
Impairment of CD4+ T and Memory B Cell Responses but Normal Memory CD8+T-Cell Activation on Crohn’s Disease after COVID-19 Vaccination: A Twin Case

 , ,
, ,
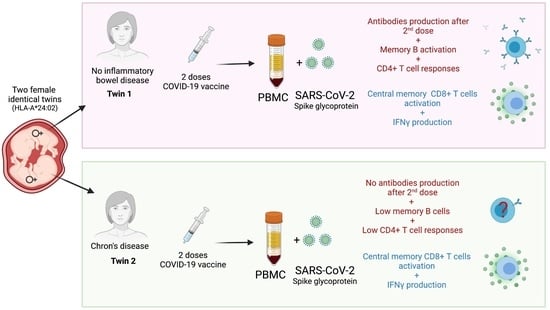
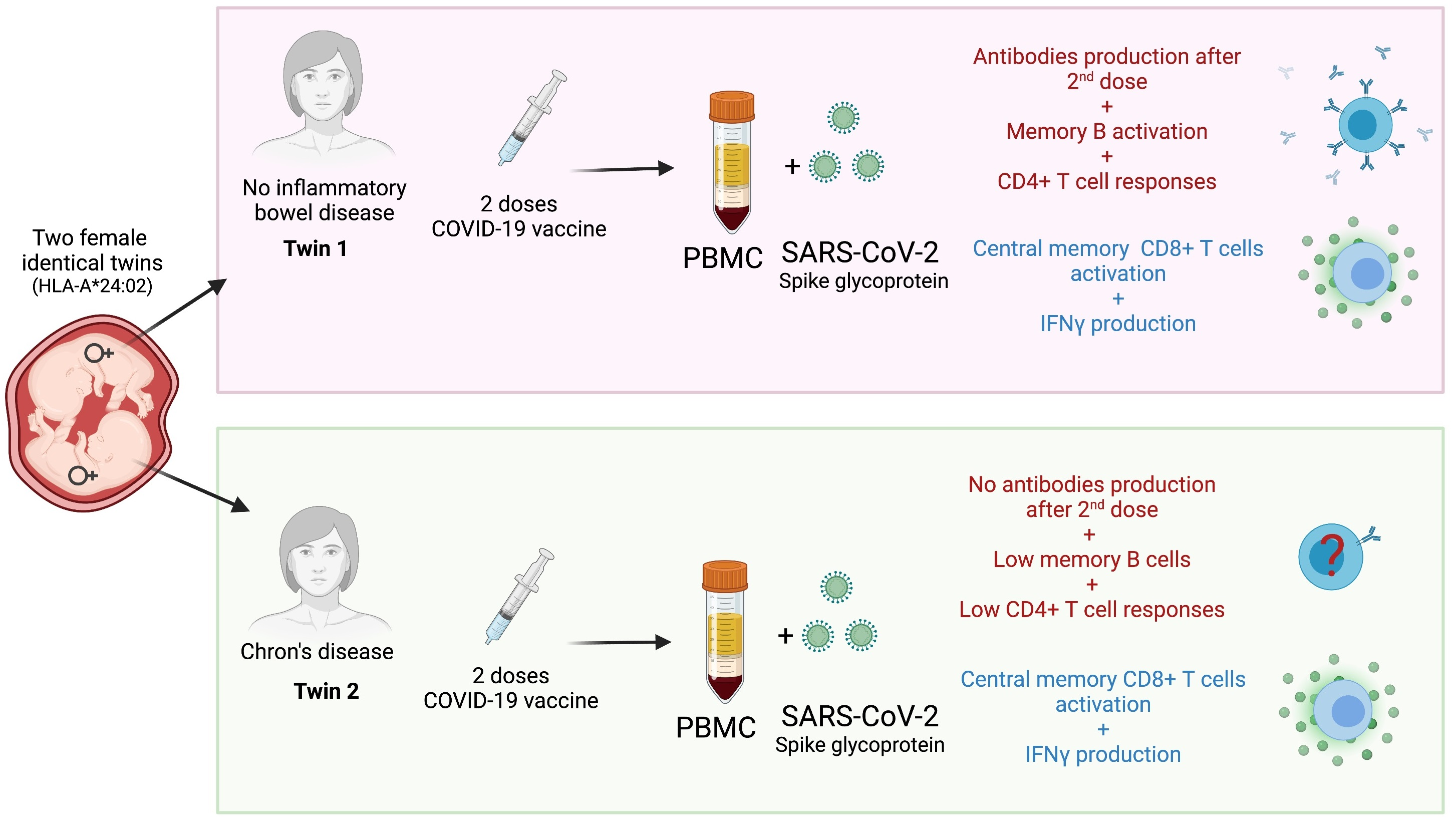
Abstract
:
1. Introduction
2. Material and Methods
2.1. Investigation
2.2. Laboratory Assays
2.3. Statistical Analysis
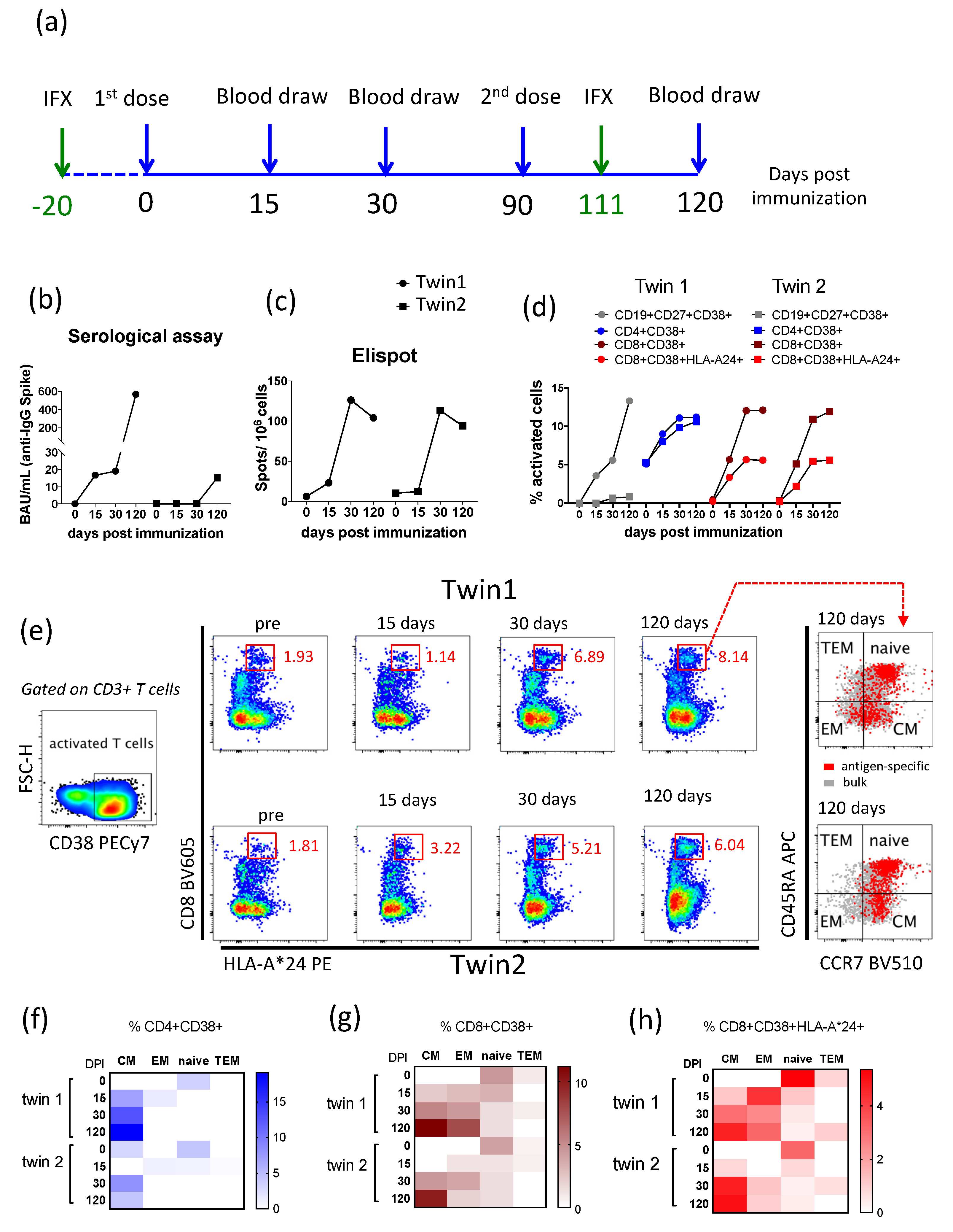
