Article Text

Abstract
Introduction The diaphragm is the main muscle of inspiration, and its dysfunction contributes to adverse clinical outcomes in critically ill patients. We recently reported the infiltration of SARS-CoV-2, and the development of fibrosis, in the diaphragm of critically ill patients with COVID-19. In the current study, we aimed to characterise myofiber structure in the diaphragm of critically ill patients with COVID-19.
Methods Diaphragm muscle specimens were collected during autopsy from patients who died of COVID-19 in three academic medical centres in the Netherlands in April and May 2020 (n=27). We studied diaphragm myofiber gene expression and structure and compared the findings obtained to those of deceased critically ill patients without COVID-19 (n=10).
Results Myofibers of critically ill patients with COVID-19 showed on average larger cross-sectional area (slow-twitch myofibers: 2441±229 vs 1571±309 µm2; fast-twitch myofibers: 1966±209 vs 1225±222 µm2). Four critically ill patients with COVID-19 showed extremely large myofibers, which were splitting and contained many centralised nuclei. RNA-sequencing data revealed differentially expressed genes involved in muscle regeneration.
Conclusion Diaphragm of critically ill patients with COVID-19 has distinct myopathic features compared with critically ill patients without COVID-19, which may contribute to the ongoing dyspnoea and fatigue in the patients surviving COVID-19 infection.
- COVID-19
- respiratory muscles
Data availability statement
Data are available on reasonable request. All data relevant to the study are included in the article or uploaded as online supplemental information.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is the key question?
RNA-sequencing results from the diaphragm of deceased patients with COVID-19 suggest the activation of gene pathways involved in myofiber regeneration.
What is the bottom line?
In line with the RNA-sequencing results, the results of structural analyses of the diaphragm specimens show enlarged and splitting myofibers with central nuclei.
What read on?
The diaphragm of deceased patients with COVID-19 displays distinct pathological changes, which may contribute to the ongoing dyspnoea and fatigue in patients who survive COVID-19.
Introduction
In critically ill patients, diaphragm weakness may develop and contribute to adverse clinical outcomes including ventilator weaning failure, prolonged hospitalisation and mortality.1 Diaphragm weakness in these patients is characterised by pathological features in myofibers, including reduced cross-sectional area (CSA), that is, atrophy.2–4 Recently, we reported evidence for infiltration of SARS-CoV-2 in diaphragm myofibers.5 We also demonstrated increased expression of genes involved in fibrosis and histological evidence for the development of fibrosis in the diaphragm of critically ill patients with COVID-19 compared with the diaphragm of critically ill patients without COVID-19.5 In the current study, we characterised myofiber structure in the diaphragm of critically ill patients with COVID-19. Myofiber structure is an important determinant of diaphragm contractility. Consequently, impaired myofiber structure might contribute to diaphragm muscle weakness, and, thereby, impact weaning outcome and contribute to the persistent dyspnoea and fatigue reported in post-COVID-19 patients.6 Thus, we aimed to study the structure of diaphragm myofibers in critically ill patients with COVID-19 and compared the findings to those of patients without COVID-19.
Methods
Patients
Diaphragm specimens of critically ill patients with COVID-19 (COVID-19-intensive care unit (ICU), n=27) and of critically ill patients without COVID-19 (control-ICU, n=10) were collected at autopsy. Diagnosis of COVID-19 was based on SARS-CoV-2 detection by real-time quantitative PCR, in nasopharyngeal swabs. Samples were obtained from the middle of the left costal diaphragm (figure 1A), 16–72 hours postmortem. Patients were recruited in three medical centres in the Netherlands. Clinical data were collected retrospectively.
