
Comparison of Physical and Biochemical Characterizations of SARS-CoV-2 Inactivated by Different Treatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus Culturing
2.2. Virus Inactivation
2.3. Validation of the Inactivation
2.4. Virus Purification
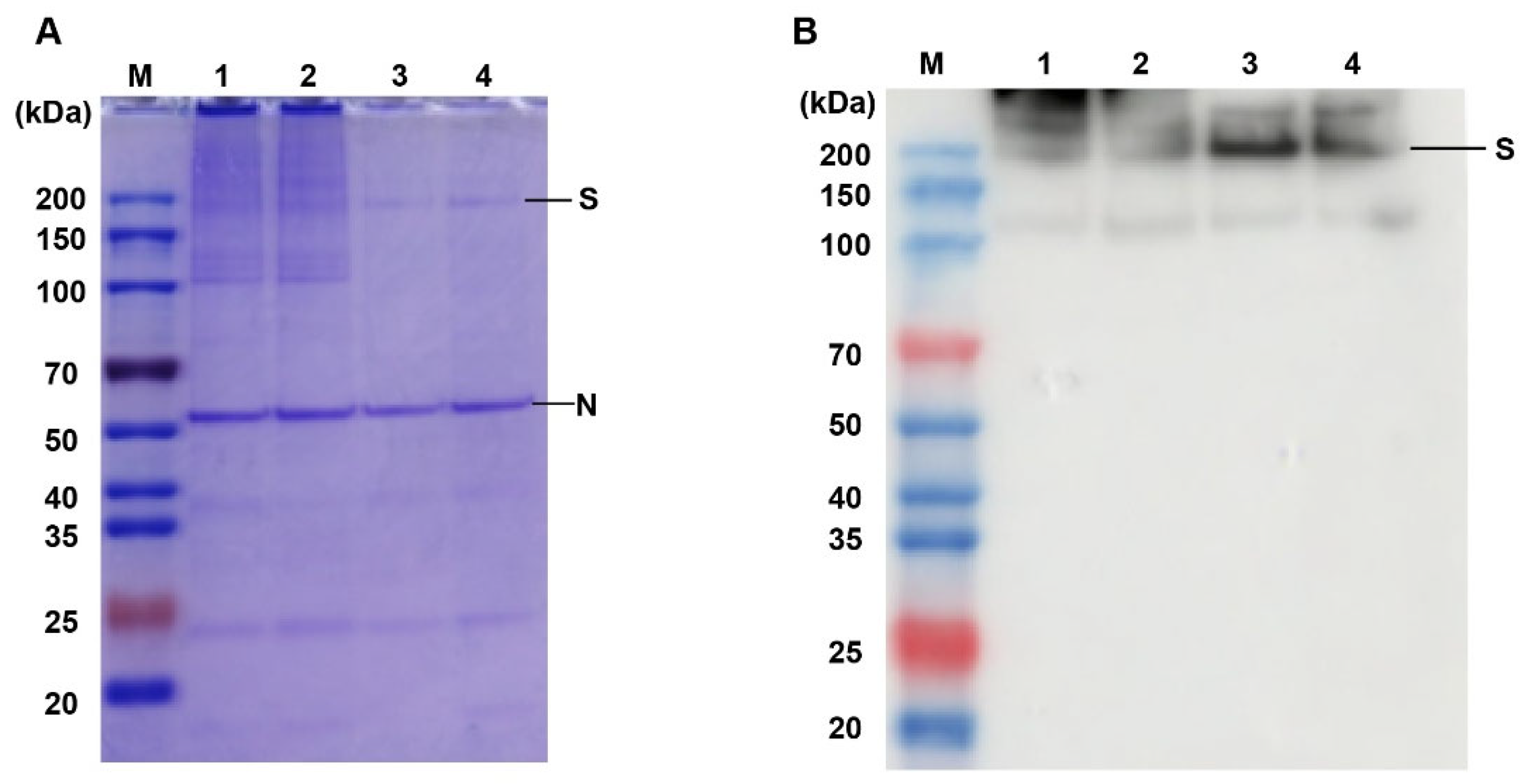
2.5. SDS-PAGE
2.6. Western Blot
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Quantitative Determination of Protein
2.9. EM Sample Preparation
2.10. Dynamic Light Scattering (DLS)
2.11. Circular Dichroism (CD) Spectroscopy
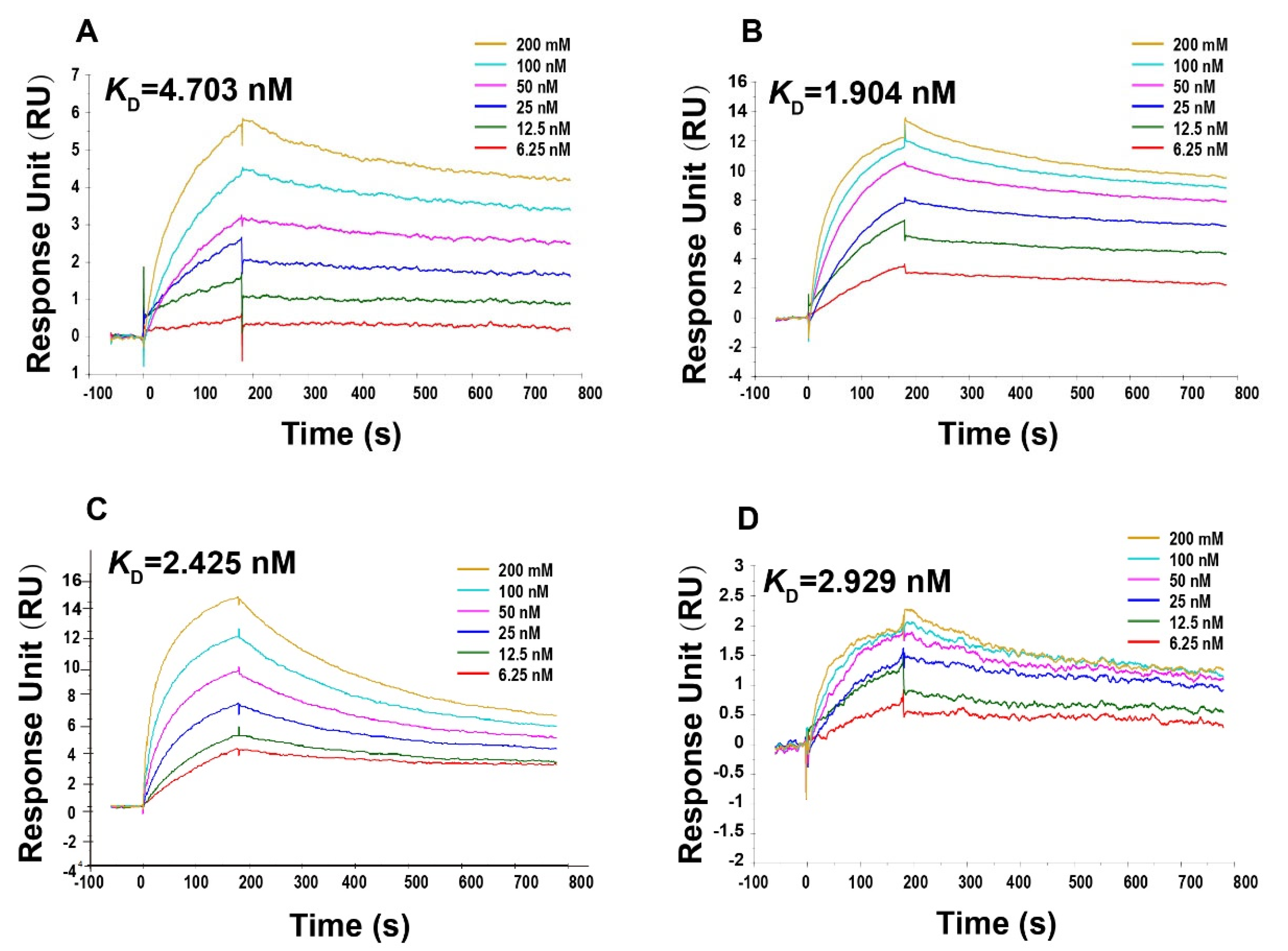
2.12. Surface Plasmon Resonance (SPR)
3. Results
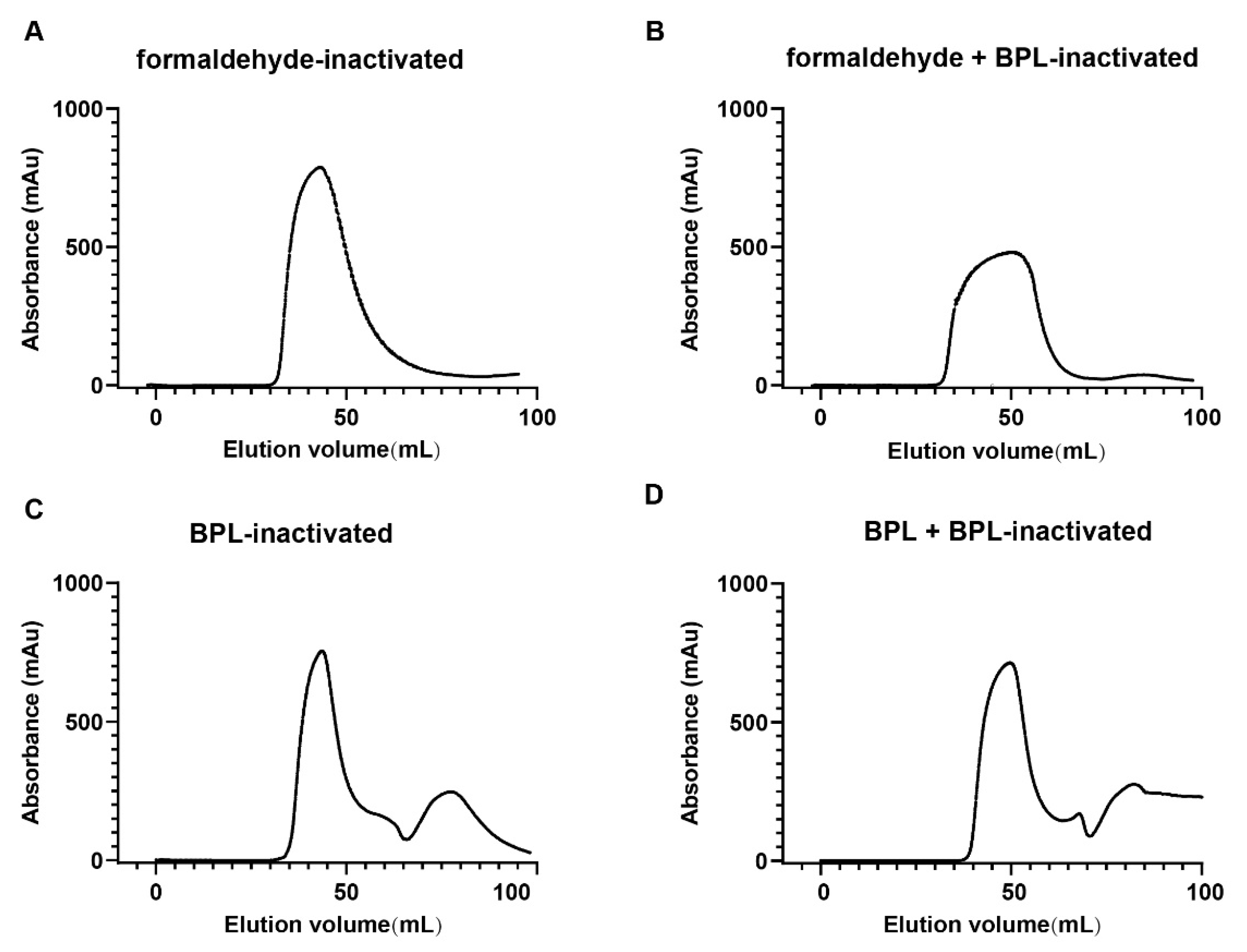
3.1. Effects of Different Inactivation Methods on the Homogeneity of Virus Particles
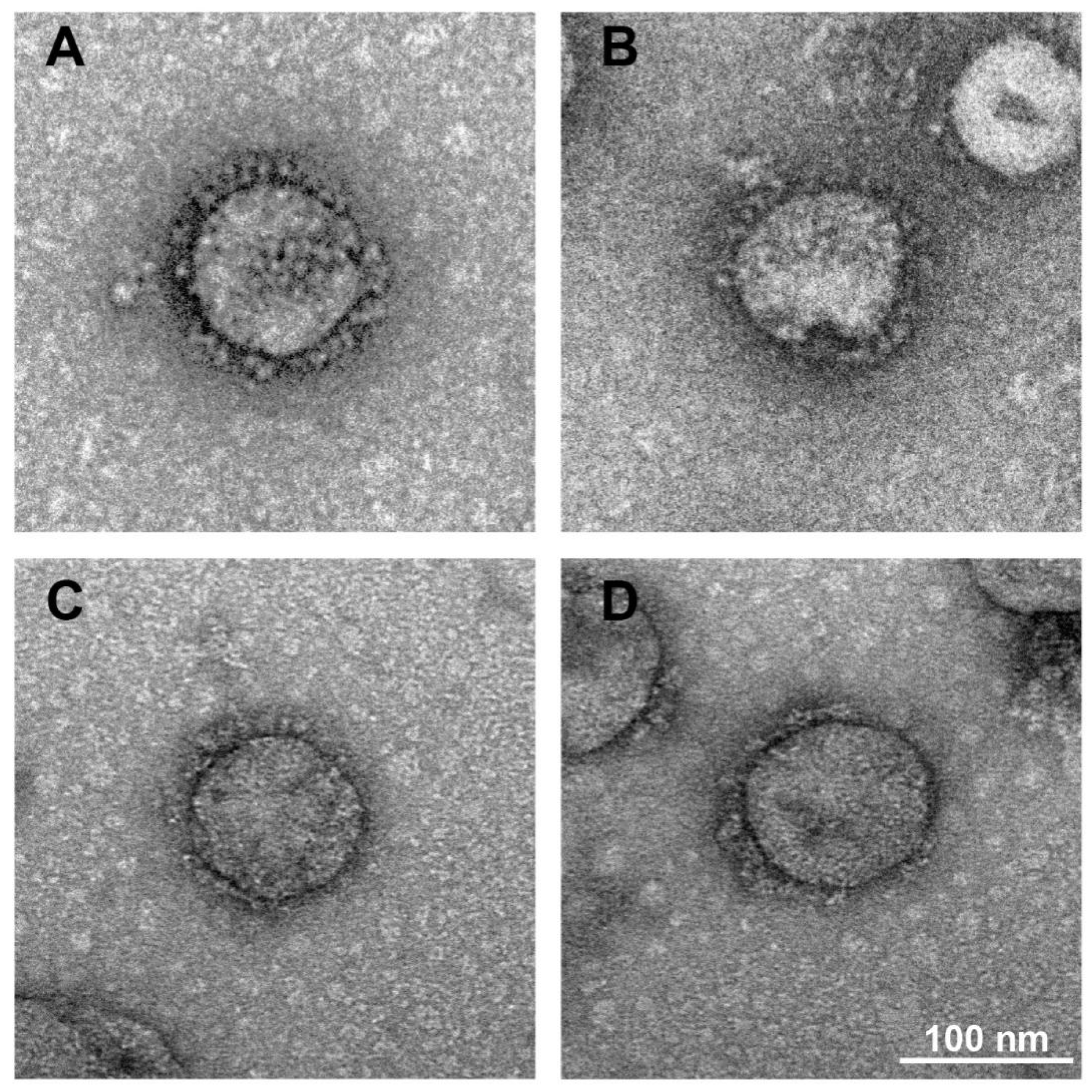
3.2. Formaldehyde and BPL Treatment Have Less of an Impact on the Morphology of SARS-CoV-2
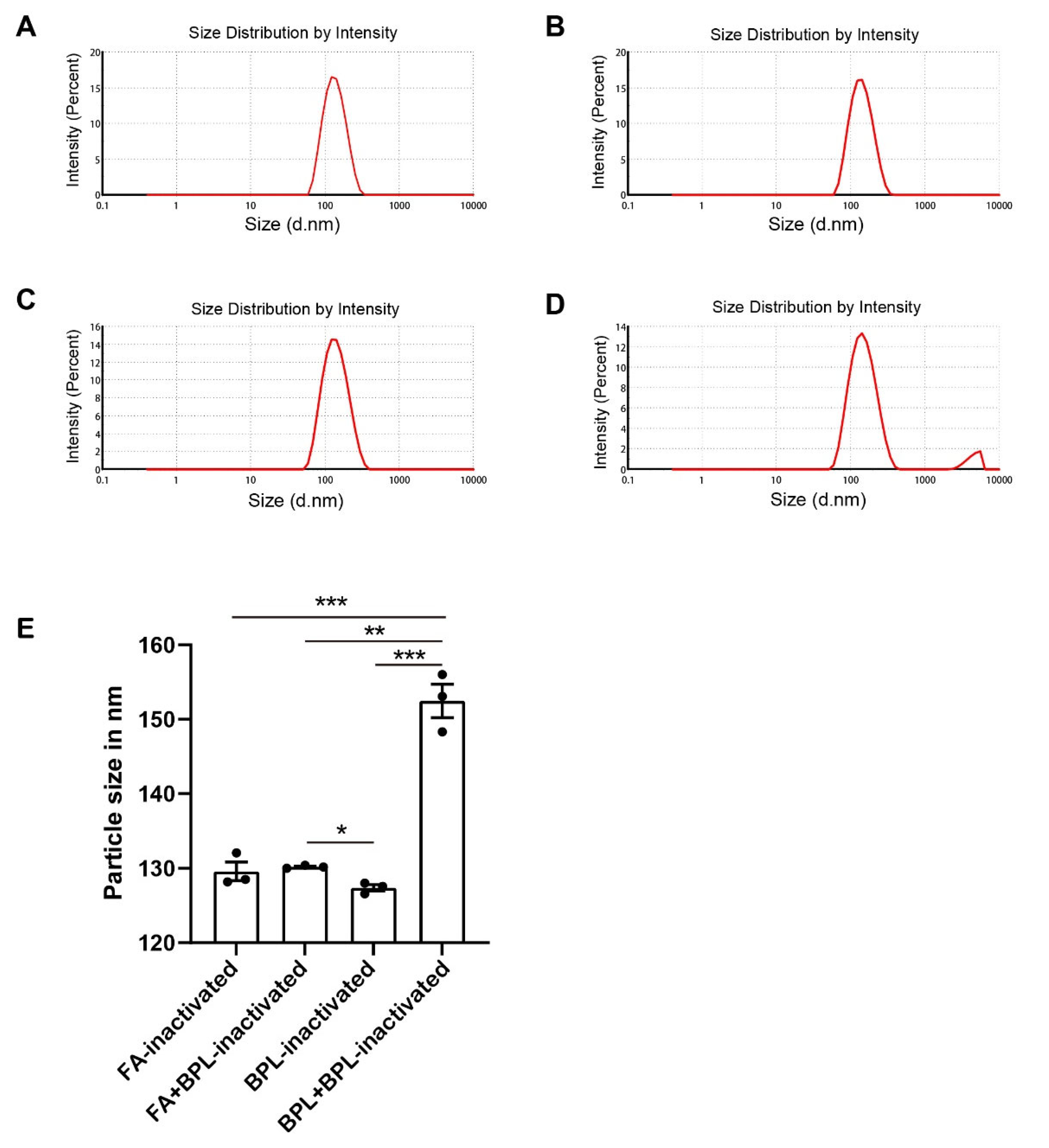
3.3. Long Time Treatment of BPL Causes the Aggregation of SARS-CoV-2
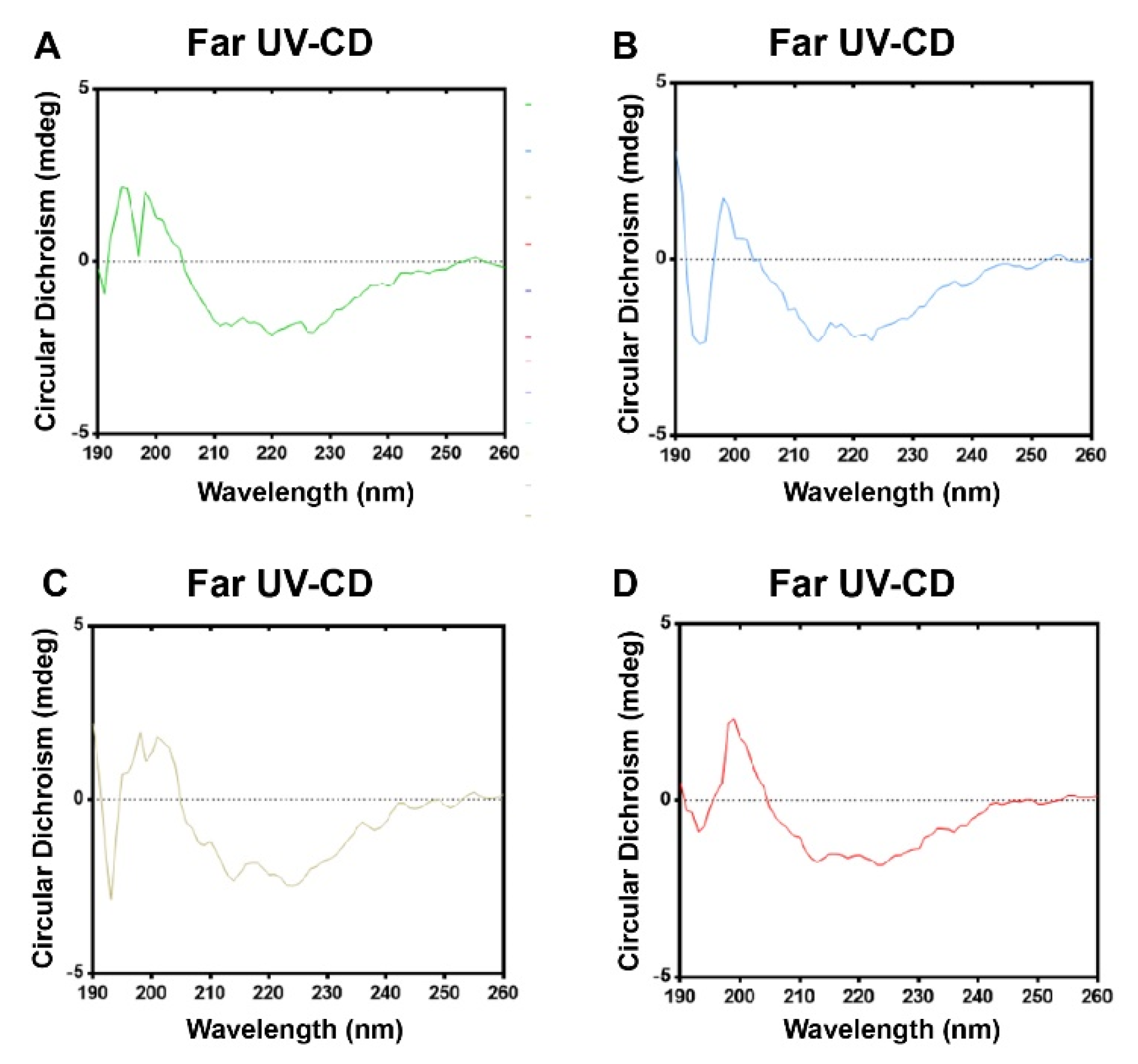
3.4. Formaldehyde and BPL Treatment Have Less of an Impact on the Secondary Structure of SARS-CoV-2
3.5. Effects of Different Inactivation Methods on the Content of S Antigen
3.6. Formaldehyde and BPL Treatment Does Not Affect the Ability of the SARS-CoV-2 to Bind to the S Protein Antibody