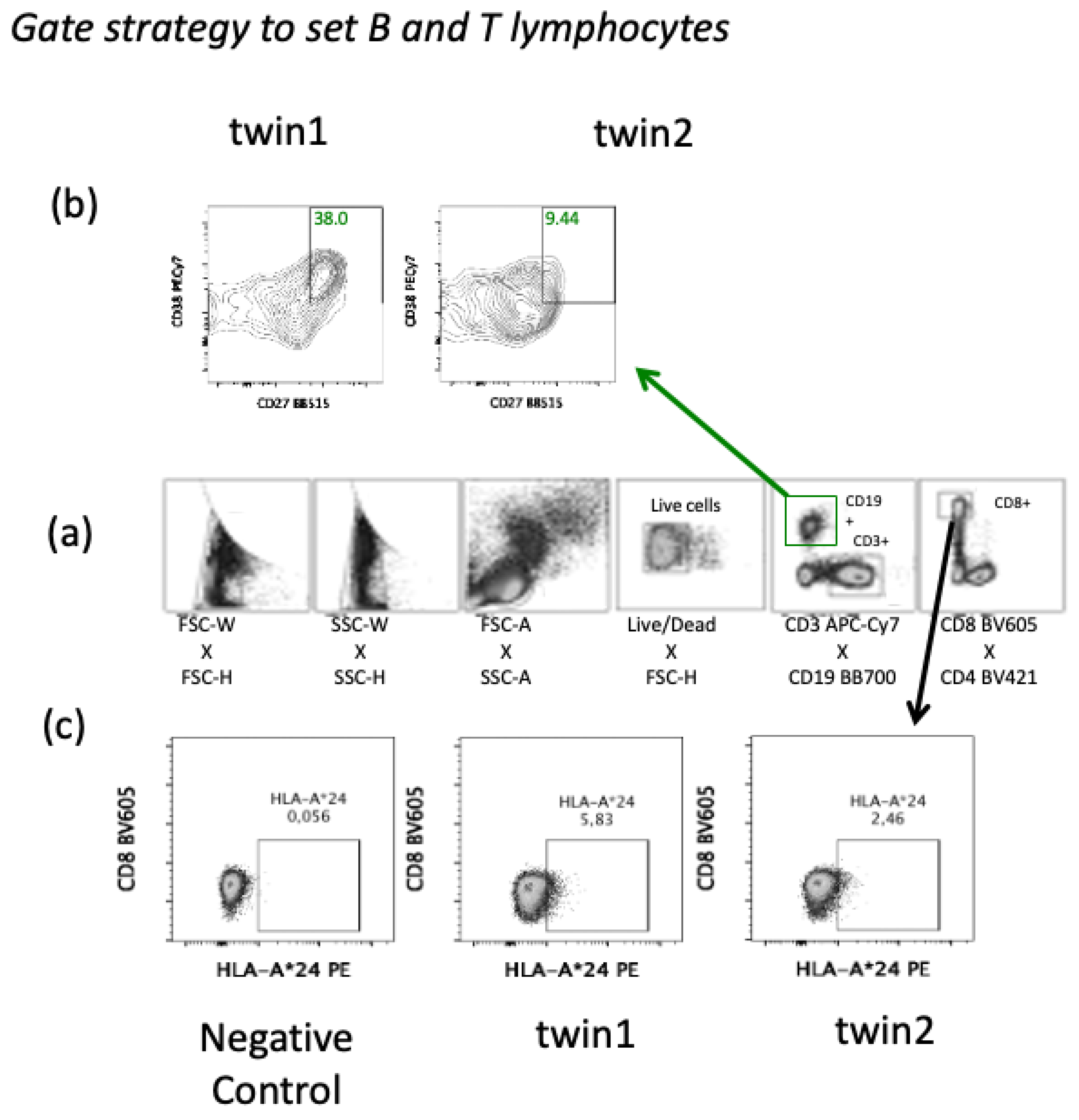
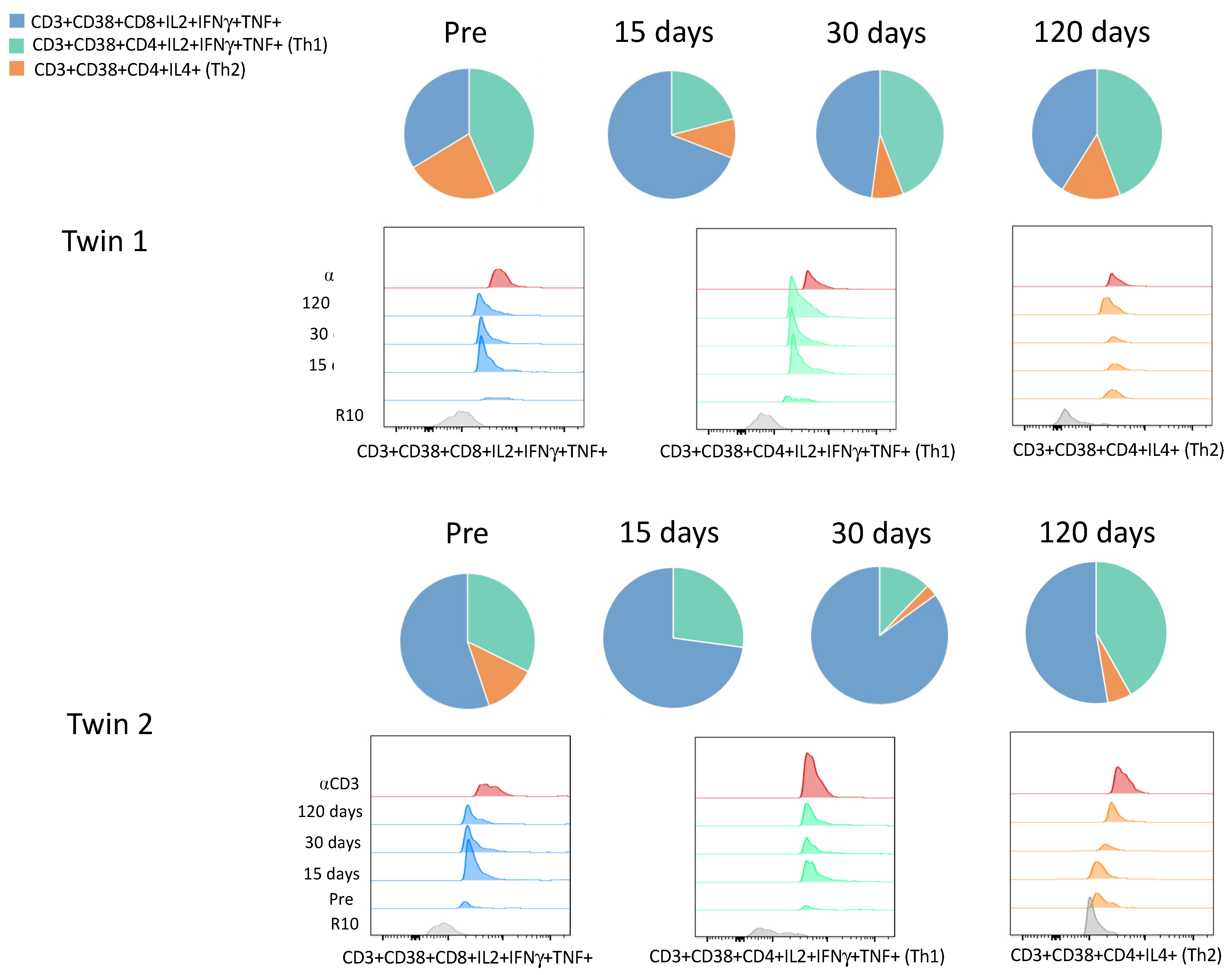
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, E.; Du, H.; Gardner, L. An Interactive Web-Based Dashboard to Track COVID-19 in Real Time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef]
- Statistics and Research - Coronavirus (COVID-19) Vaccinations. Available online: https://ourworldindata.org/covid-vaccinations (accessed on 22 July 2021).
