
Secondary Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (HP-PRRSV2) Infection Augments Inflammatory Responses, Clinical Outcomes, and Pathogen Load in Glaesserella-parasuis-Infected Piglets
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. Virus and Bacterium
2.3. Pigs, Experimental Design, and Sampling
2.4. Clinical and Pathological Evaluation
2.5. Detection of HP-PRRSV2/Gps Antibodies by ELISA
2.6. Detection of Pathogen Load by qRT-PCR
2.7. Cytokine Assays
2.8. Statistical Analysis
3. Results
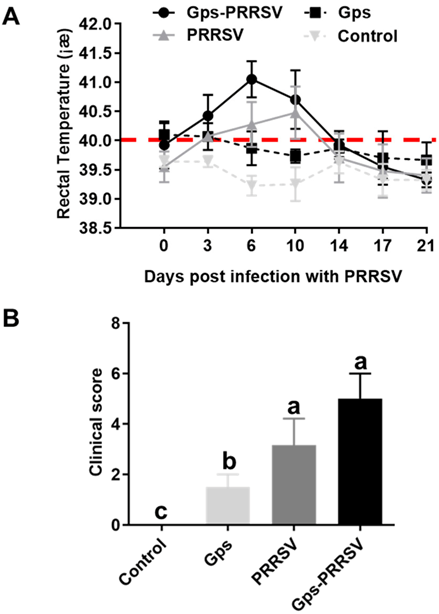
3.1. Clinical Evaluation
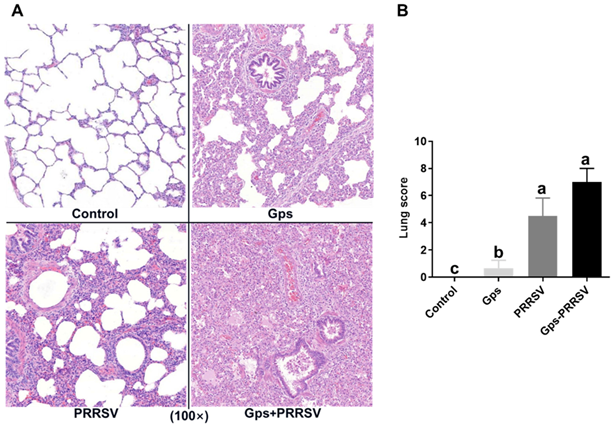
3.2. Pathological Examination
3.3. Pathogen Shedding and Load
3.4. Serum Antibody Levels of Gps and HP-PRRSV2
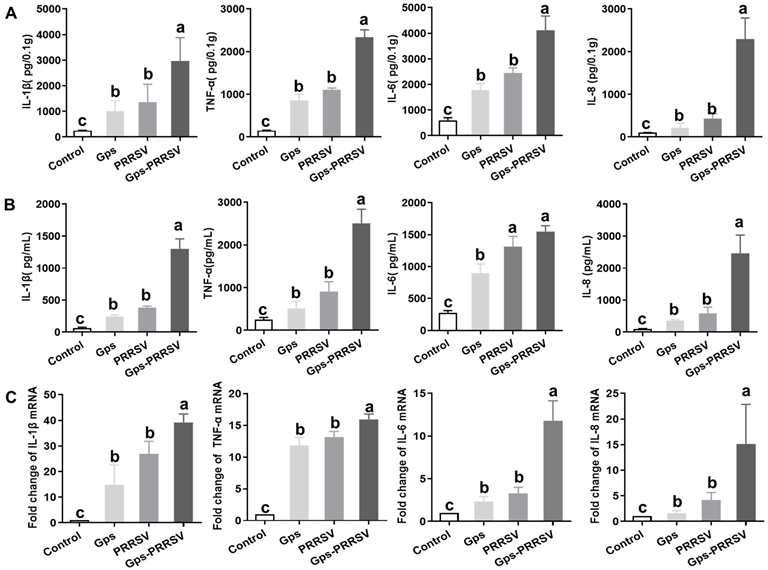
3.5. Cytokine Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, H.; Chang, X.; Zhou, J.; Wang, D.; Zhou, J.; Fan, B.; Ni, Y.; Yin, J.; Lv, L.; Zhao, Y.; et al. Co-infection analysis of bacterial and viral respiratory pathogens from clinically healthy swine in Eastern China. Vet. Med. Sci. 2021, 7, 1815–1819. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Xu, F.; Lin, Y.; Wang, Y.; Zhang, Y.; Su, K.; Li, T.; Li, H.; Song, Q. Detection of Porcine Circovirus Type 2a and Pasteurella multocida Capsular Serotype D in Growing Pigs Suffering from Respiratory Disease. Vet Sci. 2022, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Auray, G.; Lachance, C.; Wang, Y.; Gagnon, C.A.; Segura, M.; Gottschalk, M. Gottschalk. Transcriptional Analysis of PRRSV-Infected Porcine Dendritic Cell Response to Streptococcus suis Infection Reveals Up-Regulation of Inflammatory-Related Genes Expression. PLoS ONE 2016, 11, e0156019. [Google Scholar] [CrossRef] [PubMed]
- Tonni, M.; Formenti, N.; Boniotti, M.B.; Guarneri, F.; Scali, F.; Romeo, C.; Pasquali, P.; Pieters, M.; Maes, D.; Alborali, G.L. The role of co-infections in M. hyopneumoniae outbreaks among heavy fattening pigs: A field study. Vet. Res. 2022, 53, 41. [Google Scholar] [CrossRef]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Vet. Res. 2020, 51, 80. [Google Scholar] [CrossRef]
- Ouyang, T.; Zhang, X.; Liu, X.; Ren, L. Co-Infection of Swine with Porcine Circovirus Type 2 and Other Swine Viruses. Viruses 2019, 11, 185. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Li, C.; Wang, C.; Liu, Y.; Wang, G.; He, X.; Hu, L.; Liu, Y.; Cui, M.; et al. Secondary Haemophilus parasuis infection enhances highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) infection-mediated inflammatory responses. Vet. Microbiol. 2017, 204, 35–42. [Google Scholar] [CrossRef]
- Chen, N.; Xiao, Y.; Ye, M.; Li, X.; Li, S.; Xie, N.; Wei, Y.; Wang, J.; Zhu, J. High genetic diversity of Chinese porcine reproductive and respiratory syndrome viruses from 2016 to 2019. Res. Vet. Sci. 2020, 131, 38–42. [Google Scholar] [CrossRef]
