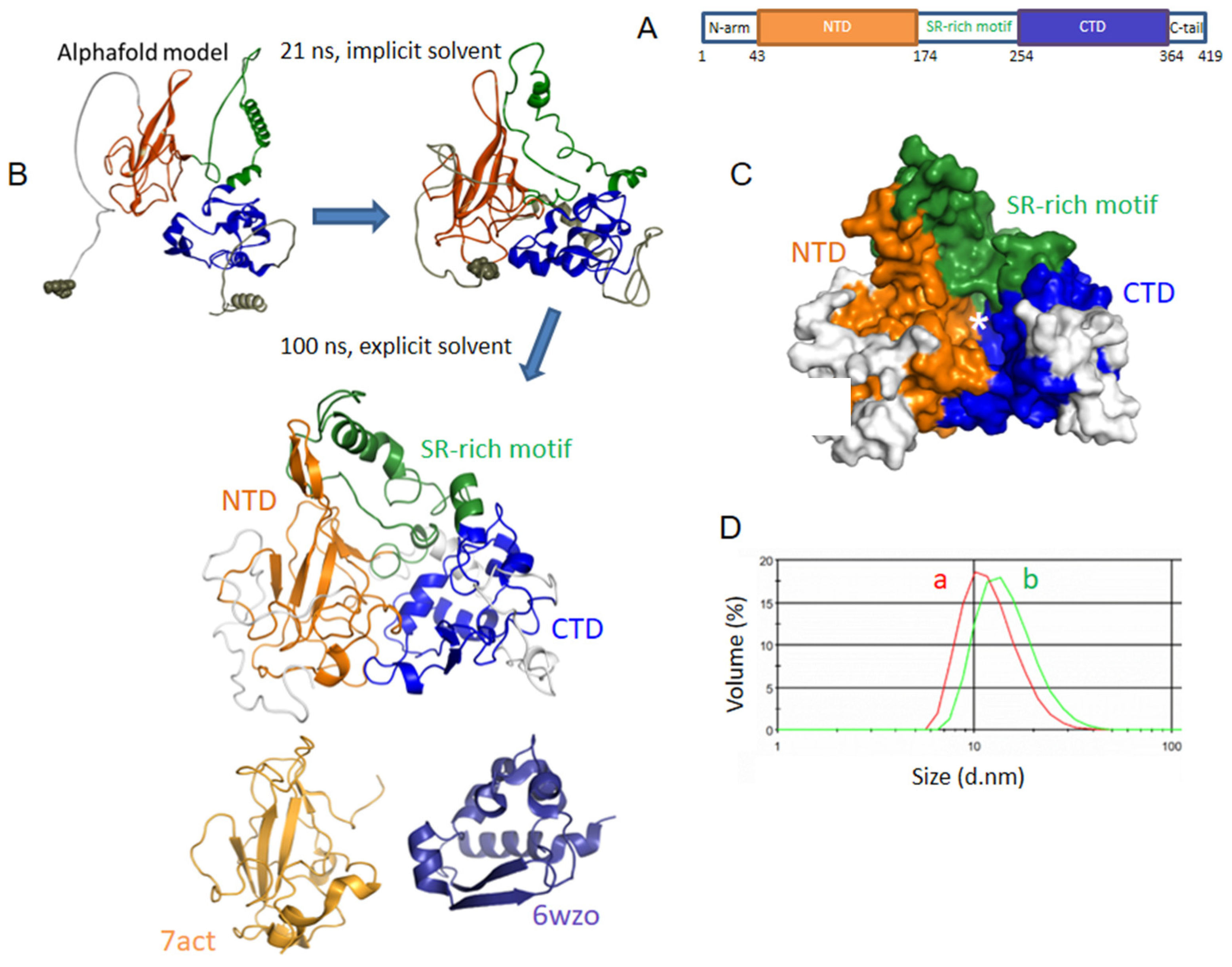
2.1. Generation of Models of Full-Length Nucleoprotein N


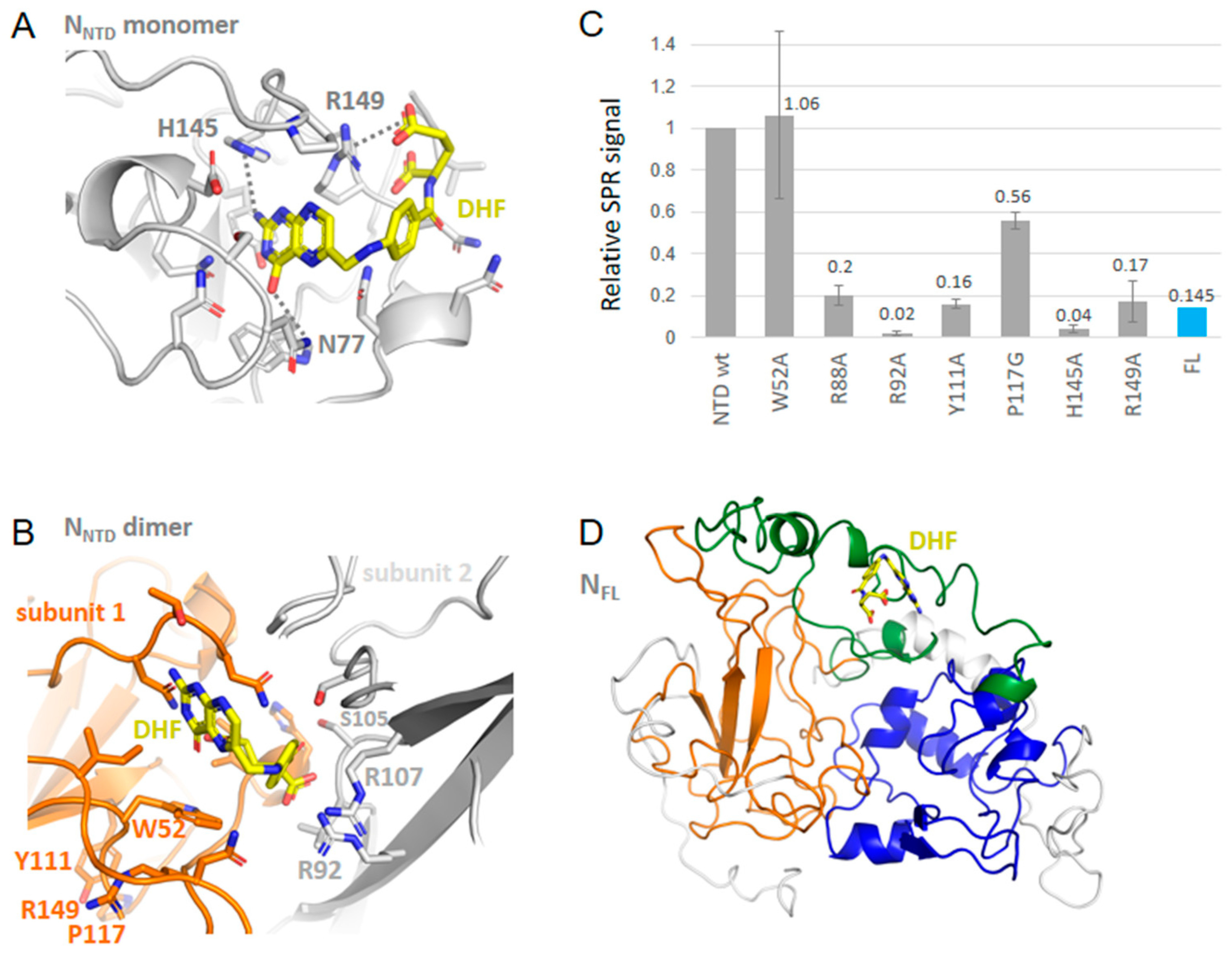
2.2. Characterization of Recombinant N, N-NTD and Its Mutants and N Full Length N-FL
2.3. Virtual Drug Screening on NTD and FL
- Ligands involved in metabolism
- Ligands involved in the COX-prostaglandins pathway:
- Neuropeptides:
- Hemin in FL:
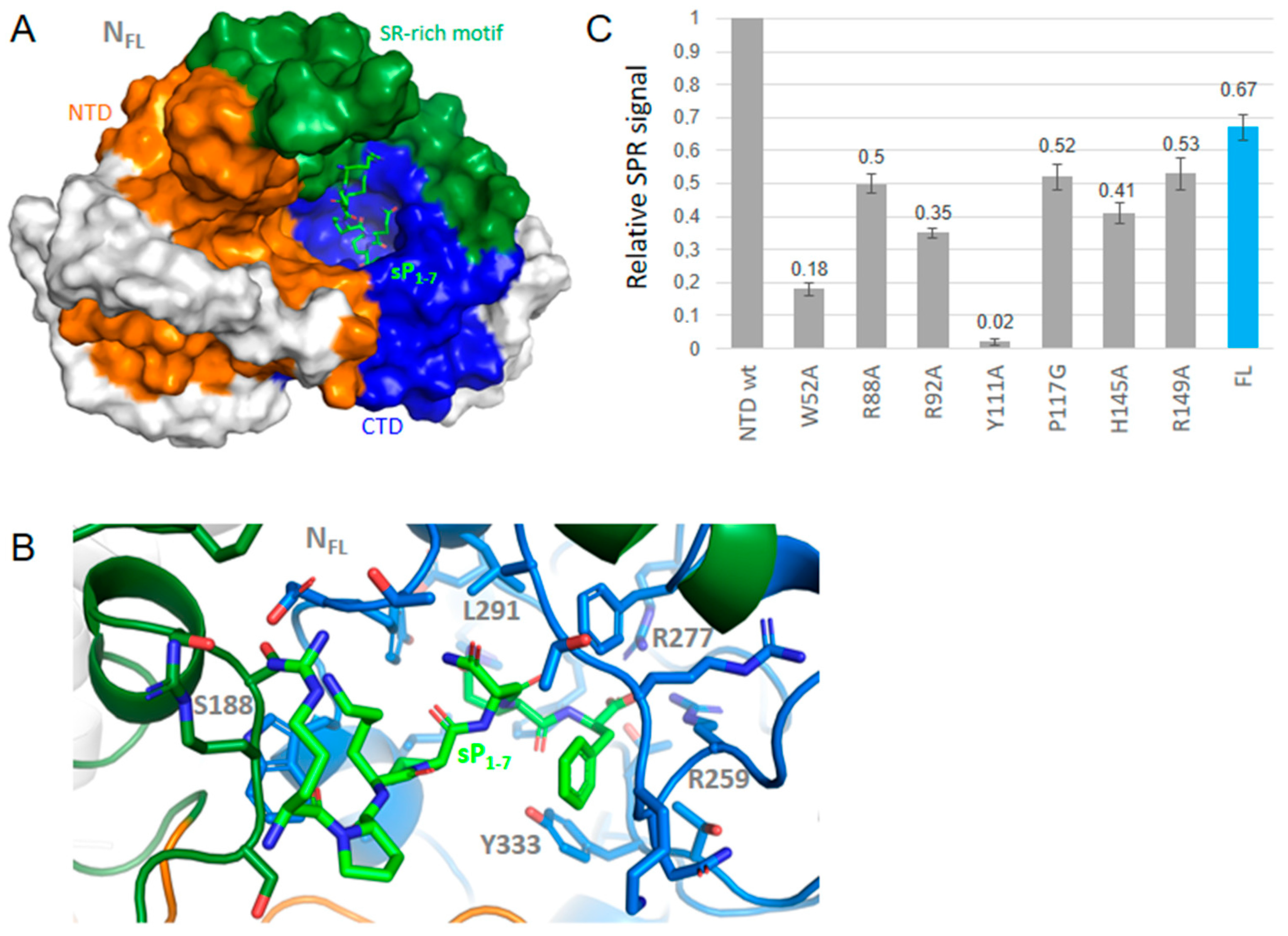
2.4. SPR Study of DHF and Substance P (1-7) Interactions with NTD WT and Mutants, and FL
2.5. FL Interactions with the Ligands by DLS; Competition with Single-Stranded DNA or RNA Binding
2.6. FL Interactions with ZnTPPS
3. Discussion
3.1. N as a Mediator of Inflammation and Its Effect in Long-Haul COVID-19
3.2. N Implication in Immunity, with Possible Long-Term Neurological Effects and Putative Viral Latency
- (i)
- T cells: Patients with persistent symptoms over 4 months following COVID-19 onset presented a lower frequency of CD8+ T cells expressing CD107a, a marker of degranulation, in response to Nucleocapsid (N) peptide pool stimulation, and a more rapid decline in the frequency of N-specific interferon-γ-producing CD8+ T cells [10].
