
SARS-CoV-2 in a Mink Farm in Italy: Case Description, Molecular and Serological Diagnosis by Comparing Different Tests

 , , , , , , , ,
, , , , , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Description and Sampling
2.2. Diagnostic Examinations
2.3. Serological Investigations
2.4. Sequencing and Phylogenetic Analysis
3. Results
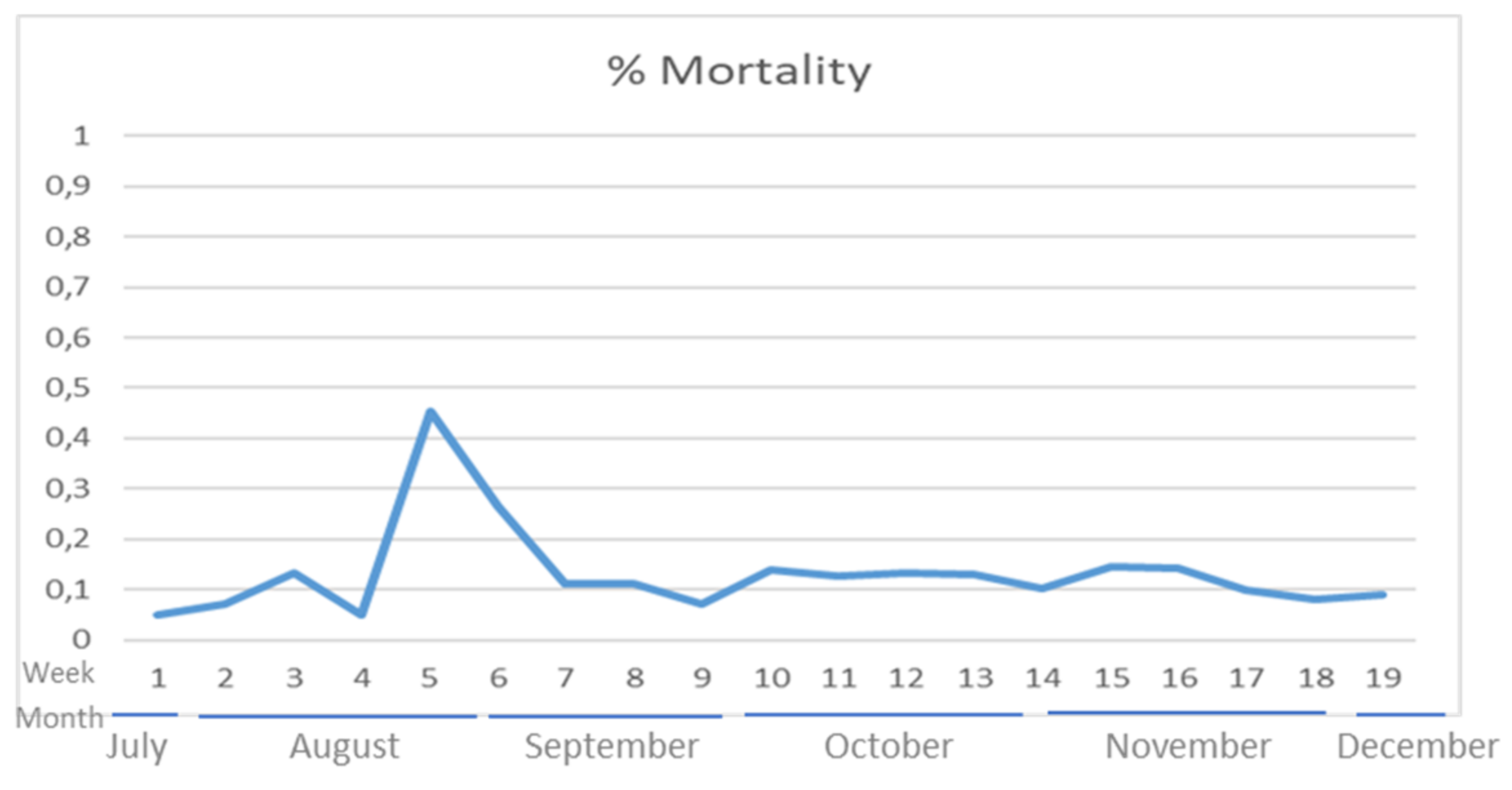
3.1. Diagnostic Examinations
3.2. Serological Investigations
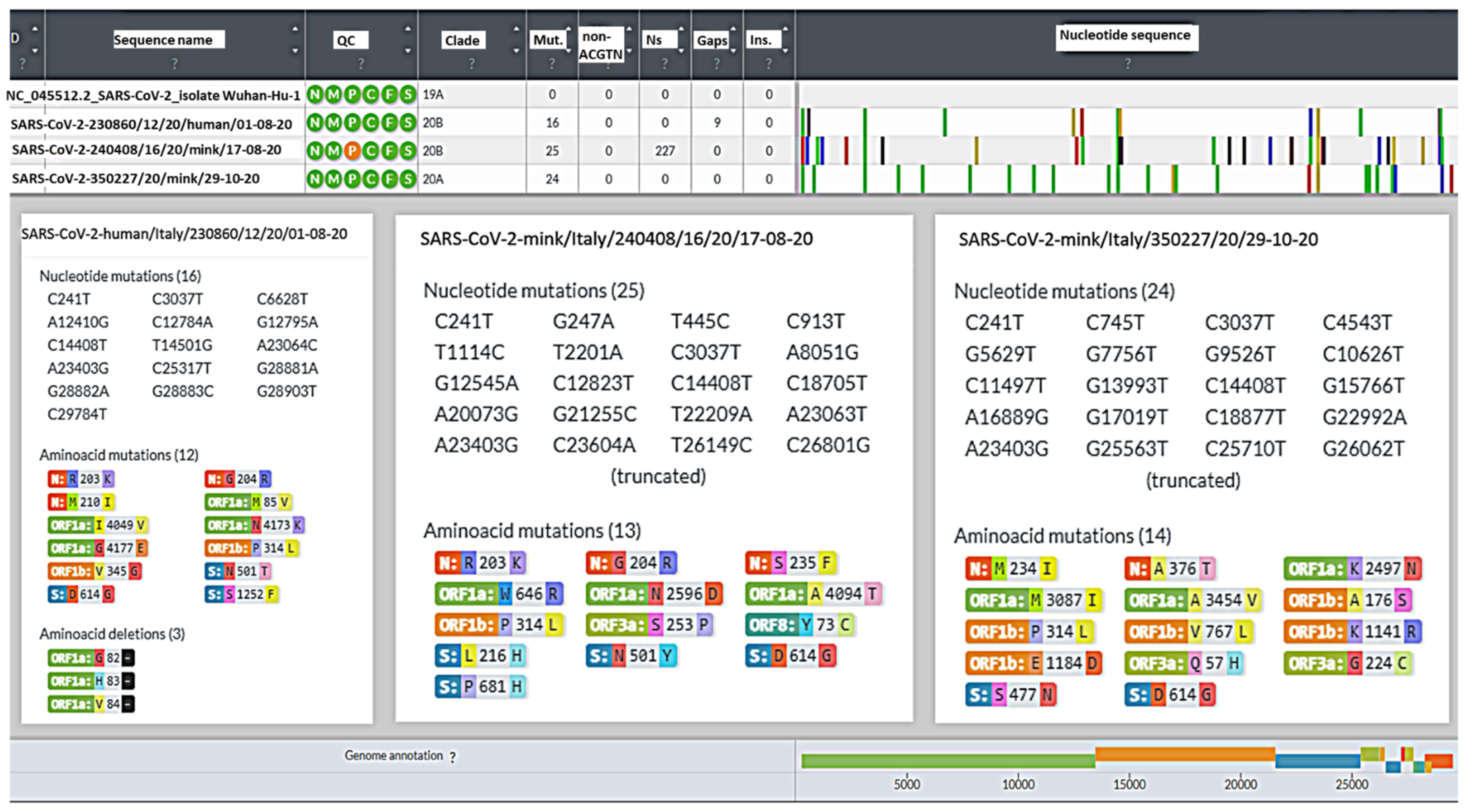
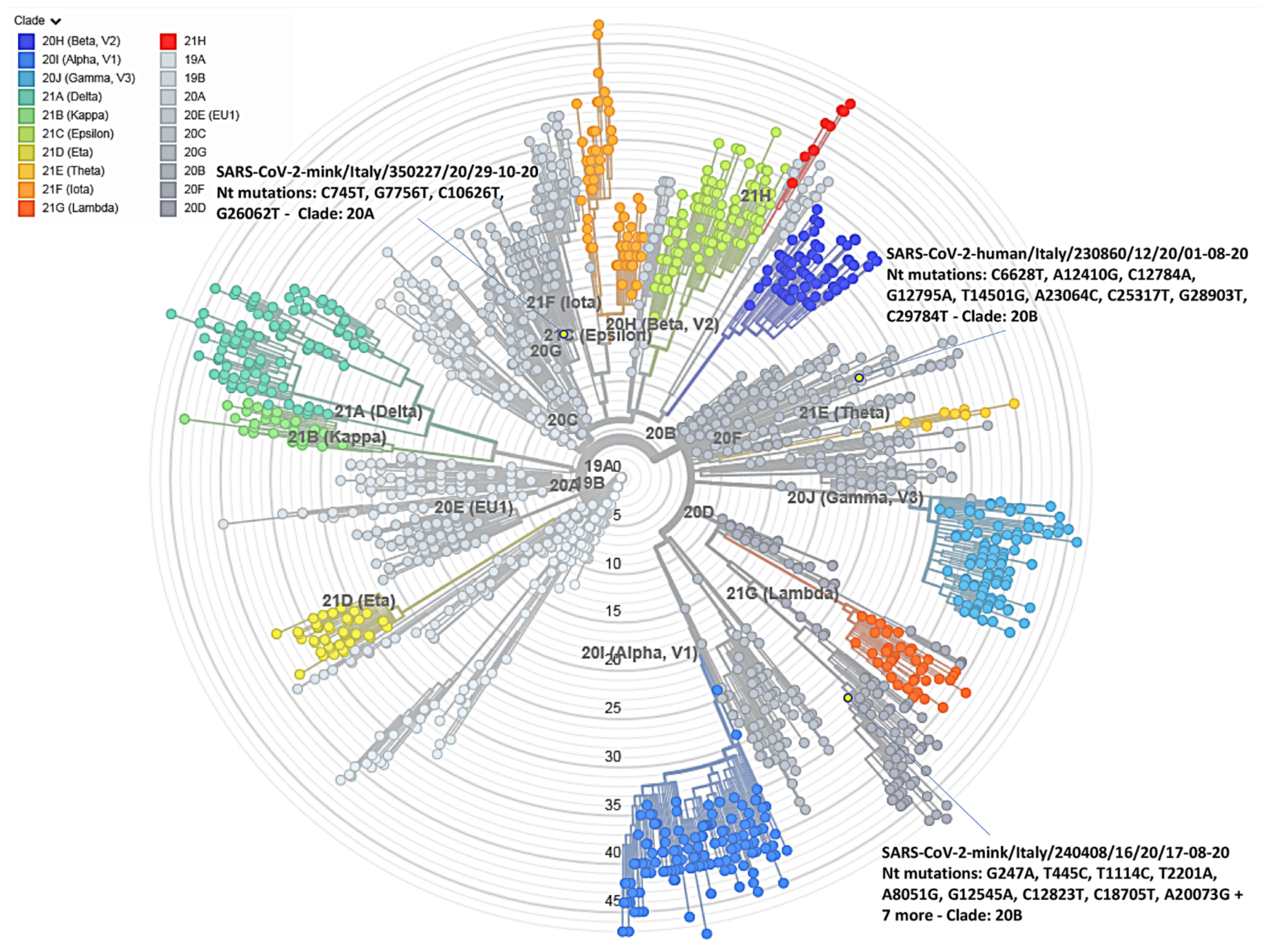
3.3. SARS-CoV-2 Genome Detection and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 23 November 2021).
- Dhama, K.; Patel, S.K.; Sharun, K.; Pathak, M.; Tiwari, R.; Yatoo, M.I.; Malik, Y.S.; Sah, R.; Rabaan, A.A.; Panwar, P.K.; et al. SARS-CoV-2 jumping the species barrier: Zoonotic lessons from SARS, MERS and recent advances to combat this pandemic virus. Travel Med. Infect. Dis. 2020, 37, 101830. [Google Scholar] [CrossRef] [PubMed]