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Acosta-Coley, I.; Cervantes-Ceballos, L.; Tejeda-Benítez, L.; Sierra-Márquez, L.; Cabarcas-Montalvo, M.; García-Espiñeira, M.; Coronell-Rodríguez, W.; Arroyo-Salgado, B. Vaccines platforms and COVID-19: What you need to know. Trop. Dis. Travel Med. Vaccines 2022, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Agrati, C.; Carsetti, R.; Bordoni, V.; Sacchi, A.; Quintarelli, C.; Locatelli, F.; Ippolito, G.; Capobianchi, M.R. The Immune Response as a Double-Edged Sword: The Lesson Learnt during the COVID-19 Pandemic. Immunology 2022. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1111/imm.13564 (accessed on 16 August 2022). [CrossRef] [PubMed]
- Marino, A.; Munafò, A.; Augello, E.; Bellanca, C.M.; Bonomo, C.; Ceccarelli, M.; Musso, N.; Cantarella, G.; Cacopardo, B.; Bernardini, R. Sarilumab Administration in COVID-19 Patients: Literature Review and Considerations. Infect. Dis. Rep. 2022, 14, 360–371. [Google Scholar] [CrossRef]
- Marino, A.; Pampaloni, A.; Scuderi, D.; Cosentino, F.; Moscatt, V.; Ceccarelli, M.; Gussio, M.; Celesia, B.M.; Bruno, R.; Borraccino, S.; et al. High-flow nasal cannula oxygenation and tocilizumab administration in patients critically ill with COVID-19: A report of three cases and a literature review. World Acad. Sci. J. 2020, 2, 23. [Google Scholar] [CrossRef]