- (ii)
- Opioid peptides: Immune system and neuronal system cross-talk; this signaling is mediated by various molecules such as opioid peptides such as enkephalin. Enkephalins can impact lymphocytes proliferation, antibody synthesis. Enkephalin can enhance the release of pro-inflammatory cytokines like IL6 [50]. The cross-talk can take place between cytokines as CCL2 and opioid peptides and alter nociceptive synaptic transmission [51]. Thus, more work is required to test whether the enkephalin peptide identified here as a ligand of N may increase pain and immune disorders in the context of COVID-19, despite its physiological pain release function.
- (iii)
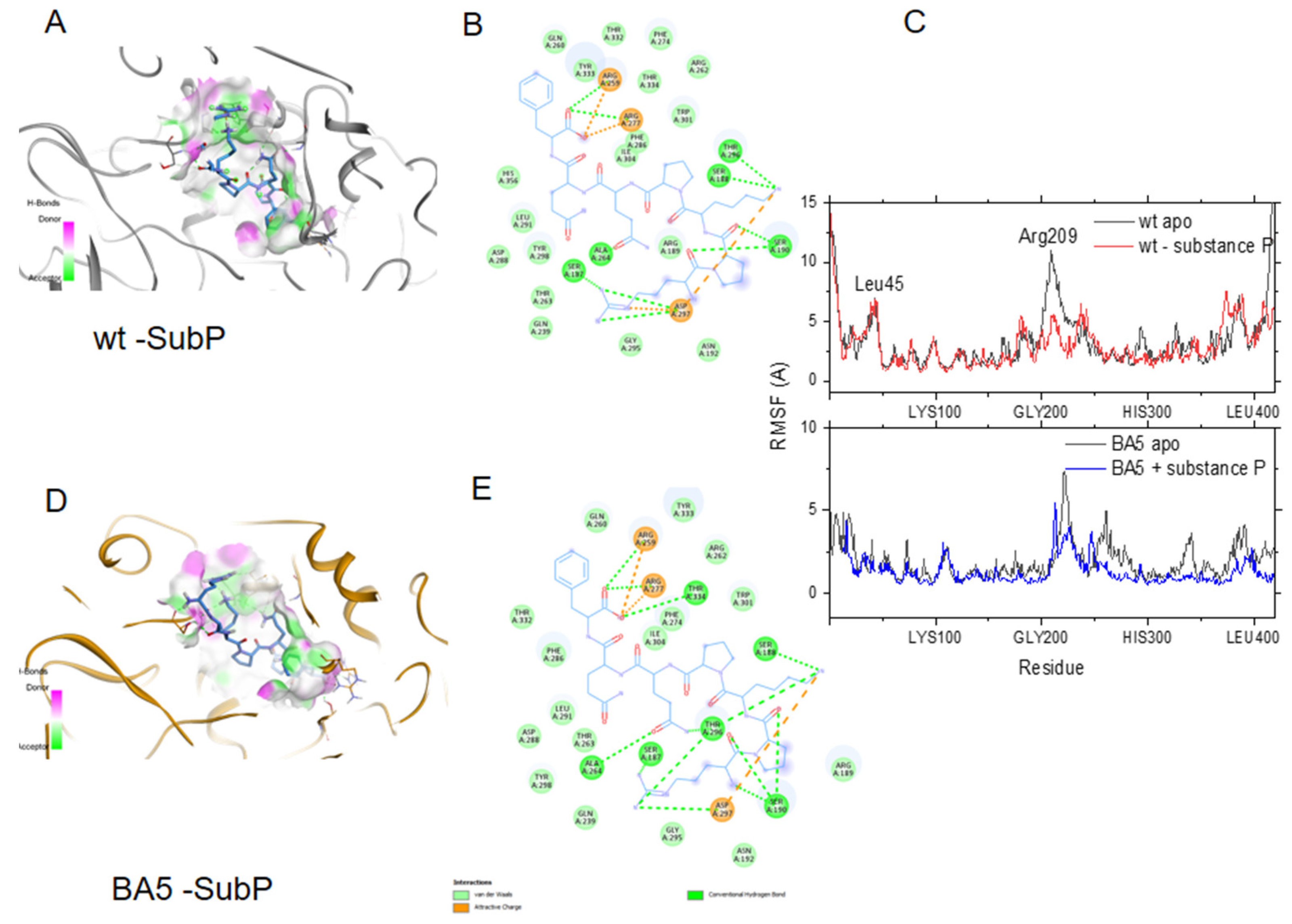
- Vasoactive peptides: Among the possible routes through which SARS-CoV-2 can invade the CNS, SARS-CoV-2 can directly invade the vagus nerve and retrograde into the CNS, or indirectly stimulate the enteric nervous system through immune pathways. Cholecystokinin could participate in this process. In addition, peripheral nerves may spread SARS-CoV-2 into the brain through the retro-neural route, including the olfactory nerve, trigeminal nerve, glossopharyngeal nerve and vagus nerve. Substance P is the main neuropeptide, neuromodulator and neuro-hormone of the trigeminal ganglion (TG), associated with nociception and inflammation through its receptor, the neurokinin-1. As observed with other viruses such as herpes or HIV, it was hypothesized that SARS-CoV-2 virus infection might become latent if it is acting through TG [41,45], involving substance P action.