- Dunkelberger, J.R.; Serão, N.V.L.; Weng, Z.; Waide, E.H.; Niederwerder, M.C.; Kerrigan, M.A.; Lunney, J.K.; Rowland, R.R.R.; Dekkers, J.C.M. Genomic regions associated with host response to porcine reproductive and respiratory syndrome vaccination and co-infection in nursery pigs. BMC Genom. 2017, 18, 865. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Loving, C.L.; Palmer, M.V.; Spear, A.; Nicholson, T.L.; Faaberg, K.S.; Lager, K.M. Comparison of Asian porcine high fever disease isolates of porcine reproductive and respiratory syndrome virus to United States isolates for their ability to cause disease and secondary bacterial infection in swine. Vet. Microbiol. 2017, 203, 6–17. [Google Scholar] [CrossRef]
- Han, J.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Pathogenesis and control of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2017, 209, 30–47. [Google Scholar] [CrossRef]
- Zhou, H.; Pan, Y.; Liu, M.; Han, Z. Prevalence of Porcine Pseudorabies Virus and Its Coinfection Rate in Heilongjiang Province in China from 2013 to 2018. Viral Immunol. 2020, 33, 550–554. [Google Scholar] [CrossRef]
- Chung, W.B.; Chan, W.H.; Chaung, H.C.; Lien, Y.; Wu, C.C.; Huang, Y.L. Real-time PCR for quantitation of porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 in naturally-infected and challenged pigs. J. Virol. Methods 2005, 124, 11–19. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Xie, C.; Ha, Z.; Shi, N.; Zhang, H.; Li, Z.; Han, J.; Xie, Y.; Qiu, X.; et al. Synergistic Pathogenicity by Coinfection and Sequential Infection with NADC30-like PRRSV and PCV2 in Post-Weaned Pigs. Viruses 2022, 14, 193. [Google Scholar] [CrossRef]
- Tregaskis, P.L.; Staines, A.; Gordon, A.; Sheridan, P.; McMenamy, M.; Duffy, C.; Collins, P.J.; Mooney, M.H.; Lemon, K. Co-infection status of novel parvovirus’s (PPV2 to 4) with porcine circovirus 2 in porcine respiratory disease complex and porcine circovirus-associated disease from 1997 to 2012. Transbound. Emerg. Dis. 2021, 68, 1979–1994. [Google Scholar] [CrossRef]
- Albina, E.; Mesplede, A.; Chenut, G.; Le Potier, M.F.; Bourbao, G.; Le Gal, S.; Leforban, Y. A serological survey on classical swine fever (CSF), Aujeszky’s disease (AD) and porcine reproductive and respiratory syndrome (PRRS) virus infections in French wild boars from 1991 to 1998. Vet. Microbiol. 2000, 77, 43–57. [Google Scholar] [CrossRef]
- Xu, M.; Wang, S.; Li, L.; Lei, L.; Liu, Y.; Shi, W.; Wu, J.; Li, L.; Rong, F.; Xu, M.; et al. Secondary infection with Streptococcus suis serotype 7 increases the virulence of highly pathogenic porcine reproductive and respiratory syndrome virus in pigs. Virol. J. 2010, 7, 184. [Google Scholar] [CrossRef]
- Palzer, A.; Eddicks, M.; Zoels, S.; Stark, J.; Reese, S.; Strutzberg-Minder, K.; Fiebig, K.; Ritzmann, M. Field evaluation of the efficacy, compatibility and serologic profiling of a combined vaccine against porcine reproductive and respiratory syndrome and Haemophilus parasuis in nursery pigs. Prev. Vet. Med. 2015, 119, 134–140. [Google Scholar] [CrossRef]
- Thacker, E.L.; Thacker, B.J.; Young, T.F.; Halbur, P.G. Effect of vaccination on the potentiation of porcine reproductive and respiratory syndrome virus (PRRSV)-induced pneumonia by Mycoplasma hyopneumoniae. Vaccine 2000, 18, 1244–1252. [Google Scholar] [CrossRef]