- Oude Munnink, B.B.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; van der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M.; et al. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Mallapaty, S. Animal source of the coronavirus continues to elude scientists. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- Do Vale, B.; Lopes, A.P.; Fontes, M.D.C.; Silvestre, M.; Cardoso, L.; Coelho, A.C. Bats, pangolins, minks and other animals—Villains or victims of SARS-CoV-2? Vet. Res. Commun. 2021, 45, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pomorska-Mól, M.; Włodarek, J.; Gogulski, M.; Rybska, M. Review: SARS-CoV-2 infection in farmed minks—An overview of current knowledge on occurrence, disease and epidemiology. Animal 2021, 15, 100272. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control; Boklund, A.; Gortazar, C.; Pasquali, P.; Roberts, H.; Nielsen, S.S.; Stahl, K.; Stegeman, A.; Baldinelli, F.; Broglia, A.; et al. Scientific Opinion on the monitoring of SARS-CoV-2 infection in mustelids. EFSA J. 2021, 19, 6459. [Google Scholar] [CrossRef]
- SARS-CoV-2 in Animals Used for Fur Farming, OIE. Available online: https://www.oie.int/app/uploads/2021/03/glews-risk-assessment-fur-animals-sars-cov-2.pdf (accessed on 20 January 2021).
- OIE. SARS-CoV-2 in Animals—Situation Report 2. Available online: https://www.oie.int/app/uploads/2021/09/sars-cov-2-situation-report-4.pdf (accessed on 31 August 2021).
- Boklund, A.; Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Lohse, L.; Strandbygaard, B.; Jørgensen, C.S.; Olesen, A.S.; Hjerpe, F.B.; Petersen, H.H.; et al. SARS-CoV-2 in Danish Mink Farms: Course of the Epidemic and a Descriptive Analysis of the Outbreaks in 2020. Animals 2021, 11, 164. [Google Scholar] [CrossRef]
- Hammer, A.S.; Quaade, M.L.; Rasmussen, T.B.; Fonager, J.; Rasmussen, M.; Mundbjerg, K.; Lohse, L.; Strandsbygaard, B.; Jørgensen, C.S.; Alfaro-Núñez, A.; et al. SARS-CoV-2 transmission between mink (Neovison vison) and humans, Denmark. Emerg. Infect. Dis. 2021, 27, 547. [Google Scholar] [CrossRef] [PubMed]
- World Organization for Animal Health (Wahis). OIE Immediate Notification 22 April 2021, Italy. Available online: https://wahis.woah.org/#/report-info?reportId=32262 (accessed on 30 May 2022).