3.3. N as a Mediator of Perturbed Metabolism via Its Ligands, with Possible Long-Term Effects
3.4. Zntpps as a Prototype of a Photoactive N Ligand for Photodynamic Therapy (PDT)
3.5. Heme Sequestering by N and Its Potential Effect on NO and ROS Signaling
4. Materials and Methods
4.1. Models of N, in Silico Screening, Docking and Molecular Dynamics Simulations
4.1.1. Models of wt N
4.1.2. Model of N Variants
4.1.3. Ligand Screening and Docking to wt N
4.1.4. Substance P (1-7) Bound to BA5 Variant
4.2. Chemicals and Oligonucleotides
4.3. Proteins Expression and Purification
4.4. SPR Experiments
4.5. Dynamic Light Scattering
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Dedication
References
- Cubuk, J.; Alston, J.J.; Incicco, J.J.; Singh, S.; Stuchell-Brereton, M.D.; Ward, M.D.; Zimmerman, M.I.; Vithani, N.; Griffith, D.; Wagoner, J.A.; et al. The SARS-CoV-2 nucleocapsid protein is dynamic, disordered, and phase separates with RNA. Nat. Commun. 2021, 12, 1936. [Google Scholar] [CrossRef]
- Dinesh, D.C.; Chalupska, D.; Silhan, J.; Koutna, E.; Nencka, R.; Veverka, V.; Boura, E. Structural basis of RNA recognition by the SARS-CoV-2 nucleocapsid phosphoprotein. PLoS Pathog. 2020, 16, e1009100. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Yang, M.; Hong, Z.; Zhang, L.; Huang, Z.; Chen, X.; He, S.; Zhou, Z.; Zhou, Z.; Chen, Q.; et al. Crystal structure of SARS-CoV-2 nucleocapsid protein RNA binding domain reveals potential unique drug targeting sites. Acta Pharm. Sin. B 2020, 10, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; West, A.M.V.; Silletti, S.; Corbett, K.D. Architecture and self-assembly of the SARS-CoV-2 nucleocapsid protein. Protein Sci. 2020, 29, 1890–1901. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.; Cortese, M.; Winter, S.L.; Wachsmuth-Melm, M.; Neufeldt, C.J.; Cerikan, B.; Stanifer, M.L.; Boulant, S.; Bartenschlager, R.; Chlanda, P. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nat. Commun. 2020, 11, 5885. [Google Scholar] [CrossRef]
- Chen, K.; Xiao, F.; Hu, D.; Ge, W.; Tian, M.; Wang, W.; Pan, P.; Wu, K.; Wu, J. SARS-CoV-2 Nucleocapsid Protein Interacts with RIG-I and Represses RIG-Mediated IFN-β Production. Viruses 2021, 13, 47. [Google Scholar] [CrossRef]
- Mu, J.; Xu, J.; Zhang, L.; Shu, T.; Wu, D.; Huang, M.; Ren, Y.; Li, X.; Geng, Q.; Xu, Y.; et al. SARS-CoV-2-encoded nucleocapsid protein acts as a viral suppressor of RNA interference in cells. Sci. China Life Sci. 2020, 63, 1413–1416. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Ye, Q.; Singh, D.; Cao, Y.; Diedrich, J.K.; Yates, J.R., 3rd; Villa, E.; Cleveland, D.W.; Corbett, K.D. The SARS-CoV-2 nucleocapsid phosphoprotein forms mutually exclusive condensates with RNA and the membrane-associated M protein. Nat. Commun. 2021, 12, 502. [Google Scholar] [CrossRef]
- Peluso, M.J.; Deitchman, A.N.; Torres, L.; Iyer, N.S.; Munter, S.E.; Nixon, C.C.; Donatelli, J.; Thanh, C.; Takahashi, S.; Hakim, J.; et al. Long-term SARS-CoV-2-specific immune and inflammatory responses in individuals recovering from COVID-19 with and without post-acute symptoms. Cell Rep. 2021, 36, 109518. [Google Scholar] [CrossRef]
- Qian, Y.; Lei, T.; Patel, P.S.; Lee, C.H.; Monaghan-Nichols, P.; Xin, H.-B.; Qiu, J.; Fu, M. Direct Activation of Endothelial Cells by SARS-CoV-2 Nucleocapsid Protein Is Blocked by Simvastatin. J. Virol. 2021, 95, e01396-21. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Cao, Y.; Liu, W.; Li, J. The SARS-CoV-2 Nucleocapsid Protein and Its Role in Viral Structure, Biological Functions, and a Potential Target for Drug or Vaccine Mitigation. Viruses 2021, 13, 1115. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Alfajaro, M.M.; Chow, R.D.; Wei, J.; Filler, R.B.; Eisenbarth, S.C.; Wilen, C.B. Nonsteroidal Anti-inflammatory Drugs Dampen the Cytokine and Antibody Response to SARS-CoV-2 Infection. J. Virol. 2021, 95, e00014-21. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Hao, Q.; Mu, Y.; Timani, K.A.; Ye, L.; Zhu, Y.; Wu, J. Nucleocapsid protein of SARS-CoV activates the expression of cyclooxygenase-2 by binding directly to regulatory elements for nuclear factor-kappa B and CCAAT/enhancer binding protein. Int. J. Biochem. Cell Biol. 2006, 38, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Tang, W.; Wang, J.; Lai, D.; Xu, Q.; Huang, R.; Hu, Y.; Gong, X.; Fan, J.; Shu, Q.; et al. SARS-CoV-2 N Protein Induces Acute Lung Injury in Mice via NF-ĸB Activation. Front. Immunol. 2021, 12, 791753. [Google Scholar] [CrossRef]