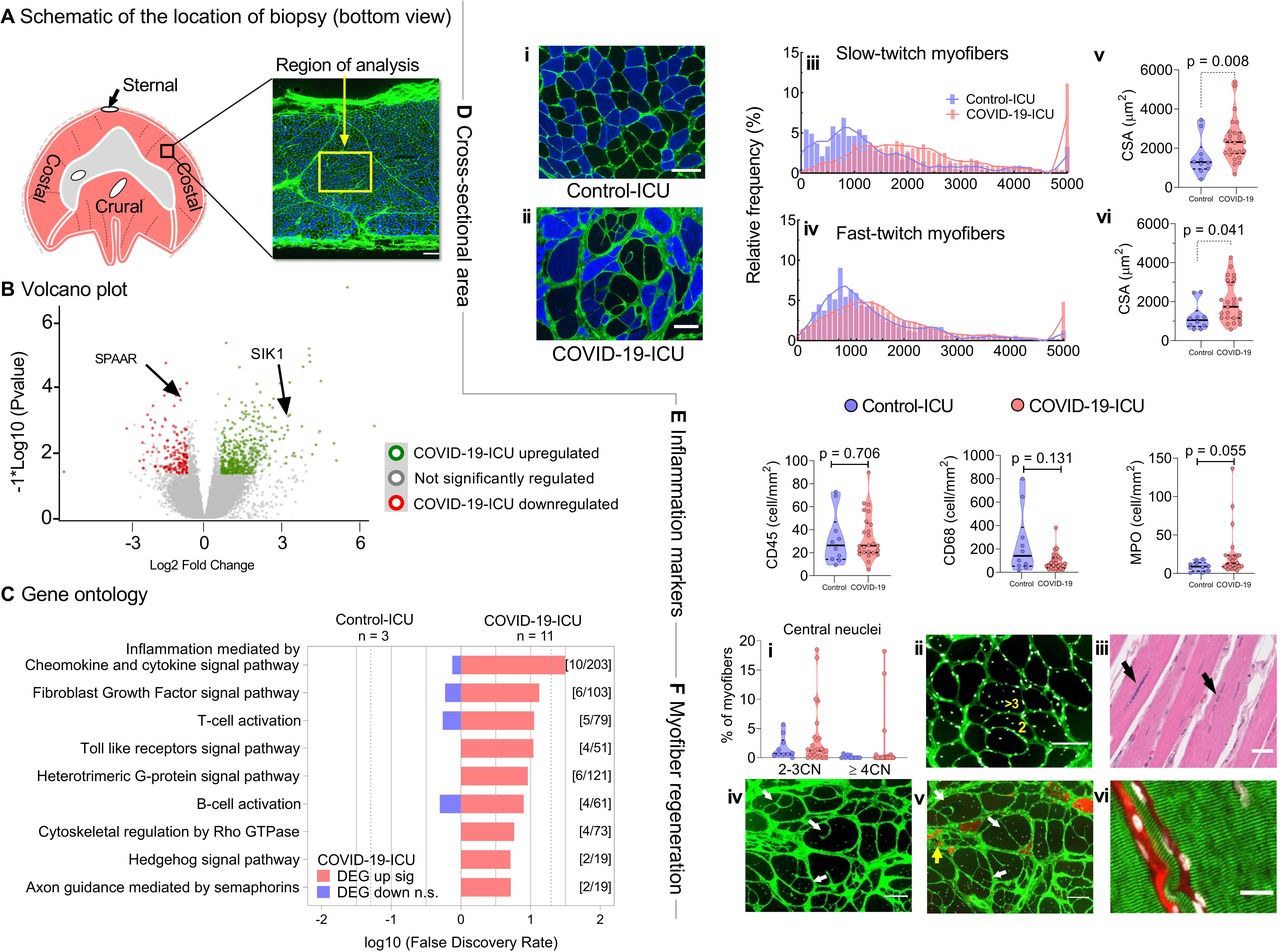
{kind=link}
(A)Schematic of the location of the biopsy obtained (balck and yellow squares). (B) Volcano plot showing the 315 upregulated and 281 downregulated genes (SPAAR: small regulatory polypeptide of amino acid response; SIK1: salt inducible kinase) and (C) Gene ontology biological process enrichment analysis (PANTHER) showing upregulated/downregulated processes in the diaphragm of COVID-19-ICU (n=11) patients compared with Control-ICU (n=3). (DEG: differentially expressed gene). (D) Cross-sectional area (CSA) in COVID-19-ICU (n=25) and Control-ICU patients (n=10). Representative images of one patient from Control-ICU (i) and COVID-19-ICU (ii), respectively. Cross-sections stained for fast myosin heavy chain isoforms (blue) to identify fast-twitch myofibers; slow-twitch myofibers are black; in green WGA to demarcate myofibers; bar=100 µm. Note the extremely large myofibers in the COVID-19-ICU patient (patient #28 is shown). Distribution of CSA of slow-twitch (iii) and fast-twitch (iv) myofibers of COVID-19-ICU patients (red) skewed towards larger CSA compared with Control-ICU patients (purple). The CSA of both the slow-twitch and fast-twitch myofibers were significantly larger in COVID-19-ICU patients compared with Control-ICU patients (v, vi). Data are visualised using histogram and violin plot. Each dot represents mean of the CSA from an individual patient. P values were obtained from linear mixed model analysis. (E) Quantification of CD68+macrophages, CD45+ leucocytes, and MPO+ neutrophils in COVID-19-ICU and Control-ICU patients. No differences were observed between groups. The data are visualised using violin plot with each dot representing the value of an individual patient. P values were obtained from Mann-Whitney U test. (F) Signs of myofiber regeneration in COVID-19-ICU patients. (i) Quantification of the number of central nuclei in cross-sectioned myofibers. Note that four COVID-19-ICU patients (#2;3;28;31) had an increased number of myofibers with 2–3 central nuclei; three of these four patients had myofibers with more than three central nuclei (up to 18 per myofiber). (ii) Cross-section of patient #31 stained with WGA and DAPI to localise the myonuclei; yellow numbers indicate representative myofibers with multiple central nuclei; bar=100 µm. (iii) H&E staining (formalin fixed sample) of a longitudinal section of patient #3 showing multiple ‘trains’ of central nuclei (arrows); bar=100 µm. (iv, v). Two serial cross-sections (separated by 