- Loveday, E.K.; Hain, K.S.; Kochetkova, I.; Hedges, J.F.; Robison, A.; Snyder, D.T.; Brumfield, S.K.; Young, M.J.; Jutila, M.A.; Chang, C.B.; et al. Effect of Inactivation Methods on SARS-CoV-2 Virion Protein and Structure. Viruses 2021, 13, 562. [Google Scholar] [CrossRef]
- Patterson, E.I.; Prince, T.; Anderson, E.R.; Casas-Sanchez, A.; Smith, S.L.; Cansado-Utrilla, C.; Solomon, T.; Griffiths, M.J.; Acosta-Serrano, Á.; Turtle, L.; et al. Methods of Inactivation of SARS-CoV-2 for Downstream Biological Assays. J. Infect. Dis. 2020, 222, 1462–1467. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an Inactivated Vaccine Candidate, BBIBP-CorV, with Potent Protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef] [PubMed]
- Al Kaabi, N.; Zhang, Y.; Xia, S.; Yang, Y.; Al Qahtani, M.M.; Abdulrazzaq, N.; Al Nusair, M.; Hassany, M.; Jawad, J.S.; Abdalla, J.; et al. Effect of 2 Inactivated SARS-CoV-2 Vaccines on Symptomatic COVID-19 Infection in Adults: A Randomized Clinical Trial. JAMA 2021, 326, 35–45. [Google Scholar] [CrossRef]