- Perico, N.; Cortinovis, M.; Suter, F.; Remuzzi, G. Home as the new frontier for the treatment of COVID-19: The case for anti-inflammatory agents. Lancet Infect. Dis. 2022. [Google Scholar] [CrossRef]
- Terrier, O.; Dilly, S.; Pizzorno, A.; Chalupska, D.; Humpolickova, J.; Bouřa, E.; Berenbaum, F.; Quideau, S.; Lina, B.; Fève, B.; et al. Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus. Molecules 2021, 26, 2593. [Google Scholar] [CrossRef]
- Terrier, O.; Dilly, S.; Pizzorno, A.; Henri, J.; Berenbaum, F.; Lina, B.; Fève, B.; Adnet, F.; Sabbah, M.; Rosa-Calatrava, M.; et al. Broad-spectrum antiviral activity of naproxen: From Influenza A to SARS-CoV-2 Coronavirus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dilly, S.; Fotso Fotso, A.; Lejal, N.; Zedda, G.; Chebbo, M.; Rahman, F.; Companys, S.; Bertrand, H.C.; Vidic, J.; Noiray, M.; et al. From Naproxen Repurposing to Naproxen Analogues and Their Antiviral Activity against Influenza A Virus. J. Med. Chem. 2018, 61, 7202–7217. [Google Scholar] [CrossRef] [Green Version]
- Lejal, N.; Tarus, B.; Bouguyon, E.; Chenavas, S.; Bertho, N.; Delmas, B.; Ruigrok, R.W.; Di Primo, C.; Slama-Schwok, A. Structure-based discovery of the novel antiviral properties of naproxen against the nucleoprotein of influenza A virus. Antimicrob. Agents Chemother. 2013, 57, 2231–2242. [Google Scholar] [CrossRef]
- Rutkai, I.; Mayer, M.G.; Hellmers, L.M.; Ning, B.; Huang, Z.; Monjure, C.J.; Coyne, C.; Silvestri, R.; Golden, N.; Hensley, K.; et al. Neuropathology and virus in brain of SARS-CoV-2 infected non-human primates. Nat. Commun. 2022, 13, 1745. [Google Scholar] [CrossRef] [PubMed]
- Thompson, E.J.; Williams, D.M.; Walker, A.J.; Mitchell, R.E.; Niedzwiedz, C.L.; Yang, T.C.; Huggins, C.F.; Kwong, A.S.F.; Silverwood, R.J.; Di Gessa, G.; et al. Long COVID burden and risk factors in 10 UK longitudinal studies and electronic health records. Nat. Commun. 2022, 13, 3528. [Google Scholar] [CrossRef] [PubMed]
- Guseva, S.; Perez, L.M.; Camacho-Zarco, A.; Bessa, L.M.; Salvi, N.; Malki, A.; Maurin, D.; Blackledge, M. (1)H, (13)C and (15)N Backbone chemical shift assignments of the n-terminal and central intrinsically disordered domains of SARS-CoV-2 nucleoprotein. Biomol. NMR Assign. 2021, 15, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.H.; Khan, N.; Mishra, A.; Gupta, S.; Bansode, V.; Mehta, D.; Bhambure, R.; Ansari, M.A.; Das, S.; Rathore, A.S. Dimerization of SARS-CoV-2 nucleocapsid protein affects sensitivity of ELISA based diagnostics of COVID-19. Int. J. Biol. Macromol. 2022, 200, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Du, N.; Lei, Y.; Dorje, S.; Qi, J.; Luo, T.; Gao, G.F.; Song, H. Structures of the SARS-CoV-2 nucleocapsid and their perspectives for drug design. EMBO J. 2020, 39, e105938. [Google Scholar] [CrossRef] [PubMed]
- Di, D.; Dileepan, M.; Ahmed, S.; Liang, Y.; Ly, H. Recombinant SARS-CoV-2 Nucleocapsid Protein: Expression, Purification, and Its Biochemical Characterization and Utility in Serological Assay Development to Assess Immunological Responses to SARS-CoV-2 Infection. Pathogens 2021, 10, 1039. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Yang, M.; He, S.; Chen, X.; Huang, Z.; Zhou, Z.; Zhou, Z.; Chen, Q.; Chen, S.; Kang, S. Structural Insight Into the SARS-CoV-2 Nucleocapsid Protein C-Terminal Domain Reveals a Novel Recognition Mechanism for Viral Transcriptional Regulatory Sequences. Front. Chem. 2020, 8, 624765. [Google Scholar] [CrossRef]
- Peronace, C.; Tallerico, R.; Colosimo, M.; Fazio, M.; Pasceri, F.; Talotta, I.; Panduri, G.; Pintomalli, L.; Oteri, R.; Calantoni, V.; et al. The First Identification in Italy of SARS-CoV-2 Omicron BA.4 Harboring KSF141_del: A Genomic Comparison with Omicron Sub-Variants. Biomedicines 2022, 10, 1839. [Google Scholar] [CrossRef]
- Wu, H.; Xing, N.; Meng, K.; Fu, B.; Xue, W.; Dong, P.; Tang, W.; Xiao, Y.; Liu, G.; Luo, H.; et al. Nucleocapsid mutations R203K/G204R increase the infectivity, fitness, and virulence of SARS-CoV-2. Cell Host Microbe 2021, 29, 1788–1801.e6. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Liu, G.; Ma, H.; Zhao, D.; Yang, Y.; Liu, M.; Mohammed, A.; Zhao, C.; Yang, Y.; Xie, J.; et al. Biochemical characterization of SARS-CoV-2 nucleocapsid protein. Biochem. Biophys. Res. Commun. 2020, 527, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Lechuga, G.C.; Souza-Silva, F.; Sacramento, C.Q.; Trugilho, M.R.O.; Valente, R.H.; Napoleao-Pego, P.; Dias, S.S.G.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Carels, N.; et al. SARS-CoV-2 Proteins Bind to Hemoglobin and Its Metabolites. Int. J. Mol. Sci. 2021, 22, 9035. [Google Scholar] [CrossRef] [PubMed]