100 µm) of patient #28, showing splitting of myofibers; arrows indicate the same myofibers with splitting events; in green WGA to demarcate myofibers. Also, note the large variation of myofiber CSA in this patient. In (v), α-MHC3 antibody staining (red) visualises myofibers expressing an embryonic isoform of myosin heavy chain. Note that the split fibers are adult, non-regenerating myofibers (white arrows) and that the tiny myofibers surrounded by fibrosis are regenerating (yellow arrows); bar=100 µm. (vi) Longitudinal section of patient #28 stained with WGA to demarcate the membrane (red) and DAPI to localise the myonuclei (white); the α-actinin antibody staining in green shows the Z-disc organisation. From left to the right, the diameters for the fibers are 40, 10 and 160 µm, respectively; bar=20 µm. ICU, intensive care unit; WGA, wheat germ agglutinin; DAPI, 4',6-diamidino-2-phenylindole; MHC, myosin heavy chain.
Transcriptome sequencing (RNA-seq)
RNA and library preparation integrity were confirmed with a LabChip Gx Touch 24 (Perkin Elmer). The function of genes was referred to the ‘National Center for Biotechnology Information (NCBI)’s Gene’ and ‘GeneCards’ databases. Because only samples of sufficient integrity were further processed for RNA sequencing, 3 control-ICU patients and 11 COVID-19-ICU patients were analysed.
Histology and immunohistochemistry to study myofiber structure
Serial cryosections (8 µm thick) were cut from the frozen biopsies (perpendicular to diaphragm fiber direction) and stained to study (1) myofiber CSA using wheat germ agglutinin staining and myofiber type using myosin heavy chain antibodies, (2) myofiber nuclei using 4',6-diamidino-2-phenylindole staining and (3) inflammatory infiltrates using CD45, CD68 and myeloperoxidase (MPO) antibodies. To study the striation pattern, longitudinal sections were cut and studied by α-acting staining (to visualise the z-disc of sarcomeres) and by H&E staining (to visualise the overall structure of the myofibers in formalin-fixed tissue). Myofiber CSA was obtained from ±100 myofibers per type per biopsy (analysis of >100 myofibers did not affect the coefficient of variation (CV) of the CSA). Inflammatory cell counts (leucocytes, macrophages and neutrophils) were obtained from three random selected viewing fields per section per biopsy (0.5 mm2 for fibrosis and 1 mm2 for inflammatory cells, respectively). Central nucleus was defined as the presence of one nucleus anywhere in the sarcoplasm and at least 5 µm inside the sarcolemma.