- Gupta, D.; Parthasarathy, H.; Sah, V.; Tandel, D.; Vedagiri, D.; Reddy, S.; Harshan, K.H. Inactivation of SARS-CoV-2 by β-propiolactone causes aggregation of viral particles and loss of antigenic potential. Virus Res. 2021, 305, 198555. [Google Scholar] [CrossRef]
- Widera, M.; Westhaus, S.; Rabenau, H.F.; Hoehl, S.; Bojkova, D.; Cinatl, J.; Ciesek, S. Evaluation of stability and inactivation methods of SARS-CoV-2 in context of laboratory settings. Med. Microbiol. Immunol. 2021, 210, 235–244. [Google Scholar] [CrossRef]
- Awadasseid, A.; Wu, Y.; Tanaka, Y.; Zhang, W. Current advances in the development of SARS-CoV-2 vaccines. Int. J. Biol. Sci. 2021, 17, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Rodriguez, J.; Signorazzi, A.; Holtrop, M.; de Vries-Idema, J.; Huckriede, A. Inactivated or damaged? Comparing the effect of inactivation methods on influenza virions to optimize vaccine production. Vaccine 2019, 37, 1630–1637. [Google Scholar] [CrossRef]
- Auerswald, H.; Yann, S.; Dul, S.; In, S.; Dussart, P.; Martin, N.J.; Karlsson, E.A.; Garcia-Rivera, J.A. Assessment of inactivation procedures for SARS-CoV-2. J. Gen. Virol. 2021, 102, 001539. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.A.; Tauraso, N.M. Effect of formalin, beta-propiolactone, merthiolate, and ultraviolet light upon influenza virus infectivity chicken cell agglutination, hemagglutination, and antigenicity. Appl. Microbiol. 