- Zhao, D.; Yang, B.; Yuan, X.; Shen, C.; Zhang, D.; Shi, X.; Zhang, T.; Cui, H.; Yang, J.; Chen, X.; et al. Advanced Research in Porcine Reproductive and Respiratory Syndrome Virus Co-infection with Other Pathogens in Swine. Front. Vet. Sci. 2021, 8, 699561. [Google Scholar] [CrossRef]
- Zuckermann, F.A.; Husmann, R.; Chen, W.; Roady, P.; Pfeiff, J.; Leistikow, K.R.; Duersteler, M.; Son, S.; King, M.R.; Augspurger, N.R. Bacillus-Based Direct-Fed Microbial Reduces the Pathogenic Synergy of a Coinfection with Salmonella enterica Serovar Choleraesuis and Porcine Reproductive and Respiratory Syndrome Virus. Infect. Immun. 2022, 90, e0057421. [Google Scholar] [CrossRef] [PubMed]
- Day, D.N.; Sparks, J.W.; Karriker, L.A.; Stalder, K.J.; Wulf, L.W.; Zhang, J.; Kinyon, J.M.; Stock, M.L.; Gehring, R.; Wang, C.; et al. Impact of an experimental PRRSV and Streptococcus suis coinfection on the pharmacokinetics of ceftiofur hydrochloride after intramuscular injection in pigs. J. Vet. Pharmacol. Ther. 2015, 38, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Nelsen, A.; Lin, C.-M.; Hause, B.M. Porcine Parvovirus 2 Is Predominantly Associated with Macrophages in Porcine Respiratory Disease Complex. Front. Vet. Sci. 2021, 8, 726884. [Google Scholar] [CrossRef] [PubMed]
- van Dixhoorn, I.D.E.; Beest, D.E.T.; Bolhuis, J.E.; Parmentier, H.K.; Kemp, B.; van Mourik, S.; Stockhofe-Zurwieden, N.; Van Reenen, C.G.; Rebel, J.M.J. Animal-Based Factors Prior to Infection Predict Histological Disease Outcome in Porcine Reproductive and Respiratory Syndrome Virus- and Actinobacillus pleuropneumoniae-Infected Pigs. Front. Vet. Sci. 2021, 8, 742877. [Google Scholar] [CrossRef]
- Yan, X.; Zeng, J.; Li, X.; Zhang, Z.; Din, A.U.; Zhao, K.; Zhou, Y. High incidence and characteristic of PRRSV and resistant bacterial Co-Infection in pig farms. Microb. Pathog. 2020, 149, 104536. [Google Scholar]
- Fu, S.; Xu, L.; Li, S.; Qiu, Y.; Liu, Y.; Wu, Z.; Ye, C.; Hou, Y.; Hu, C.A. Baicalin suppresses NLRP3 inflammasome and nuclear factor-kappa B (NF-kappaB) signaling during Haemophilus parasuis infection. Vet. Res. 2016, 47, 80. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Q.; Wu, W.; Chen, M.; Zhang, P.; Guo, M.; Lin, H.; Ma, Z.; Zhou, H.; Fan, H. Glaesserella parasuis serotype 5 breaches the porcine respiratory epithelial barrier by inducing autophagy and blocking the cell membrane Claudin-1 replenishment. PLoS Pathog. 2022, 18, e1010912. [Google Scholar] [CrossRef]
- Fablet, C.; Marois, C.; Dorenlor, V.; Eono, F.; Eveno, E.; Jolly, J.; Le Devendec, L.; Kobisch, M.; Madec, F.; Rose, N. Bacterial pathogens associated with lung lesions in slaughter pigs from 125 herds. Res. Vet. Sci. 2012, 93, 627–630. [Google Scholar] [CrossRef]
- Hansen, M.S.; Pors, S.E.; Jensen, H.E.; Bille-Hansen, V.; Bisgaard, M.; Flachs, E.M.; Nielsen, O.L. An investigation of the pathology and pathogens associated with porcine respiratory disease complex in Denmark. J. Comp. Pathol. 2010, 143, 120–131. [Google Scholar] [CrossRef]