- Ordinanza 21 Novembre 2020 Gazzetta Ufficiale. Available online: https://www.gazzettaufficiale.it/eli/id/2020/11/23/20A06501/sg (accessed on 21 November 2020).
- Corman, V.; Bleicker, T.; Brünink, S.; Drosten, C. Diagnostic Detection of 2019-nCoV by Real-Time RT-PCR Protocol. Available online: https://www.who.int/docs/default-source/coronaviruse/protocol-v2-1.pdf (accessed on 17 January 2020).
- Real-Time RT-PCR Panel for Detection 2019-Novel Coronavirus. Centers for Disease Control and Prevention, Respiratory Viruses Branch, Division of Viral Diseases. Available online: https://www.who.int/docs/default-source/coronaviruse/uscdcrt-pcr-panel-for-detection-instructions.pdf?sfvrsn=3aa07934_2 (accessed on 24 January 2020).
- Frisk, A.L.; König, M.; Moritz, A.; Baumgärtner, W. Detection of canine distemper virus nucleoprotein RNA by reverse transcription-PCR using serum, whole blood, and cerebrospinal fluid from dogs with distemper. J. Clin. Microbiol. 1999, 37, 3634–3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a real-time reverse transcriptase PCR assay for type A influenza virus and the avian H5 and H7 hemagglutinin subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- BSOP 54 National Standard Method Inoculation of Culture Media, Issued by Standards Unit, Evaluations and Standards Laboratory, Specialist and Reference Microbiology Division, Issue No. 4, Issue Date 03.05.05. Available online: www.evaluations-standards.org.uk (accessed on 3 May 2005).
- ISO/TR 6579-3:2014(en) Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 3: Guidelines for Serotyping of Salmonella spp. 2014. Available online: https://www.iso.org/obp/ui/#iso:std:iso:tr:6579:-3:ed-1:v1:en (accessed on 8 September 2020).
- Detection of Clostridium Strains Producing Botulinum Toxin of Type A, B, E, F, C, D, CD and DC (internal method ISS N-RL CNRB31.011, 2019). Available online: https://www.iss.it/documents/20126/0/CNRB30.011.pdf/59a7faf1-15f5-dd54-ae13-d066de9dcea2?t=1582362668086 (accessed on 12 July 2019).
- Fernández-Bellon, H.; Rodon, J.; Fernández-Bastit, L.; Almagro, V.; Padilla-Solé, P.; Lorca-Oró, C.; Valle, R.; Roca, N.; Grazioli, S.; Trogu, T.; et al. Monitoring Natural SARS-CoV-2 Infection in Lions (Panthera leo) at the Barcelona Zoo: Viral Dynamics and Host Responses. Viruses 2021, 13, 1683. [Google Scholar] [CrossRef] [PubMed]
- Rijkers, G.; Murk, J.L.; Wintermans, B.; van Looy, B.; van den Berge, M.; Veenemans, J.; Stohr, J.; Reusken, C.; van der Pol, P.; Reimerink, J. Differences in antibody kinetics and functionality between severe and mild severe acute respiratory syndrome coronavirus 2 infections. J. Infect. Dis. 2020, 222, 1265. [Google Scholar] [CrossRef] [PubMed]
- De Sabato, L.; Vaccari, G.; Knijn, A.; Ianiro, G.; Di Bartolo, I.; Morabito, S. SARS-CoV-2 RECoVERY: A multi-platform open-source bioinformatic pipeline for the automatic construc-tion and analysis of SARS-CoV-2 genomes from NGS sequencing data. bioRxiv 2021. [Google Scholar] [CrossRef]
- Nextclade Web 1.5.3. Available online: https://clades.nextstrain.org/ (accessed on 10 August 2021).
- Ncov/Clades.tsv at Master Nextstrain/Ncov—GitHub. Available online: https://github.com/nextstrain/ncov/blob/master/defaults/clades.tsv (accessed on 8 August 2021).
- Auspice.us. Available online: https://auspice.us/ (accessed on 8 August 2021).