- Caron, B.; Neuville, E.; Peyrin-Biroulet, L. Inflammatory Bowel Disease and COVID-19 Vaccination: A Patients’ Survey. Dig. Dis. Sci. 2021. [Google Scholar] [CrossRef]
- Chebli, J.M.F.; Queiroz, N.S.F.; Damião, A.O.M.C.; Chebli, L.A.; de Magalhães Costa, M.H.; Parra, R.S. How to Manage Inflammatory Bowel Disease during the COVID-19 Pandemic: A Guide for the Practicing Clinician. World J. Gastroenterol. 2021, 27, 1022–1042. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.; Fatima, R.; Haghbin, H.; Lee-Smith, W.; Nawras, A. The Incidence and Outcomes of COVID-19 in IBD Patients: A Rapid Review and Meta-Analysis. Inflamm. Bowel Dis. 2020, 26, e132–e133. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, N.A.; Lin, S.; Goodhand, J.R.; Chanchlani, N.; Hamilton, B.; Bewshea, C.; Nice, R.; Chee, D.; Cummings, J.F.; Fraser, A.; et al. Infliximab Is Associated with Attenuated Immunogenicity to BNT162b2 and ChAdOx1 NCoV-19 SARS-CoV-2 Vaccines in Patients with IBD. Gut 2021, 70, 1884–1893. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.-Y.; Dixon, R.; Martinez Pazos, V.; Gnjatic, S.; Colombel, J.-F.; Cadwell, K.; Gold, S.; Helmus, D.; Neil, J.A.; Sota, S.; et al. Serologic Response to Messenger RNA Coronavirus Disease 2019 Vaccines in Inflammatory Bowel Disease Patients Receiving Biologic Therapies. Gastroenterology 2021, 161, 715–718.e4. [Google Scholar] [CrossRef] [PubMed]
- Wallace, K.L. Immunopathology of Inflammatory Bowel Disease. World J. Gastroenterol. 2014, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Geremia, A.; Biancheri, P.; Allan, P.; Corazza, G.R.; Di Sabatino, A. Innate and Adaptive Immunity in Inflammatory Bowel Disease. Autoimmun. Rev. 2014, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Ardizzone, S.; Bianchi Porro, G. Biologic Therapy for Inflammatory Bowel Disease. Drugs 2005, 65, 2253–2286. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Edwards, J.E. Type 1/Type 2 Immunity in Infectious Diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Methot, N.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; da Silva Antunes, R.; Frazier, A.; et al. Negligible Impact of SARS-CoV-2 Variants on CD4+ and CD8+ T Cell Reactivity in COVID-19 Exposed Donors and Vaccinees. Immunology 2021. [Google Scholar] [CrossRef]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.-L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM Structure of the 2019-NCoV Spike in the Prefusion Conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirai, S.; Hara, M.; Sakata, Y.; Tsuruoka, N.; Yamamoto, K.; Shimoda, R.; Gomi, Y.; Yoshii, H.; Fujimoto, K.; Iwakiri, R. Immunogenicity of Quadrivalent Influenza Vaccine for Patients with Inflammatory Bowel Disease Undergoing Immunosuppressive Therapy. Inflamm. Bowel Dis. 2018, 24, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Wellens, J.; Colombel, J.-F.; Satsangi, J.J.; Wong, S.-Y. SARS-CoV-2 Vaccination in IBD: Past Lessons, Current Evidence, and Future Challenges. J. Crohns Colitis 2021, 15, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, I.; Lindley, K.J.; Londei, M.; Quaratino, S. Anti Tumour Necrosis-α Therapy Increases the Number of FOXP3+ Regulatory T Cells in Children Affected by Crohn’s Disease. Immunology 2008, 125, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Noble, A.; Durant, L.; Hoyles, L.; Mccartney, A.L.; Man, R.; Segal, J.; Costello, S.P.; Hendy, P.; Reddi, D.; Bouri, S.; et al. Deficient Resident Memory T Cell and CD8 T Cell Response to Commensals in Inflammatory Bowel Disease. J. Crohns Colitis 2020, 14, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Franco Salinas, G.; De Rycke, L.; Barendregt, B.; Paramarta, J.E.; Hreggvidstdottir, H.; Cantaert, T.; van der Burg, M.; Tak, P.P.; Baeten, D. Anti-TNF Treatment Blocks the Induction of T Cell-Dependent Humoral Responses. Ann. Rheum. Dis. 2013, 72, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Alonso, G.T.; Fomin, D.S.; Rizzo, L.V. Human Follicular Helper T Lymphocytes Critical Players in Antibody Responses. Einstein São Paulo 2021, 19, eRB6077. [Google Scholar] [CrossRef] [PubMed]