- Oliveira, S.; Pijoan, C. Haemophilus parasuis: New trends on diagnosis, epidemiology and control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef]
- Yu, J.; Wu, J.; Zhang, Y.; Guo, L.; Cong, X.; Du, Y.; Li, J.; Sun, W.; Shi, J.; Peng, J.; et al. Concurrent highly pathogenic porcine reproductive and respiratory syndrome virus infection accelerates Haemophilus parasuis infection in conventional pigs. Vet. Microbiol. 2012, 158, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kwok, A.H.Y.; Jiang, J.; Zou, Y.; Zheng, F.; Chen, P.; Hou, C.; Leung, F.C.; Jiang, P. Complete genome analysis of a Haemophilus parasuis serovar 12 strain from China. PLoS ONE. 2013, 8, e68350. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Zhang, C.; Aragon, V.; Zhou, X.; Zou, M.; Wu, C.; Shen, Z. Investigation of Haemophilus parasuis from healthy pigs in China. Vet. Microbiol. 2019, 231, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Rampelotto, P.H.; dos Santos, A.C.R.; Varela, A.P.M.; Takeuti, K.L.; Loiko, M.R.; Mayer, F.Q.; Roehe, P.M. Comparative Analysis of the Upper Respiratory Bacterial Communities of Pigs with or without Respiratory Clinical Signs: From Weaning to Finishing Phase. Biology 2022, 11, 1111. [Google Scholar] [CrossRef]
- Kavanova, L.; Matiaskova, K.; Leva, L.; Stepanova, H.; Nedbalcova, K.; Matiasovic, J.; Faldyna, M.; Salat, J. Concurrent infection with porcine reproductive and respiratory syndrome virus and Haemophilus parasuis in two types of porcine macrophages: Apoptosis, production of ROS and formation of multinucleated giant cells. Vet. Res. 2017, 48, 28. [Google Scholar] [CrossRef]
- Kavanová, L.; Matiašková, K.; Levá, L.; Nedbalcová, K.; Matiašovic, J.; Faldyna, M.; Salát, J. Concurrent infection of monocyte-derived macrophages with porcine reproductive and respiratory syndrome virus and Haemophilus parasuis: A role of IFNα in pathogenesis of co-infections. Vet. Microbiol. 2018, 225, 64–71. [Google Scholar] [CrossRef]
- Luo, L.; Van Dixhoorn, I.D.E.; Reimert, I.; Kemp, B.; Bolhuis, J.E.; Parmentier, H.K. Effect of enriched housing on levels of natural (auto-)antibodies in pigs co-infected with porcine reproductive and respiratory syndrome virus (PRRSV) and Actinobacillus pleuropneumoniae. Vet. Res. 2017, 48, 75. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, L.; Song, T.; Pan, X.; Li, X.; Lu, G.; Tang, Y.; Wu, X.; Zhao, B.; Zhang, R. A candidate multi-epitope vaccine against porcine reproductive and respiratory syndrome virus and Mycoplasma hyopneumoniae induces robust humoral and cellular response in mice. Vaccine 2022, 40, 2370–2378. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, Y.; Xia, Q.; Guan, Z.; Zhang, J.; Li, B.; Qiu, Y.; Liu, K.; Shao, D.; Ma, Z.; et al. Genetic characterization of porcine reproductive and respiratory syndrome virus from Eastern China during 2017–2022. Front. Microbiol. 2022, 13, 971817. [Google Scholar] [CrossRef]
- Goyal, S.M. Porcine reproductive and respiratory syndrome. J. Vet. Diagn. Investig. 1993, 5, 656–664. [Google Scholar] [CrossRef]