1970, 19, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Ye, X.; Ku, Z.; Kong, L.; Liu, Q.; Xu, C.; Cong, Y.; Huang, Z. Beta-Propiolactone Inactivation of Coxsackievirus A16 Induces Structural Alteration and Surface Modification of Viral Capsids. J. Virol. 2017, 91, e00038-17. [Google Scholar] [CrossRef]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Guo, J.; Wan, X.; Zhou, J.G.; Jin, W.P.; Lu, J.; Wang, W.H.; Yang, A.N.; Liu, D.X.; Shi, Z.L.; et al. Biochemical and antigenic characterization of the structural proteins and their post-translational modifications in purified SARS-CoV-2 virions of an inactivated vaccine candidate. Emerg. Microbes Infect. 2020, 9, 2653–2662. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xie, Z.; Long, R.; Fan, S.; Li, H.; He, Z.; Xu, K.; Liao, Y.; Wang, L.; Zhang, Y.; et al. A valid protective immune response elicited in rhesus macaques by an inactivated vaccine is capable of defending against SARS-CoV-2 infection. bioRxiv 2020, 2020.08.04.235747. [Google Scholar]
- Basso, C.R.; Malossi, C.D.; Haisi, A.; de Albuquerque Pedrosa, V.; Barbosa, A.N.; Grotto, R.T.; Araujo Junior, J.P. Fast and reliable detection of SARS-CoV-2 antibodies based on surface plasmon resonance. Anal. Methods Adv. Methods Appl. 2021, 13, 3297–3306. [Google Scholar] [CrossRef]