- Devaux, C.A.; Pinault, L.; Delerce, J.; Raoult, D.; Levasseur, A.; Frutos, R. Spread of Mink SARS-CoV-2 Variants in Humans: A Model of Sarbecovirus Interspecies Evolution. Front. Microbiol. 2021, 12, 675528. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goraichuk, I.V.; Arefiev, V.; Stegniy, B.T.; Gerilovych, A.P. Zoonotic and Reverse Zoonotic Transmissibility of SARS-CoV-2. Virus Res. 2021, 302, 198473. [Google Scholar] [CrossRef]
- Vergara-Alert, J.; Rodon, J.; Carrillo, J.; Te, N.; Izquierdo-Useros, N.; Rodríguez de la Concepción, M.L.; Ávila-Nieto, C.; Guallar, V.; Valencia, A.; Cantero, G.; et al. Pigs are not susceptible to SARS-CoV-2 infection but are a model for viral immunogenicity studies. Transbound. Emerg. Dis. 2021, 68, 1721–1725. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Puig, M.; Rodon, J.; Avila-Nieto, C.; Carrillo, J.; Cantero, G.; Terrón, M.T.; Cruz, S.; Parera, M.; Noguera-Julián, M.; et al. Detection of SARS-CoV-2 in a cat owned by a COVID-19-affected patient in Spain. Proc. Natl. Acad. Sci. USA 2020, 117, 24790–24793. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.C.; Tiu, C.; Hu, Z.; Chen, V.C.W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2–spike protein–protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Embregts, C.W.E.; Verstrepen, B.; Langermans, J.A.M.; Böszörményi, K.P.; Sikkema, R.S.; de Vries, R.D.; Hoffmann, D.; Wernike, K.; Smit, L.A.M.; Zhao, S.; et al. Evaluation of a multi-species SARS-CoV-2 surrogate virus neutralization test. One Health 2021, 13, 100313. [Google Scholar] [CrossRef]
- Berguido, F.J.; Burbelo, P.D.; Bortolami, A.; Bonfante, F.; Wernike, K.; Hoffmann, D.; Balkema-Buschmann, A.; Beer, M.; Dundon, W.G.; Lamien, C.E.; et al. Serological Detection of SARS-CoV-2 Antibodies in Naturally-Infected Mink and Other Experimentally-Infected Animals. Viruses 2021, 13, 1649. [Google Scholar] [CrossRef] [PubMed]
- Lassaunière, R.; Fonager, J.; Rasmussen, M.; Frische, A.; Polacek, C.; Rasmussen, T.B.; Lohse, L.; Belsham, G.J.; Underwood, A.; Winckelmann, A.A.; et al. In vitro Characterization of Fitness and Convalescent Antibody Neutralization of SARS-CoV-2 Cluster 5 Variant Emerging in Mink at Danish Farms. Front. Microbiol. 2021, 25, 698944. [Google Scholar] [CrossRef] [PubMed]
- Elaswad, A.; Fawzy, M.; Basiouni, S.; Shehata, A.A. Mutational spectra of SARS-CoV-2 isolated from animals. PeerJ 2020, 8, e10609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Mann, M.; Syed, Z.A.; Reynolds, H.M.; Tian, E.; Samara, N.L.; Zeldin, D.C.; Tabak, L.A.; Ten Hagen, K.G. Furin cleavage of the SARS-CoV-2 spike is modulated by O-glycosylation. Proc. Natl. Acad. Sci. USA 2021, 118, e2109905118. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, T.B.; Fonager, J.; Jørgensen, C.S.; Lassaunière, R.; Hammer, A.S.; Quaade, M.L.; Boklund, A.; Lohse, L.; Strandbygaard, B.; Rasmussen, M.; et al. Infection, recovery and re-infection of farmed mink with SARS-CoV-2. PLoS Pathog. 2021, 17, e1010068. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n.samples | IDScreen SARS-CoV-2N IgGIndirect Multispecies Conjugate | ERADIKIT™ COVID19-Multispecies | Double-Antigen N ELISA IZSLER | Wantai SARS-CoV-2 Ab ELISA | GenScript Surrogate VNT | Proteogenix SARS-CoV-2 Surrogate VNT | VNT (Rijkers et al., 2020) |