Statistical analysis
Data were analysed with IBM SPSS Statistics V.22 (IBM) and visualised with GraphPad Prism V.7.0 (GraphPad Software, USA). For repeated measurements (ie, CSA and inflammatory cells), CV was calculated to assess the variations in individual patients. Linear mixed model was applied with patients as the random factor and group as fixed factor to compare the difference between two groups of patients. Assumption of normality was tested by the Shapiro-Wilk normality test. Log transformation was performed if necessary. For non-repeated measurements, the Mann-Whitney U test or Student’s t-test was applied for quantitative variables. A χ2 or Fisher’s exact test was used to compare qualitative variables. Data were expressed as mean (±SEM), median (IQR) or frequencies and percentage, as indicated. We used a two-sided significance level of 5% for all analyses.
Patient and public involvement
Patients or the public were not involved in the design, conduct, reporting and dissemination plans of our research.
Results
Between both groups, there were no significant differences in the number of days on mechanical ventilation, the length of ICU stay, and the administration of neuromuscular blocking agents and steroids. None of the patients had pre-existing neuromuscular disease or chronic cardiopulmonary disease. All patients with COVID-19 were admitted to the ICU because of COVID-19 infection and required invasive mechanical ventilation. More clinical characteristics are in table 1 and online supplemental table 1.
Supplemental material
Summary of the demographic and clinical characteristics
First, we investigated gene expression in diaphragm tissue from eleven COVID-19-ICU patients and three control-ICU patients by RNA sequencing (only samples with RNA of sufficient integrity were used). A total of 596 (315 upregulated; 281 downregulated) of 27 647 detected genes were differentially expressed (fold-change >±1.5; p<0.05; figure 1B). Among the five most upregulated and downregulated genes were the small regulatory polypeptide of amino acid response (SPAAR, downregulated) and the salt inducible kinase (SIK1, upregulated), both expressed in the direction of promoting muscle regeneration.7 8 However, genes that are known to be involved in critical illness-associated muscle atrophy, such as the ubiquitin-proteasome and autophagy-lysosome pathways,9 were not differentially expressed between groups. Gene ontology analysis by protein analysis through evolutionary relationships (PANTHER) revealed upregulation of the Fibroblast Growth Factor pathway (as reported before,5 and of several pathways involved in inflammation (figure 1C).
Prompted by the gene expression analyses, diaphragm sections were stained to study myofiber CSA, inflammatory infiltrates, localisation of myonuclei and the striation pattern of the myofibers. The distribution of myofiber CSA in COVID-19-ICU patients shifted towards larger myofibers compared with control-ICU patients (figure 1D). The average myofiber CSA was larger in COVID-19-ICU compared with control-ICU patients (slow-twitch: 2441±229 vs 1571±309 µm2, respectively, p=0.008; figure 1Diii/v; fast-twitch: 1966±209 vs . 1225±222 µm2, respectively, p=0.041; figure 1Div/vi; note that analysis of minimal Feret diameter rendered comparable results. Data are shown in the online supplement, online supplemental figure 1). In COVID-19-ICU patients, the maximum CV was 140% for slow-twitch myofiber and 132% for fast-twitch myofibers. The average CV, however, was comparable between COVID-19-ICU and control-ICU patients (slow-twitch: 49±21% vs 51±12%, respectively, p=0.46; fast-twitch: 50±23% vs 48±12%, respectively, p=0.653). No significant differences were observed between groups for the number of lymphocytes (CD45+), macrophages (CD68+) and neutrophilic granulocytes (MPO+) (figure 1E).
In four COVID-19-ICU patients (mechanical ventilation duration 5–49 day), we observed diaphragm myofibers with central nuclei (figure 1Fi–iii), indicative of myofiber regeneration. Longitudinal sections showed that, occasionally, these central nuclei were organised in ‘trains’ (figure 1Fiii), as frequently observed in muscular dystrophies. These four patients also displayed extremely large myofibers (CSA >10 000 µm2) and splitting of these large myofibers (figure 1Fiv-v), consistent with diaphragm myopathy and regeneration, which was not present in control-ICU patients. Significant higher variation in myofiber CSA was observed in these four patients compared with control-ICU patients (CV: slow-twitch: 81%±40%; fast-twitch: 90%±32%). Importantly, longitudinal sections of the diaphragms showed clear striation patterns across these extremely large myofibers, indicating increased myofibrillar content rather than the accumulation of organelles such as mitochondria or of degradation products resulting from postmortem autolysis (figure 1Fvi).