- He, Y.; Qi, J.; Xiao, L.; Shen, L.; Yu, W.; Hu, T. Purification and characterization of the receptor-binding domain of SARS-CoV-2 spike protein from Escherichia coli. Eng. Life Sci. 2021, 21, 453–460. [Google Scholar] [CrossRef]
- Liu, C.; Mendonça, L.; Yang, Y.; Gao, Y.; Shen, C.; Liu, J.; Ni, T.; Ju, B.; Liu, C.; Tang, X.; et al. The Architecture of Inactivated SARS-CoV-2 with Postfusion Spikes Revealed by Cryo-EM and Cryo-ET. Structure 2020, 28, 1218–1224.e4. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zheng, B.; Zhang, L.; Cui, L.; Su, X.; Zhang, Q.; Guo, Z.; Guo, Y.; Zhang, W.; Zhu, L.; et al. Serotype specific epitopes identified by neutralizing antibodies underpin immunogenic differences in Enterovirus B. Nat. Commun. 2020, 11, 4419. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738.e13. [Google Scholar] [CrossRef]
- Xia, S.; Duan, K.; Zhang, Y.; Zhao, D.; Zhang, H.; Xie, Z.; Li, X.; Peng, C.; Zhang, Y.; Zhang, W.; et al. Effect of an Inactivated Vaccine Against SARS-CoV-2 on Safety and Immunogenicity Outcomes: Interim Analysis of 2 Randomized Clinical Trials. JAMA 2020, 324, 951–960. [Google Scholar] [CrossRef]