|---|---|---|---|---|---|---|---|
| CUT-Off | S/P% ≥ 40 | S/P% ≥ 20 | S/P% ≥ 10 | OD > 1000 | % Inhibition > 30 | % Inhibition | VNT50 ≥ 1/10 |
| 1 | <0.3 | 8.0 | 11.4 | 4549 | 95 | 0 | 1/80 |
| 2 | <0.3 | 100.3 | 40.5 | 4257 | 96 | 0 | 1/80 |
| 3 | <0.3 | 200.7 | 37.2 | 4923 | 96 | 6 | 1/160 |
| 4 | <0.3 | 287.9 | 107.0 | 4584 | 95 | 0 | 1/160 |
| 5 | <0.3 | 155.2 | 73.6 | 4051 | 96 | 0 | 1/160 |
| 6 | <0.3 | NT | NT | 4217 | 95 | NT | 1/160 |
| 7 | <0.3 | 39.7 | 28.1 | 4209 | 96 | 1 | 1/160 |
| 8 | <0.3 | 120.2 | 64.5 | 4157 | 96 | 0 | 1/80 |
| 9 | <0.3 | 190.9 | 61.2 | 3481 | 78 | 0 | 1/80 |
| 10 | <0.3 | 125.3 | 57.4 | 4171 | 95 | 10 | 1/80 |
| 11 | <0.3 | 103.7 | 36.0 | 4305 | 95 | 0 | 1/40 |
| 12 | <0.3 | 247.8 | 64.2 | 4104 | 94 | 23 | 1/160 |
| 13 | <0.3 | 279.3 | 83.5 | 4462 | 95 | 0 | 1/320 |
| 14 | <0.3 | 17.0 | 14.6 | 4310 | 48 | 0 | 1/10 |
| 15 | <0.3 | NT | 114.4 | 4438 | 95 | NT | 1/40 |
| 16 | <0.3 | 167.0 | 49.5 | 4500 | 95 | 0 | 1/80 |
| 17 | <0.3 | 106.5 | 35.9 | 5064 | 96 | 0 | 1/80 |
| 18 | <0.3 | 51.6 | 23.1 | 4416 | 95 | 0 | 1/160 |
| 19 | <0.3 | 108.3 | 38.0 | 4365 | 96 | 0 | 160 |
| 20 | <0.3 | 160.0 | 43.2 | 4247 | 95 | 0 | 1/160 |
| 21 | <0.3 | 17.7 | 2.2 | 4954 | 81 | 0 | 1/40 |
| 22 | <0.3 | 270.5 | 47.1 | 4688 | 78 | 0 | 1/10 |
| 23 | <0.3 | 292.4 | 87.1 | 6068 | 96 | 16 | 1/160 |
| 24 | <0.3 | 194.8 | 69.1 | 4415 | 49 | 0 | 1/10 |
| 25 | <0.3 | 210.6 | 57.2 | 5290 | 96 | 0 | 1/160 |
| 26 | <0.3 | 93.0 | 73.6 | 4512 | 94 | 0 | 1/160 |
| 27 | <0.3 | 39.4 | 73.3 | 5027 | 96 | 0 | 1/80 |
| 28 | <0.3 | 94.0 | 33.8 | 5165 | 94 | 0 | 1/160 |
| 29 | <0.3 | 177.2 | 53.6 | 6016 | 91 | 0 | 1/80 |
| 30 | <0.3 | 178.3 | 44.2 | 4840 | 96 | 0 | 1/80 |
| 31 | <0.3 | 8.8 | 15.1 | 4694 | 94 | 0 | 1/80 |
| 32 | <0.3 | 33.0 | 18.2 | 4472 | 96 | 0 | 1/160 |
| 33 | <0.3 | 14.4 | 0.21 | 4870 | 67 | 0 | 1/10 |
| 34 | <0.3 | 3.1 | 0.2 | 5171 | 95 | 0 | 1/80 |
| 35 | <0.3 | 99.1 | 27.0 | 5171 | 89 | 0 | 1/40 |
| 36 | <0.3 | 137.9 | 50.2 | 4460 | 89 | 0 | 1/160 |
| 37 | <0.3 | 27.4 | 6.12 | 4419 | 44 | 0 | 1/10 |
| 39 | <0.3 | NT | NT | 4875 | 95 | NT | 1/160 |
| 40 | <0.3 | 135.4 | 24.3 | 5419 | 95 | 0 | 1/80 |
| 41 | <0.3 | 1.8 | 12.2 | 6021 | 95 | 3 | 1/160 |
| 42 | <0.3 | 54.5 | 4 | 4817 | 95 | 0 | 1/40 |
| 43 | <0.3 | 238.4 | 96.5 | 5118 | 95 | 0 | 1/80 |
| 45 | <0.3 | 45.7 | 17.4 | 4907 | 96 | 6 | 1/40 |
| 46 | <0.3 | 277.1 | 112.5 | 4845 | 95 | 11 | 1/640 |
| 47 | <0.3 | 13.4 | 0.66 | 6068 | 80 | 0 | 1/40 |
| 48 | <0.3 | 234.1 | 65.3 | 5222 | 94 | 0 | 1/80 |
| 49 | <0.3 | 31.5 | 7.7 | 4892 | 94 | 0 | 1/80 |
| 50 | <0.3 | 38.8 | 17.4 | 4590 | 95 | 17 | 1/320 |
| 51 | <0.3 | 275.4 | 53 | 4687 | 95 | 0 | 1/160 |
| 52 | <0.3 | NT | NT | 4536 | 94 | NT | NT |
| 53 | <0.3 | 158.2 | 63.7 | 4477 | 94 | 0 | 1/80 |
| 54 | <0.3 | 26.8 | 14.8 | 4477 | 95 | 0 | 1/160 |
| 55 | <0.3 | 79.3 | 20.3 | 4521 | 96 | 11 | 1/160 |