- Panagiotopoulos, A.; Tseliou, M.; Karakasiliotis, I.; Kotzampasi, D.M.; Daskalakis, V.; Kesesidis, N.; Notas, G.; Lionis, C.; Kampa, M.; Pirintsos, S.; et al. p-cymene impairs SARS-CoV-2 and Influenza A (H1N1) viral replication: In Silico predicted interaction with SARS-CoV-2 nucleocapsid protein and H1N1 nucleoprotein. Pharm. Res. Perspect. 2021, 9, e00798. [Google Scholar] [CrossRef] [PubMed]
- Amici, C.; Di Caro, A.; Ciucci, A.; Chiappa, L.; Castilletti, C.; Martella, V.; Decaro, N.; Buonavoglia, C.; Capobianchi, M.R.; Santoro, M.G. Indomethacin has a potent antiviral activity against SARS coronavirus. Antivir. Ther. 2006, 11, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. SARS-CoV-2 hijacks folate and one-carbon metabolism for viral replication. Nat. Commun. 2021, 12, 1676. [Google Scholar] [CrossRef] [PubMed]
- Frere, J.J.; Serafini, R.A.; Pryce, K.D.; Zazhytska, M.; Oishi, K.; Golynker, I.; Panis, M.; Zimering, J.; Horiuchi, S.; Hoagland, D.A.; et al. SARS-CoV-2 infection in hamsters and humans results in lasting and unique systemic perturbations post recovery. Sci. Transl. Med. 2022, 14, eabq3059. [Google Scholar] [CrossRef]
- Ricke-Hoch, M.; Stelling, E.; Lasswitz, L.; Gunesch, A.P.; Kasten, M.; Zapatero-Belinchon, F.J.; Brogden, G.; Gerold, G.; Pietschmann, T.; Montiel, V.; et al. Impaired immune response mediated by prostaglandin E2 promotes severe COVID-19 disease. PLoS ONE 2021, 16, e0255335. [Google Scholar] [CrossRef]
- Kishk, A.; Pacheco, M.P.; Sauter, T. DCcov: Repositioning of drugs and drug combinations for SARS-CoV-2 infected lung through constraint-based modeling. iScience 2021, 24, 103331. [Google Scholar] [CrossRef]
- Ravichandran, R.; Mohan, S.K.; Sukumaran, S.K.; Kamaraj, D.; Daivasuga, S.S.; Ravi, S.; Vijayaraghavalu, S.; Kumar, R.K. Author Correction: An open label randomized clinical trial of Indomethacin for mild and moderate hospitalised COVID-19 patients. Sci. Rep. 2022, 12, 10389, Correction in Sci. Rep. 2022, 12, 6413. [Google Scholar] [CrossRef] [PubMed]
- Shah, F.H.; Lim, K.H.; Kim, S.J. Do fever-relieving medicines have anti-COVID activity: An in silico insight. Future Virol. 2021, 16, 293–300. [Google Scholar] [CrossRef]
- Mehboob, R.; Kurdi, M.; Bamaga, A.; Aldardeir, N.; Nasief, H.; Moshref, L.H.; Alsinani, T.; Rayes, A.O.; Jabbad, R.H. Substance P/Neurokinin-1 Receptor, Trigeminal Ganglion, Latency, and Coronavirus Infection-Is There Any Link? Front. Med. 2021, 8, 727593. [Google Scholar] [CrossRef] [PubMed]
- Salina, A.C.G.; Dos-Santos, D.; Rodrigues, T.S.; Fortes-Rocha, M.; Freitas-Filho, E.G.; Alzamora-Terrel, D.L.; Castro, I.M.S.; Fraga da Silva, T.F.C.; de Lima, M.H.F.; Nascimento, D.C.; et al. Efferocytosis of SARS-CoV-2-infected dying cells impairs macrophage anti-inflammatory functions and clearance of apoptotic cells. eLife 2022, 11, e74443. [Google Scholar] [CrossRef] [PubMed]
- Chau, C.W.; Sugimura, R. COVID-19: Locked in a pro-inflammatory state. eLife 2022, 11, e80699. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.A.; Faust, B.; Gondin, A.B.; Damgen, M.A.; Suomivuori, C.M.; Veldhuis, N.A.; Cheng, Y.; Dror, R.O.; Thal, D.M.; Manglik, A. Selective G protein signaling driven by substance P-neurokinin receptor dynamics. Nat. Chem. Biol. 2022, 18, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Mehboob, R.; Lavezzi, A.M. Neuropathological explanation of minimal COVID-19 infection rate in newborns, infants and children—A mystery so far. New insight into the role of Substance P. J. Neurol. Sci. 2021, 420, 117276. [Google Scholar] [CrossRef] [PubMed]
- Corder, G.; Castro, D.C.; Bruchas, M.R.; Scherrer, G. Endogenous and Exogenous Opioids in Pain. Annu. Rev. Neurosci. 2018, 41, 453–473. [Google Scholar] [CrossRef]
- Wang, Q.; Chu, C.H.; Qian, L.; Chen, S.H.; Wilson, B.; Oyarzabal, E.; Jiang, L.; Ali, S.; Robinson, B.; Kim, H.C.; et al. Substance P exacerbates dopaminergic neurodegeneration through neurokinin-1 receptor-independent activation of microglial NADPH oxidase. J. Neurosci. Off. J. Soc. Neurosci. 2014, 34, 12490–12503. [Google Scholar] [CrossRef] [Green Version]
- Karamyan, V.T. Between two storms, vasoactive peptides or bradykinin underlie severity of COVID-19? Physiol. Rep. 2021, 9, e14796. [Google Scholar] [CrossRef]