- Abdoli, A.; Aalizadeh, R.; Aminianfar, H.; Kianmehr, Z.; Teimoori, A.; Azimi, E.; Emamipour, N.; Eghtedardoost, M.; Siavashi, V.; Jamshidi, H.; et al. Safety and potency of BIV1-CovIran inactivated vaccine candidate for SARS-CoV-2: A preclinical study. Rev. Med. Virol. 2022, 32, e2305. [Google Scholar] [CrossRef] [PubMed]
- Kozlovskaya, L.I.; Piniaeva, A.N.; Ignatyev, G.M.; Gordeychuk, I.V.; Volok, V.P.; Rogova, Y.V.; Shishova, A.A.; Kovpak, A.A.; Ivin, Y.Y.; Antonova, L.P.; et al. Long-term humoral immunogenicity, safety and protective efficacy of inactivated vaccine against COVID-19 (CoviVac) in preclinical studies. Emerg. Microbes Infect. 2021, 10, 1790–1806. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Particle Size (nm) | PDI (Polydispersity Index) |
|---|---|---|
| Formaldehyde-inactivated | 129.60 ± 1.77 | 0.11 ± 0.01 |
| Formaldehyde + BPL-inactivated | 130.20 ± 0.16 | 0.13 ± 0.02 |
| BPL-inactivated | 127.40 ± 0.59 | 0.13 ± 0.03 |
| BPL + BPL-inactivated | 152.47 ± 3.18 | 0.25 ± 0.01 |
| Samples | α-Helixes | Antiparallel β-Sheets | Parallel β-Sheets | β-Turns | Random Coils |
|---|---|---|---|---|---|
| Formaldehyde-inactivated | 6.80% | 47.30% | 3.80% | 16.70% | 29.40% |
| Formaldehyde + BPL-inactivated | 6.50% | 48.20% | 3.80% | 16.50% | 29.60% |
| BPL-inactivated | 6.60% | 47.50% | 3.80% | 16.50% | 29.80% |