- Cárdeno, A.; Magnusson, M.K.; Quiding-Järbrink, M.; Lundgren, A. Activated T Follicular Helper-like Cells Are Released into Blood after Oral Vaccination and Correlate with Vaccine Specific Mucosal B-Cell Memory. Sci. Rep. 2018, 8, 2729. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Twin 1 | Twin 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| DPI | 0 | 15 | 30 | 120 | 0 | 15 | 30 | 120 |
| %CD19+ | 15.16 | 14.27 | 13.17 | 13.30 | 11.90 | 13.46 | 13.39 | 12.01 |
| %CD3+ | 62.40 | 65.50 | 62.10 | 62.10 | 60.10 | 62.50 | 65.70 | 60.90 |
| %CD4+ | 42.10 | 48.90 | 44.70 | 36.80 | 34.00 | 39.30 | 47.00 | 38.50 |
| %CD8+ | 21.20 | 21.60 | 21.40 | 20.80 | 12.70 | 15.50 | 16.80 | 10.80 |
| CD4/CD8 | 2.00 | 2.30 | 2.10 | 1.80 | 2.70 | 2.50 | 2.80 | 3.60 |
| HLA-AI type | A*24:02 | A*24:02 | ||||||
| HLA-AII type | A*68:02 | A*68:02 | ||||||
| BMI | 28.9 | 27.1 | ||||||
| AEPV | ||||||||
| 1st dose | yes | mild fever (37.7 °C), local pain, myalgia, headache | yes | local pain, myalgia, headache | ||||
| 2nd dose | no | no | ||||||
| Twin 1 | Twin 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 15 | Day 30 | Day 120 | Day 0 | Day 15 | Day 30 | Day 120 | |
| CD4+IL2+IFNγ+TNF+ (Th1) | 0.40 | 2.64 | 4.70 | 6.44 | 0.41 | 2.33 | 0.40 | 4.39 |
| CD4+IL4+ (Th2) | 0.21 | 1.10 | 0.85 | 2.14 | 0.23 | 0.01 | 0.09 | 0.58 |
| CD4+TNF+ | 0.96 | 38.37 | 27.35 | 17.00 | 5.44 | 3.32 | 3.47 | 11.02 |
| CD4+IL2+ | 0.00 | 0.00 | 13.58 | 15.85 | 0.49 | 3.33 | 0.67 | 1.83 |
| CD4+IFNγ+ | 0.00 | 8.95 | 10.57 | 12.5 | 1.48 | 5.62 | 0.71 | 2.98 |
| CD4+IFNγ+IL2+ | 0.00 | 0.00 | 1.82 | 2.83 | 0.00 | 0.22 | 0.01 | 0.04 |
| CD4+IL2+TNF+ | 0.01 | 0.50 | 3.43 | 0.29 | 0.12 | 0.14 | 0.19 | 0.21 |
| CD4+IFNγ+TNF+ | 0.00 | 7.56 | 2.30 | 0.48 | 0.40 | 0.61 | 0.00 | 0.83 |
| CD8+IL2+ IFNγ+TNF+ | 0.51 | 7.70 | 5.11 | 5.98 | 0.83 | 7.09 | 2.77 | 5.53 |
| CD8+IFNγ+IL2+ | 0.00 | 5.41 | 3.29 | 1.05 | 1.48 | 5.88 | 3.05 | 0.88 |
| CD8+IFNγ+TNF+ | 0.00 | 5.52 | 4.93 | 5.34 | 0.04 | 1.19 | 0.38 | 0.87 |
| CD8+IL2+TNF+ | 0.06 | 4.84 | 3.07 | 3.22 | 0.11 | 2.8 | 0.85 | 1.07 |
| CD8+TNF+ | 1.20 | 45.46 | 35.97 | 32.9 | 1.12 | 21.38 | 8.30 | 15.21 |
| CD8+IFNγ+ | 0.00 | 15.60 | 13.86 | 18.30 | 2.95 | 18.25 | 10.81 | 17.60 |
| CD8+IL2+ | 0.12 | 14.33 | 11.00 | 18.29 | 6.39 | 19.21 | 9.73 | 6.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melgaço, F.G.; Azamor, T.; Villar, L.M.; Ano Bom, A.P.D.; Melgaço, J.G. Impairment of CD4+ T and Memory B Cell Responses but Normal Memory CD8+T-Cell Activation on Crohn’s Disease after COVID-19 Vaccination: A Twin Case. Viruses 2021, 13, 2143. https://doi.org/10.3390/v13112143
Melgaço FG, Azamor T, Villar LM, Ano Bom APD, Melgaço JG. Impairment of CD4+ T and Memory B Cell Responses but Normal Memory CD8+T-Cell Activation on Crohn’s Disease after COVID-19 Vaccination: A Twin Case. Viruses. 2021; 13(11):2143. https://doi.org/10.3390/v13112143
Chicago/Turabian StyleMelgaço, Fabiana Gil, Tamiris Azamor, Livia Melo Villar, Ana Paula Dinis Ano Bom, and Juliana Gil Melgaço. 2021. "Impairment of CD4+ T and Memory B Cell Responses but Normal Memory CD8+T-Cell Activation on Crohn’s Disease after COVID-19 Vaccination: A Twin Case" Viruses 13, no. 11: 2143. https://doi.org/10.3390/v13112143