- Albina, E.; Baron, T.; Leforban, Y. Blue-eared pig disease in Brittany. Vet. Rec. 1992, 130, 58–59. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, R.; Qiao, S.; Zhang, G. The prevalent status and genetic diversity of porcine reproductive and respiratory syndrome virus in China: A molecular epidemiological perspective. Virol. J. 2018, 15, 2. [Google Scholar] [CrossRef]
- Neumann, E.J.; Kliebenstein, J.B.; Johnson, C.D.; Mabry, J.W.; Bush, E.J.; Seitzinger, A.H.; Green, A.L.; Zimmerman, J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005, 227, 385–392. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, L.; Zhang, J.; Ge, X.; Zhou, R.; Zheng, H.; Geng, G.; Guo, X.; Yang, H. Nsp9 and Nsp10 contribute to the fatal virulence of highly pathogenic porcine reproductive and respiratory syndrome virus emerging in China. PLoS Pathog. 2014, 10, e1004216. [Google Scholar] [CrossRef]
- Tong, G.Z.; Zhou, Y.J.; Hao, X.F.; Tian, Z.J.; An, T.Q.; Qiu, H.J. Highly pathogenic porcine reproductive and respiratory syndrome, China. Emerg. Infect. Dis. 2007, 13, 1434–1436. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Bo, K.; Wang, X.; Tang, B.; Yang, B.; Jiang, W.; Jiang, P. Emergence of a highly pathogenic porcine reproductive and respiratory syndrome virus in the Mid-Eastern region of China. Vet. J. 2007, 174, 577–584. [Google Scholar] [CrossRef]
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; McCullough, S.; Morrison, R.B.; Joo, H.S.; et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Investig. 1992, 4, 117–126. [Google Scholar] [CrossRef]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; Ter Laak, E.A.; Bloemraad, M.; De Kluyver, E.P.; Kragten, C.; Van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bào, Y.; Ng, T.F.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef]
- Chrun, T.; Maze, E.A.; Vatzia, E.; Martini, V.; Paudyal, B.; Edmans, M.D.; McNee, A.; Manjegowda, T.; Salguero, F.J.; Wanasen, N.; et al. Simultaneous Infection with Porcine Reproductive and Respiratory Syndrome and Influenza Viruses Abrogates Clinical Protection Induced by Live Attenuated Porcine Reproductive and Respiratory Syndrome Vaccination. Front. Immunol. 2021, 12, 758368. [Google Scholar] [CrossRef]
- Wang, X.; Yang, X.; Zhou, R.; Zhou, L.; Ge, X.; Guo, X.; Yang, H. Genomic characterization and pathogenicity of a strain of type 1 porcine reproductive and respiratory syndrome virus. Virus Res. 2016, 225, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Han, Z.; Liu, Z.; Meng, F.; Wang, H.; Cao, L.; Li, Y.; Jiao, Q.; Liu, S.; Liu, M. Epidemiological investigation of porcine pseudorabies virus and its coinfection rate in Shandong Province in China from 2015 to 2018. J. Vet. Sci. 2020, 21, e36. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.B.; Frossard, J.P.; Pallares, F.J.; Gough, J.; Stadejek, T.; Graham, S.P.; Steinbach, F.; Drew, T.W.; Salguero, F.J. Pathology and Virus Distribution in the Lung and Lymphoid Tissues of Pigs Experimentally Inoculated with Three Distinct Type 1 PRRS Virus Isolates of Varying Pathogenicity. Transbound. Emerg. Dis. 2016, 63, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Sun, Y.F.; Jiang, X.; Zhang, A.; Ma, J.F.; Yu, X.X.; Li, L.A.; Yu, H. Early infection of Streptococcus suis serotype 2 increases the virulence of highly pathogenic porcine reproductive and respiratory syndrome MLV-like virus in pigs. Res. Vet. Sci. 2020, 130, 68–72. [Google Scholar] [CrossRef]