| BPL + BPL-inactivated | 6.50% | 47.60% | 3.80% | 16.50% | 29.70% |
| Sample | Protein Concentration (μg/mL) | Antigen Concentration (U/mL) | Ratio (U/μg) |
|---|---|---|---|
| Formaldehyde-inactivated | 332.4 | 267.9 | 0.81 |
| Formaldehyde + BPL-inactivated | 211.33 | 224.57 | 1.06 |
| BPL-inactivated | 327.56 | 623.38 | 1.90 |
| BPL + BPL-inactivated | 308.19 | 372.56 | 1.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Wei, Y.; Liang, H.; Ji, W.; Chang, Z.; Xie, S.; Wang, Y.; Li, W.; Liu, Y.; Wu, H.; et al. Comparison of Physical and Biochemical Characterizations of SARS-CoV-2 Inactivated by Different Treatments. Viruses 2022, 14, 1938. https://doi.org/10.3390/v14091938
Yu S, Wei Y, Liang H, Ji W, Chang Z, Xie S, Wang Y, Li W, Liu Y, Wu H, et al. Comparison of Physical and Biochemical Characterizations of SARS-CoV-2 Inactivated by Different Treatments. Viruses. 2022; 14(9):1938. https://doi.org/10.3390/v14091938
Chicago/Turabian StyleYu, Shouzhi, Yangyang Wei, Hongyang Liang, Wenheng Ji, Zhen Chang, Siman Xie, Yichuan Wang, Wanli Li, Yingwei Liu, Hao Wu, and et al. 2022. "Comparison of Physical and Biochemical Characterizations of SARS-CoV-2 Inactivated by Different Treatments" Viruses 14, no. 9: 1938. https://doi.org/10.3390/v14091938