- Lattmann, E.; Bhalerao, P.; ShashiBhushan, B.L.; Nargundkar, N.; Lattmann, P.; Pillai, K.S.; Balaram, P.N. Randomized, Comparative, Clinical Trial to Evaluate Efficacy and Safety of PNB001 in Moderate COVID-19 Patients. medRxiv 2021. [Google Scholar] [CrossRef]
- Singh, V.; Allawadhi, P.; Khurana, A.; Banothu, A.K.; Bharani, K.K. Critical neurological features of COVID-19: Role of imaging methods and biosensors for effective diagnosis. Sens. Int. 2021, 2, 100098. [Google Scholar] [CrossRef]
- Heles, M.; Mrozkova, P.; Sulcova, D.; Adamek, P.; Spicarova, D.; Palecek, J. Chemokine CCL2 prevents opioid-induced inhibition of nociceptive synaptic transmission in spinal cord dorsal horn. J. Neuroinflammation 2021, 18, 279. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Chalupsky, K.; Stefani, E.; Cai, H. Mechanistic insights into folic acid-dependent vascular protection: Dihydrofolate reductase (DHFR)-mediated reduction in oxidant stress in endothelial cells and angiotensin II-infused mice: A novel HPLC-based fluorescent assay for DHFR activity. J. Mol. Cell. Cardiol. 2009, 47, 752–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oak, J.H.; Cai, H. Attenuation of angiotensin II signaling recouples eNOS and inhibits nonendothelial NOX activity in diabetic mice. Diabetes 2007, 56, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.; Tanaka, K.; Hirota, Y. Immune response to influenza vaccine in healthy adults and the elderly: Association with nutritional status. Vaccine 2005, 23, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Tang, Y.S.; Khan, R.A.; Zhang, Y.; Kusumanchi, P.; Stabler, S.P.; Jayaram, H.N.; Antony, A.C. Influence of physiologic folate deficiency on human papillomavirus type 16 (HPV16)-harboring human keratinocytes in vitro and in vivo. J. Biol. Chem. 2012, 287, 12559–12577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziganshyna, S.; Szczepankiewicz, G.; Kuehnert, M.; Schulze, A.; Liebert, U.G.; Pietsch, C.; Eulenburg, V.; Werdehausen, R. Photodynamic Inactivation of SARS-CoV-2 Infectivity and Antiviral Treatment Effects In Vitro. Viruses 2022, 14, 1301. [Google Scholar] [CrossRef] [PubMed]
- Courrol, L.C.; de Oliveira Silva, F.R.; Masilamani, V. SARS-CoV-2, hemoglobin and protoporphyrin IX: Interactions and perspectives. Photodiagnosis Photodyn. Ther. 2021, 34, 102324. [Google Scholar] [CrossRef]
- Chang, C.; Michalska, K.; Jedrzejczak, R.; Maltseva, N.; Endres, M.; Godzik, A.; Kim, Y.; Joachimiak, A. Crystal structure of RNA binding domain of nucleocapsid phosphoprotein from SARS coronavirus 2. Protein Data Bank 2020. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Mirdita, M.; Steinegger, M.; Soding, J. MMseqs2 desktop and local web server app for fast, interactive sequence searches. Bioinformatics 2019, 35, 2856–2858. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Šali, A.; Blundell, T.L. Comparative protein modeling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; States, D.J.; Swaminathan, S.; Karplus, M. CHARMM: A program for macromolec ular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]
- Dagistan, Y.; Kilinc, E.; Balci, C.N. Cervical sympathectomy modulates the neurogenic inflammatory neuropeptides following experimental subarachnoid hemorrhage in rats. Brain Res. 2019, 1722, 146366. [Google Scholar] [CrossRef] [PubMed]
- Lorente, L.; Martín, M.M.; Almeida, T.; Hernández, M.; Ramos, L.; Argueso, M.; Cáceres, J.J.; Solé-Violán, J.; Jiménez, A. Serum substance P levels are associated with severity and mortality in patients with severe traumatic brain injury. Crit. Care 2015, 19, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, A.N.; Philipp, M.T. Substance P and Antagonists of the Neurokinin-1 Receptor in Neuroinflammation Associated with Infectious and Neurodegenerative Diseases of the Central Nervous System. J. Neurol. Neuromedicine 2016, 1, 29–36. [Google Scholar] [CrossRef]
- Niemela, V.; Landtblom, A.M.; Nyholm, D.; Kneider, M.; Constantinescu, R.; Paucar, M.; Svenningsson, P.; Abujrais, S.; Burman, J.; Shevchenko, G.; et al. Proenkephalin Decreases in Cerebrospinal Fluid with Symptom Progression of Huntington’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2021, 36, 481–491. [Google Scholar] [CrossRef]
- Otsuka, K.; Niimi, A.; Matsumoto, H.; Ito, I.; Yamaguchi, M.; Matsuoka, H.; Jinnai, M.; Oguma, T.; Takeda, T.; Nakaji, H.; et al. Plasma substance P levels in patients with persistent cough. Respiration 2011, 82, 431–438. [Google Scholar] [CrossRef] [Green Version]