- Matiaskova, K.; Kavanova, L.; Kulich, P.; Gebauer, J.; Nedbalcova, K.; Kudlackova, H.; Tesarik, R.; Faldyna, M. The Role of Antibodies Against the Crude Capsular Extract in the Immune Response of Porcine Alveolar Macrophages to In Vitro Infection of Various Serovars of Glaesserella (Haemophilus) parasuis. Front. Immunol. 2021, 12, 635097. [Google Scholar] [CrossRef]
- Guo, B.Q.; Chen, Z.S.; Liu, W.X.; Cui, Y.Z. Isolation of PRRSV from suspected PRRS aborted fetuses. Chin. J. Prev. Vet. Med. 1996, 2, 3–7. [Google Scholar]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Liu, Y.; Yang, J.; Guo, L.; Ren, S.; Chen, Z.; Liu, Z.; Zhang, Y.; Qiu, W.; et al. Porcine reproductive and respiratory syndrome virus NADC30-like strain accelerates Streptococcus suis serotype 2 infection in vivo and in vitro. Transbound. Emerg. Dis. 2019, 66, 729–742. [Google Scholar] [CrossRef]
- Chen, N.; Cao, Z.; Yu, X.; Deng, X.; Zhao, T.; Wang, L.; Liu, Q.; Li, X.; Tian, K. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J. Gen. Virol. 2011, 92, 880–892. [Google Scholar] [CrossRef]
- Zhang, H.L.; Zhang, W.L.; Xiang, L.R.; Leng, C.L.; Tian, Z.J.; Tang, Y.D.; Cai, X.H. Emergence of novel porcine reproductive and respiratory syndrome viruses (ORF5 RFLP 1-7-4 viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, L.; Li, C.; Zhao, J.; Liu, N.; Li, Y.; Wang, J.; Liu, L. Identification of enterobacteria in viscera of pigs afflicted with porcine reproductive and respiratory syndrome and other viral co-infections. Microb. Pathog. 2020, 147, 104385. [Google Scholar] [CrossRef]
- Mahmmod, Y.S.; Correa-Fiz, F.; Aragon, V. Variations in association of nasal microbiota with virulent and non-virulent strains of Glaesserella (Haemophilus) parasuis in weaning piglets. Vet. Res. 2020, 51, 7. [Google Scholar] [CrossRef]
- Wang, Q.; Chang, X.; Liu, M.; Lu, Q.; Zhu, M.; Lin, H.; Fan, H. Glaesserella parasuis serotype 4 HPS4-YC disrupts the integrity of the swine tracheal epithelial barrier and facilitates bacterial translocation. Vet. Res. 2021, 52, 135. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, J.; Zhang, X.; Zhao, C.; Zhou, S.; Du, C.; Tan, Y.; Zhang, Y.; Shi, K. Transcriptome profiling identifies immune response genes against porcine reproductive and respiratory syndrome virus and Haemophilus parasuis co-infection in the lungs of piglets. J. Vet. Sci. 2022, 23, e2. [Google Scholar] [CrossRef]
- Pomorska-Mol, M.; Dors, A.; Kwit, K.; Czyzewska-Dors, E.; Pejsak, Z. Coinfection modulates inflammatory responses, clinical outcome and pathogen load of H1N1 swine influenza virus and Haemophilus parasuis infections in pigs. BMC Vet. Res. 2017, 13, 376. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; Chen, H.; Yu, J.; Luo, Y.; Zheng, P.; He, J. Effects of dietary ferulic acid supplementation on growth performance and skeletal muscle fiber type conversion in weaned piglets. J. Sci. Food Agric. 2021, 101, 5116–5123. [Google Scholar] [CrossRef]