| 56 | <0.3 | 164.9 | 90.8 | 4337 | 94 | 0 | 1/160 |
| 57 | <0.3 | 124.7 | 43.6 | 4932 | 95 | 0 | 1/640 |
| 58 | <0.3 | 23.3 | 15.3 | 5601 | 95 | 0 | 1/320 |
| 59 | <0.3 | NT | NT | 4615 | 94 | NT | 1/20 |
| 60 | <0.3 | 36.2 | 20.0 | 4716 | 88 | 0 | 1/40 |
| 61 | <0.3 | NT | NT | 6078 | 95 | NT | NT |
| 62 | <0.3 | 52.2 | 30.0 | 4902 | 95 | 0 | 1/160 |
| 63 | <0.3 | 142.7 | 44.2 | 4692 | 51 | 0 | 1/20 |
| 64 | <0.3 | 228.6 | 91.0 | 4838 | 96 | 0 | 1/160 |
| 65 | <0.3 | 64.1 | 15.1 | 4537 | 88 | 0 | 1/40 |
| 66 | <0.3 | 176.3 | 92.3 | 4509 | 95 | 0 | 1/160 |
| 67 | <0.3 | 26.0 | 19.5 | 4434 | 95 | 0 | 1/160 |
| 68 | <0.3 | 15.9 | 19.2 | 4201 | 94 | 0 | 1/80 |
| 69 | <0.3 | 84.1 | 25.2 | 4652 | 95 | 0 | 1/160 |
| 70 | <0.3 | 123.1 | 50.2 | 4236 | 95 | 0 | 1/160 |
| 71 | <0.3 | 29.5 | 13.5 | 4270 | 89 | 0 | 1/40 |
| 72 | <0.3 | 245.4 | 58.8 | 4115 | 94 | 0 | 1/80 |
| 73 | <0.3 | 115.2 | 86.9 | 3995 | 96 | 0 | 1/160 |
| 74 | <0.3 | NT | NT | 3999 | 89 | NT | 1/40 |
| n.samples | IDScreen SARS-CoV-2N IgGIndirect Multispecies conjugate | ERADIKIT™ COVID19-Multispecies | Double-Antigen N ELISA IZSLER | Wantai SARS-CoV-2 Ab ELISA | GenScript Surrogate VNT | Proteogenix SARS-CoV-2 surrogate VNT | VNT (Rijkers et al., 2020) |
| CUT-off | S/P% ≥ 40 | S/P% ≥ 20 | S/P% ≥ 10 | OD > 1000 | % inhibition > 30 | % inhibition | VNT50 ≥ 1/10 |
| 75 | <0.3 | 11.7 | 4.9 | 98 | 11 | NT | <1/5 |
| 76 | <0.3 | 12.6 | 3.8 | 83 | 0 | NT | <1/5 |
| 77 | <0.3 | 10.0 | 5 | 74 | 9 | NT | <1/5 |
| 78 | <0.3 | 13.8 | 4.6 | 77 | 9 | NT | <1/5 |
| 79 | <0.3 | 13.6 | 3.8 | 35 | 5 | NT | <1/5 |
| 80 | <0.3 | 6.0 | 4.1 | 87 | 0 | NT | <1/5 |
| 81 | <0.3 | 5.8 | 4.5 | 84 | 0 | NT | <1/5 |
| 82 | <0.3 | 6.8 | 4.3 | 159 | 2 | NT | <1/5 |
| 83 | <0.3 | 3.2 | 4.2 | 139 | 4 | NT | <1/5 |
| 84 | <0.3 | 8.6 | 4.5 | 109 | 0 | NT | <1/5 |
| 85 | <0.3 | 10.0 | 4.3 | 135 | 2 | NT | <1/5 |
| 86 | <0.3 | 13.1 | 4.7 | 39 | 5 | NT | <1/5 |
| 87 | <0.3 | 10 | 3.9 | 136 | 3 | NT | <1/5 |
| 88 | <0.3 | 4.2 | 6 | 137 | 5 | NT | <1/5 |
| 89 | <0.3 | 18.7 | 5.6 | 148 | 7 | NT | <1/5 |
| 90 | <0.3 | 8.1 | 5.2 | 216 | 3 | NT | <1/5 |
| 91 | <0.3 | 1.8 | 3.7 | 29 | 0 | NT | <1/5 |
| 92 | <0.3 | 5.5 | 4.2 | 68 | 6 | NT | <1/5 |
| 93 | <0.3 | 3.3 | 4.6 | 84 | 4 | NT | <1/5 |
| 94 | NT | 0.4 | 4 | 20 | 0 | NT | <1/5 |
| 95 | NT | 3 | 4.4 | 113 | 1 | NT | <1/5 |
| 96 | NT | 5.8 | 4.1 | 82 | 0 | NT | <1/5 |
| 97 | NT | 10 | 5.2 | 84 | 0 | NT | <1/5 |
| 98 | NT | 12 | 4.8 | 174 | 4 | NT | <1/5 |
| 99 | NT | 5 | 3.9 | 110 | 0 | NT | <1/5 |
| 100 | NT | 4.5 | 3.7 | 97 | 0 | NT | <1/5 |
| 101 | NT | 10.3 | 3.8 | 97 | 7 | NT | <1/5 |
| 102 | NT | 2.7 | 3.9 | 102 | 1 | NT | <1/5 |
| 103 | NT | 0.1 | 4 | 74 | 0 | NT | <1/5 |
| 104 | NT | 3.6 | 4.1 | 107 | 0 | NT | <1/5 |
| 105 | NT | 4.3 | 4.6 | 73 | 6 | NT | <1/5 |
| 106 | NT | 8.9 | 3.9 | 33 | 10 | NT | <1/5 |
| 107 | NT | 1.9 | 5.3 | 85 | 7 | NT | <1/5 |
| 108 | NT | 10.7 | 4.9 | 127 | 0 | NT | <1/5 |
| 109 | NT | 5.7 | 4.5 | 86 | 3 | NT | <1/5 |
| 110 | NT | 4.6 | 5.2 | 151 | 0 | NT | <1/5 |
| 111 | NT | 3 | 4.3 | 127 | 0 | NT | <1/5 |
| 112 | NT | 2.1 | 4.4 | 17 | 1 | NT | <1/5 |