- Smieszek, S. Late Breaking Abstract—Increased Substance P levels in COVID-19 hospitalized patients. Eur. Respir. J. 2021, 58, OA4114. [Google Scholar] [CrossRef]
- Tyagi, A.; Daliri, E.B.-M.; Kwami Ofosu, F.; Yeon, S.-J.; Oh, D.-H. Food-Derived Opioid Peptides in Human Health: A Review. Int. J. Mol. Sci. 2020, 21, 8825. [Google Scholar] [CrossRef]
- Boaventura, P.; Macedo, S.; Ribeiro, F.; Jaconiano, S.; Soares, P. Post-COVID-19 Condition: Where Are We Now? Life 2022, 12, 517. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NTD Ligands | FL Ligands | Biological Function |
|---|---|---|
| DHF | DHF | Metabolism |
| THF | THF | Metabolism |
| AICAR (AMPK agonist) | Lauroyl coA | Metabolism |
| Hemin | Metabolism | |
| Naproxen Acetamine Indomethacin | Prostaglandin E2 and F2 and Other eicosanoids | COX pathway/NSAID COX pathway/NSAID COX pathway/NSAID |
| D Ala2-Leu5-Enkephalin YAGFL-OH (DADLE) | Neuropeptide involved in pain reduction, agonist of the μ− opioid receptor | |
| Substance P (1-7) RPKPQQF-OH Cholecystokinin DYMGWMDF (CCK8) | Neuropeptide mediator of inflammation, pain, and vasodilation, agonist of neuro- -kinin-1 receptor. Intestinal hormone peptide binding to a receptor on nerve fibers of the vagus nerve |
| Ligand | Protein | -CDocker Interaction Energy Kcal/mol | Critical Residues | Mutations SPR (% Inhibition/NTD wt) | KD | |
|---|---|---|---|---|---|---|
| DHF |  | NTD monomer | 58.0 | G44, L45, N77, H145, R149, N150, N153 | H145 (96%), R149 (83%) | 1 µM |
| THF |  | NTD monomer | 61.7 | |||
| Naproxen |  | NTD monomer | 38.0 | W52, I146, R149 (site 1) [17] A50, R88, R92, R93 (site 2) | R88 (55%), R149 (44%) | 4.4 ± 1.4 µM [17] |
| Acetamine |  | FL | 39.7 | F286, I304, S318, T334, G335, I337 | ||
| DHF | FL | 57.9 | R189, K233, K237, N239 | |||
| Enkephalin |  | FL | 64.4 | A155, K261 a, F274, F286, G287, Y298, I304, S318, Y333, G335, A336 | ||
| Lauroyl coA | FL | 79.5 | S187, S188, R189, K261 a, R262 a, | |||
| Substance P (1-7) |  | FL | 82.4 | S188, R189, R259 a,T263 a, A264 a, R277, F286, L291, G295, Y298, Y333, T334, H356 | ||
| Cholecystokinin |  | FL | 82.0 | A55, R107, Y109, V158, A264 a, V270, L291, W301, P302, H356 | ||
| Hemin |  | FL | 47 | A264 a, V270,F274, L291, I304, A305, I337 | ||
| ZnTPPS |  | FL | 0.4 ± 0.1 µM |
| Ligand | Ligand Concentration (µM) | Size (nm) Intensity | Size (nm) Volume | N NTD/FL |
|---|---|---|---|---|
| none | 5.3 ± 0.3 (80%) | Does not fit | NTD (60 µM) | |
| none | 13.2 ± 1.0 (85%) | 12.0 ± 1.0 (96%) | FL 2 µM | |
| Naproxen | 2 6 | 12.2 ± 0.5 (40%) 8.7 ± 0.3 (60%) | 11.3 ± 0.4 (99%) 8.9 ± 0.3 (96%) | FL 2 µM |
| Indomethacin | 2 6 | 11.0 ± 0.4 (30%) 6.7 ± 0.3 (60%) | 10.3 ± 0.5 (100%) 6.3 ± 0.4 (96%) | FL 2 µM |
| Enkephalin | 2 6 | 12.6 ± 0.6 (31%) 8.7 ± 0.2 (40%) | 11.3 ± 0.6 (96%) 8.0 ± 0.3 (100%) | FL 2 µM |
| Substance P | 2 6 | 10.3 ± 0.6 (50%) 8.0 ± 1.0 (76%) | 9.5 ± 0.6 (98%) 7.2 ± 1.1 (99%) | FL 2 µM |
| ZnTPPS | 2 | 7.6 ± 0.6 (25%) | 7.3 ± 0.6 (99%) | FL 2 µM |
| RNA (TAR-polyA) | 4 | 21.5 ± 2.0 (75%) | 15.1 ± 1.1 (98%) | FL 4 µM |
| 48m-DNA | 4 | 25.3 ± 3.1 (56%) | 15.7 ± 1.2 (96%) | FL 4 µM |
| RNA + subP | 4 | 16.1 ± 0.6 (75%) | 11.3 ± 0.6 (97%) | FL 4 µM |
| DNA + subP | 4 | 13.8 ± 0.6 (70%) | 10.1 ± 0.6 (91%) | FL 4 µM |
| DNA + enkephalin | 4 | 10.5 ± 1.2 (89%) | 9.0 ± 1.7 (99%) | FL 4 µM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henri, J.; Minder, L.; Mohanasundaram, K.; Dilly, S.; Goupil-Lamy, A.; Di Primo, C.; Slama Schwok, A. Neuropeptides, New Ligands of SARS-CoV-2 Nucleoprotein, a Potential Link between Replication, Inflammation and Neurotransmission. Molecules 2022, 27, 8094. https://doi.org/10.3390/molecules27228094
Henri J, Minder L, Mohanasundaram K, Dilly S, Goupil-Lamy A, Di Primo C, Slama Schwok A. Neuropeptides, New Ligands of SARS-CoV-2 Nucleoprotein, a Potential Link between Replication, Inflammation and Neurotransmission. Molecules. 2022; 27(22):8094. https://doi.org/10.3390/molecules27228094
Chicago/Turabian StyleHenri, Julien, Laetitia Minder, Kevin Mohanasundaram, Sébastien Dilly, Anne Goupil-Lamy, Carmelo Di Primo, and Anny Slama Schwok. 2022. "Neuropeptides, New Ligands of SARS-CoV-2 Nucleoprotein, a Potential Link between Replication, Inflammation and Neurotransmission" Molecules 27, no. 22: 8094. https://doi.org/10.3390/molecules27228094