- Duvigneau, J.C.; Hartl, R.T.; Groiss, S.; Gemeiner, M. Quantitative simultaneous multiplex real-time PCR for the detection of porcine cytokines. J. Immunol. Methods. 2005, 306, 16–27. [Google Scholar] [CrossRef]
- Jablonski, A.; Zebek, S.; Kolacz, R.; Pejsak, Z. Usefulness of PCR/RFLP and ERIC PCR techniques for epidemiological study of Haemophilus parasuis infections in pigs. Pol. J. Vet. Sci. 2011, 14, 111–116. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gimenez-Lirola, L.G.; Halbur, P.G. Polymicrobial respiratory disease in pigs. Anim. Health Res. Rev. 2011, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Chen, A.; Wang, X.; Pan, Z.; Xu, S.; Yu, H.; Zhang, B.; Liao, M. The Glaesserella parasuis phosphoglucomutase is partially required for lipooligosaccharide synthesis. Vet. Res. 2020, 51, 97. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Li, F.; Chen, K.; Ding, W.; Xue, Y.; Wang, Y.; Wang, H.; Ding, K.; Zhao, Z. Comparison of the Glaesserella parasuis Virulence in Mice and Piglets. Front. Vet. Sci. 2021, 8, 659244. [Google Scholar] [CrossRef] [PubMed]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Wiencek, I.; Hartmann, M.; Merkel, J.; Trittmacher, S.; Kreienbrock, L.; Hennig-Pauka, I. Temporal Patterns of Phenotypic Antimicrobial Resistance and Coinfecting Pathogens in Glaesserella parasuis Strains Isolated from Diseased Swine in Germany from 2006 to 2021. Pathogens 2022, 11, 721. [Google Scholar] [CrossRef]
- Yang, Y.; An, T.; Gong, D.; Li, D.; Peng, J.; Leng, C.; Yuan, Z.; Tong, G.; Tian, Z.; Zhang, D. Identification of porcine serum proteins modified in response to HP-PRRSV HuN4 infection by two-dimensional differential gel electrophoresis. Vet. Microbiol. 2012, 158, 237–246. [Google Scholar] [CrossRef]
- Guo, B.; Lager, K.M.; Henningson, J.N.; Miller, L.C.; Schlink, S.N.; Kappes, M.A.; Kehrli, M.E., Jr.; Brockmeier, S.L.; Nicholson, T.L.; Yang, H.-C.; et al. Experimental infection of United States swine with a Chinese highly pathogenic strain of porcine reproductive and respiratory syndrome virus. Virology 2013, 435, 372–384. [Google Scholar] [CrossRef]
- Liu, J.K.; Zhou, X.; Zhai, J.Q.; Li, B.; Wei, C.H.; Dai, A.L.; Yang, X.Y.; Luo, M.L. Emergence of a novel highly pathogenic porcine reproductive and respiratory syndrome virus in China. Transbound. Emerg. Dis. 2017, 64, 2059–2074. [Google Scholar] [CrossRef]
- Oba, P.; Dione, M.M.; Wieland, B.; Mwiine, F.N.; Erume, J. Correlations between lung pneumonic lesions and serologic status for key respiratory pathogens in slaughtered pigs in northern Uganda. Porc. Health Manag. 2021, 7, 53. [Google Scholar] [CrossRef]
- Lee, J.A.; Oh, Y.R.; Hwang, M.A.; Lee, J.B.; Park, S.Y.; Song, C.S.; Choi, I.S.; Lee, S.W. Mycoplasma hyorhinis is a potential pathogen of porcine respiratory disease complex that aggravates pneumonia caused by porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2016, 177, 48–51. [Google Scholar] [CrossRef]