Discussion
The present study reveals distinct pathological changes in the diaphragm myofibers of critically ill patients with COVID-19. A striking observation was the larger CSA of myofibers, which was independent of the duration of mechanical ventilation (table 1). It is likely that the increased variation of myofiber CSA reflects myofiber regeneration and splitting, as supported by the central nuclei in myofibers and the RNA sequencing data showing DEGs involved in myofiber regeneration. Our previous work on these diaphragm samples showed increased fibrosis,5 which suggests that the myofiber regeneration was—at least partly—pathological. We speculate that these pathological changes may impact the contractility of the diaphragm in patients with COVID-19, and contribute to the persisting sensation of dyspnoea and fatigue that a subset of patients experience.6 Myofiber regeneration and splitting occurs in response to myofiber injury. The aetiology of myofiber injury in COVID-19-ICU patients is unclear. It is tempting to speculate that the pathological changes might be a direct effect of the virus, as we have recently shown that SARS-CoV-2 particles may infiltrate diaphragm myofibers,5 but it remains to be established whether this affects gene expression. Alternatively, it may be explained by a high respiratory drive in mechanically ventilated patients with COVID-19,10 as high respiratory loading is known to cause diaphragm myofiber injury.11 However, this was not specifically studied in our patients.
This study has some limitations. First, without non-critically ill patients as a control group, it is unclear whether myofibers of COVID-19-ICU patients are less atrophic than those of control-ICU patients or are more hypertrophic compared with non-critically ill patients. Furthermore, as the current study was cross-sectional of nature, we cannot exclude that baseline variations affected the study outcomes. Importantly, none of the patients had a history of neuromuscular diseases, which reduces the possibility of pre-existing diaphragm pathology. Second, the number of patients recruited was small, particularly for the control-ICU patients (n=10). Consequently, although groups were comparable regarding the duration of mechanical ventilation and the use of corticosteroids, both risk factors for diaphragm pathology,1 they were not for body mass index. Additionally, although proposed as a risk factor for diaphragm pathology,1 sepsis was present in only three of the control-ICU patients (and in none of the COVID-19-ICU patients), thus, it is unlikely that this contributed to the diaphragm pathology observed in the current study. Furthermore, some of the pathological changes were observed in a subset of COVID-19-ICU patients. However, we have published several studies2–4 that included critically ill patients undergoing elective surgery and not in a single case did we observe the pathological changes (increased variation of myofiber CSA, extremely large myofiber CSA, central nuclei, myofiber splitting) reported here. Nevertheless, caution is warranted when drawing conclusions based on these findings. They should be considered as hypothesis generating, and future studies should focus on the underlying mechanisms and risk factors for these myopathic features in the diaphragm. Third, in both groups the samples were collected 16–72 hours postmortem, and tissue degradation may have occurred. However, the longitudinal sections revealed preserved striation pattern (figure 1Fvi), indicating that gross myofiber morphology was intact. Finally, only 3 of the 10 control-ICU patients had a viral infection, which warrants caution when comparing direct effects of SARS-CoV-2 with those of other viruses. Furthermore, RNA-sequencing analyses were performed in a subset of patients, therefore, caution is warranted when relating the findings from the RNA-sequencing analyses to those from the histological analyses.
Conclusion
In summary, the diaphragm of deceased COVID-19 patients shows distinct myopathic changes. Although it is unknown whether these changes in the diaphragm persist in COVID-19 survivors, it is tempting to speculate that they contribute to the ongoing dyspnoea and fatigue in those patients.
Data availability statement
Data are available on reasonable request. All data relevant to the study are included in the article or uploaded as online supplemental information.
Ethics statements
Patient consent for publication
Ethics approval
The study was approved by the medical ethical committee at Amsterdam UMC (#2020-167).
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors CACO, LH, ZS and HvdH contributed to conception and design. ZS, SJPB, SC, YO, PE, DIB, MB, MvdL, MvdB and RAB contributed to acquisition and analysis of data. ZS, SJPB, MvdB, MvdL, RAB, CACO and LH contributed to interpretation of data. ZS, CACO, LH and HvdH contributed to drafting the manuscript. All authors reviewed, edited and approved the final manuscript.
Funding Research reported in this work was supported by a grant from the National Institutes of Health-Heart Lung and Blood Institute R01HL121500 (CACO).
Competing interests LH reports grants and personal fees from Liberate medical, personal fees from Getinge, personal fees from Fisher and Paykel, outside the submitted work.
Provenance and peer review Not commissioned; internally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.