| 113 | NT | 0.9 | 4.5 | 394 | 1 | NT | <1/5 |
| 114 | NT | 0.0 | 4.3 | 26 | 3 | NT | <1/5 |
| 115 | NT | 0.1 | 5.6 | 532 | 0 | NT | <1/5 |
| 116 | NT | 12.4 | 4.8 | 143 | 2 | NT | <1/5 |
| 117 | NT | 5.0 | 4.2 | 87 | 7 | NT | <1/5 |
| 118 | NT | 4.3 | 4.6 | 68 | 2 | NT | <1/5 |
| Diagnostic test 2×2table | NT | Se: 86.15% (95%CI: 75.34–93.47) Sp: 100% (95%CI: 91.95–100) | Se: 89.39% (95%CI: 75.36–95.63) Sp: 100% (95%CI: 91.95–100) | Se: 100% (95%CI: 95–100) Sp: 100% (95%CI: 91.96–100) | Se: 100% (95%CI: 95–100) Sp: 100% (95%CI: 91.96–100) | NT | Se: 100% (95%CI: 94.87–100) Sp: 100% (95%CI: 91.96–100) |
| K cohen agreement vs. VNT | NT | K: 0.83 (95%CI: 0.73–0.93) | K: 0.87 (95%CI: 0.78–0.96) | K: 1 (95%: +/−0) | K: 1 (95%: +/−0) | NT |
| Name of Samples | |||
|---|---|---|---|
| GenBank accession number | 230860/12/20 (ISS81/21/3) Human 1-08-20 OL738656 | 240408/16/20 (ISS221/21/2) Mink 17-08-20 OL739154 | 350227/1/20 (ISS93/21/1) Mink 29-10-20 OL739160 |
| Sequence quality parameters | |||
| Coverage (cutoff > di 30×) | 515× | 1.072× | 2.360× |
| Genome length (Ref. Acc. N° NC_045512.2; 29,993 bp) | 29.826 | 29.612 | 29.835 |
| Number of total reads | 85.050 | 657.253 | 3.484.458 |
| Number of mapped reads | 83.850 (98.59%) | 222.056 (33.7%) | 392.713 (11.27%) |
| Spike Protein-Amino Acid Mutations | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Collection date | Number of sequences | L216H | S477N | N501Y | D614G | P681H | Y453F | Mink Cluster V | Pango lin | |
| Human sequence | 1 August 2020 | 1 | no | no | no | yes | no | no | no | B.1 |
| Mink sequence | 17 August 2020 | 1 | yes | no | yes | yes | yes | no | no | B.1.1 |
| Mink sequence | 29 October 2020 | 1 | no | yes | no | yes | no | no | no | B.1.160 |
| Human sequences from Italy available in GISAID | 2020 | 5.634 | 0% | 230 (4.1%) | 173 (3.1%) | 5.343 (94.8%) | 180 (3.2%) | no | no | |
| Human sequences from Italy available in GISAID | 2021 | 90.147 | 0% | 2.748 (3%) | 33.363 (37%) | 86.701 (96.2%) | 32.622 (36.2%) | 3 (0.003%) | no | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, A.; Lelli, D.; Trogu, T.; Lavazza, A.; Barbieri, I.; Boniotti, M.; Pezzoni, G.; Salogni, C.; Giovannini, S.; Alborali, G.; et al. SARS-CoV-2 in a Mink Farm in Italy: Case Description, Molecular and Serological Diagnosis by Comparing Different Tests. Viruses 2022, 14, 1738. https://doi.org/10.3390/v14081738
Moreno A, Lelli D, Trogu T, Lavazza A, Barbieri I, Boniotti M, Pezzoni G, Salogni C, Giovannini S, Alborali G, et al. SARS-CoV-2 in a Mink Farm in Italy: Case Description, Molecular and Serological Diagnosis by Comparing Different Tests. Viruses. 2022; 14(8):1738. https://doi.org/10.3390/v14081738
Chicago/Turabian StyleMoreno, Ana, Davide Lelli, Tiziana Trogu, Antonio Lavazza, Ilaria Barbieri, MariaBeatrice Boniotti, Giulia Pezzoni, Cristian Salogni, Stefano Giovannini, Giovanni Alborali, and et al. 2022. "SARS-CoV-2 in a Mink Farm in Italy: Case Description, Molecular and Serological Diagnosis by Comparing Different Tests" Viruses 14, no. 8: 1738. https://doi.org/10.3390/v14081738