- Turlewicz-Podbielska, H.; Czyzewska-Dors, E.; Pomorska-Mol, M. Respiratory viral infections drive different lung cytokine profiles in pigs. BMC Vet. Res. 2021, 17, 5. [Google Scholar] [CrossRef]
- Kavanova, L.; Prodelalova, J.; Nedbalcova, K.; Matiasovic, J.; Volf, J.; Faldyna, M.; Salat, J. Immune response of porcine alveolar macrophages to a concurrent infection with porcine reproductive and respiratory syndrome virus and Haemophilus parasuis in vitro. Vet. Microbiol. 2015, 180, 28–35. [Google Scholar] [CrossRef]
- Avadhanula, V.; Rodriguez, C.A.; Devincenzo, J.P.; Wang, Y.; Webby, R.J.; Ulett, G.C.; Adderson, E.E. Respiratory viruses augment the adhesion of bacterial pathogens to respiratory epithelium in a viral species- and cell type-dependent manner. J. Virol. 2006, 80, 1629–1636. [Google Scholar] [CrossRef]
- Didierlaurent, A.; Goulding, J.; Patel, S.; Snelgrove, R.; Low, L.; Bebien, M.; Lawrence, T.; Van Rijt, L.S.; Lambrecht, B.N.; Sirard, J.C.; et al. Sustained desensitization to bacterial Toll-like receptor ligands after resolution of respiratory influenza infection. J. Exp. Med. 2008, 205, 323–329. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scoring Criteria | Respiratory Signs | Nasal Discharge, Coughing, Anorexia, and Sneezing | Temperature | Lung Lesions |
|---|---|---|---|---|
| 0 | normal (<34 breaths/min) | absent | normal | no lesions |
| 1 | slightly elevated (35–40 breaths/min) | present | rectal temperature exceeded 40 °C | lesions affecting <25% of the lobe surface |
| 2 | moderately elevated (41–45 breaths/min with slight abdominal breathing) | / | / | lesions affecting 25–49% of the lobe surface |
| 3 | elevated (>46 breaths/min with distinct abdominal breathing) | / | / | lesions affecting 50–74% of the lobe surface |
| 4 | / | / | / | lesions affecting >75% of the lobe surface |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, Z.; Pang, L.; Ouyang, Y.; Jiang, Y.; Zhang, J.; Qiu, Y.; Li, Z.; Li, B.; Liu, K.; Shao, D.; et al. Secondary Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (HP-PRRSV2) Infection Augments Inflammatory Responses, Clinical Outcomes, and Pathogen Load in Glaesserella-parasuis-Infected Piglets. Vet. Sci. 2023, 10, 365. https://doi.org/10.3390/vetsci10050365
Guan Z, Pang L, Ouyang Y, Jiang Y, Zhang J, Qiu Y, Li Z, Li B, Liu K, Shao D, et al. Secondary Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (HP-PRRSV2) Infection Augments Inflammatory Responses, Clinical Outcomes, and Pathogen Load in Glaesserella-parasuis-Infected Piglets. Veterinary Sciences. 2023; 10(5):365. https://doi.org/10.3390/vetsci10050365
Chicago/Turabian StyleGuan, Zhixin, Linlin Pang, Yan Ouyang, Yifeng Jiang, Junjie Zhang, Yafeng Qiu, Zongjie Li, Beibei Li, Ke Liu, Donghua Shao, and et al. 2023. "Secondary Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus (HP-PRRSV2) Infection Augments Inflammatory Responses, Clinical Outcomes, and Pathogen Load in Glaesserella-parasuis-Infected Piglets" Veterinary Sciences 10, no. 5: 365. https://doi.org/10.3390/vetsci10050365