
Lipidomics Revealed Plasma Phospholipid Profile Differences between Deceased and Recovered COVID-19 Patients
 ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Lipids and Quantification of Phospholipid Content
2.2. Phospholipid Profiling by Hydrophilic Interaction Liquid Chromatography Coupled with High-Resolution Tandem Mass Spectrometry (Hilic-Ms/MS)
2.3. Determination of COX-1 and COX-2 Activity
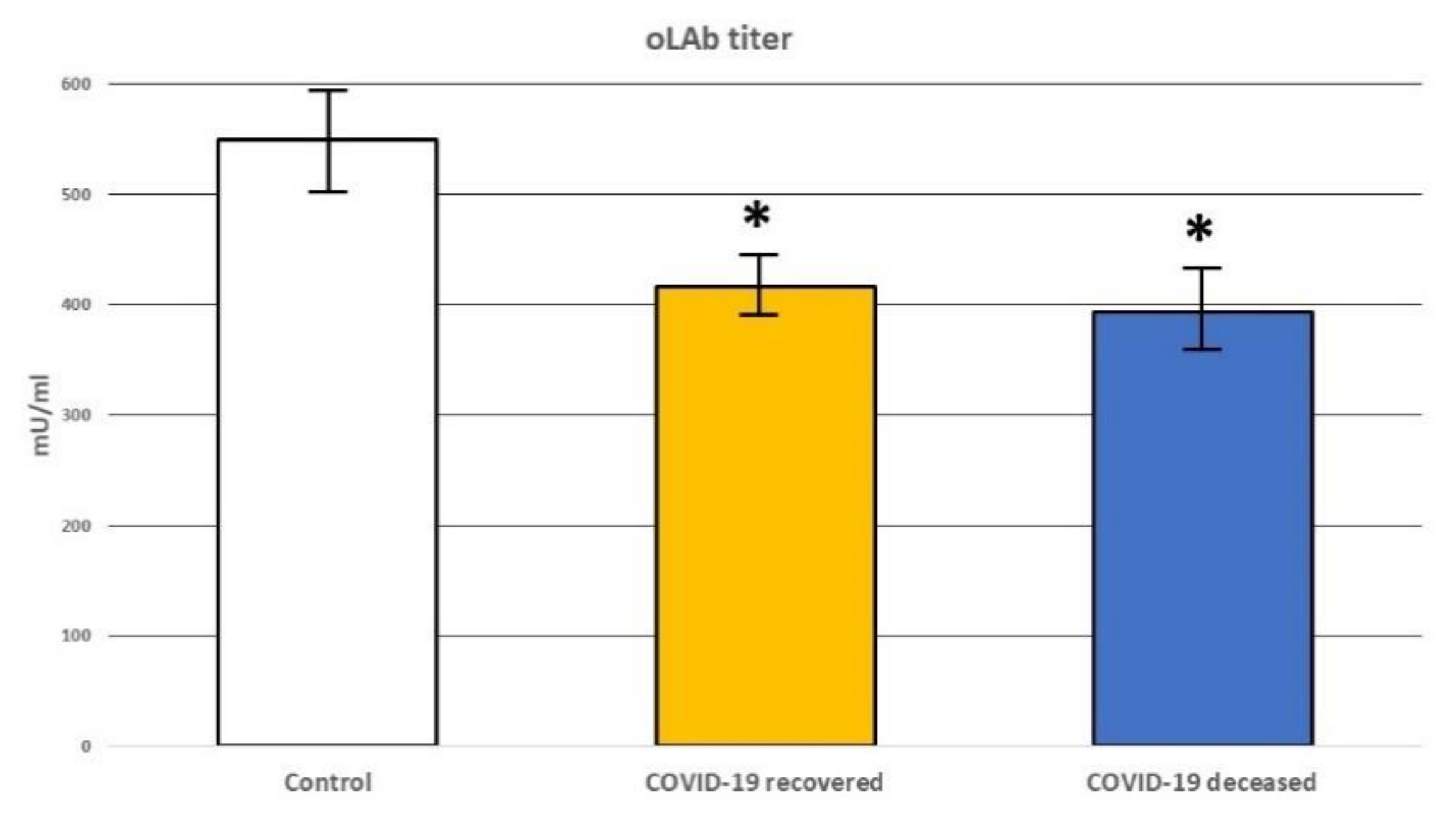
2.4. Determination of the Antibodies Directed against Oxidative Modifications of Low-Density Lipoprotein (Olab) Titer
2.5. Data Processing
2.6. Statistical Analysis
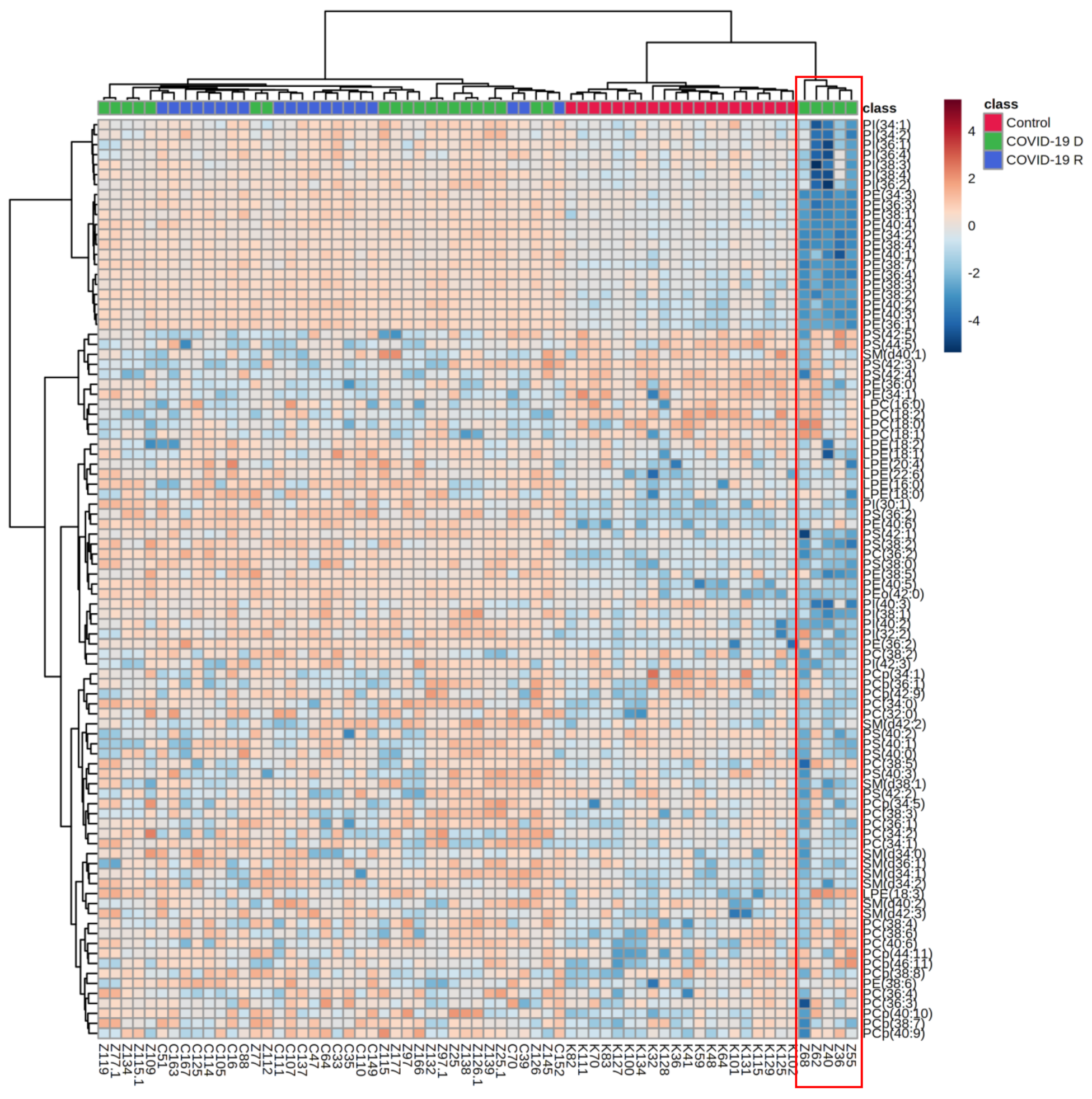
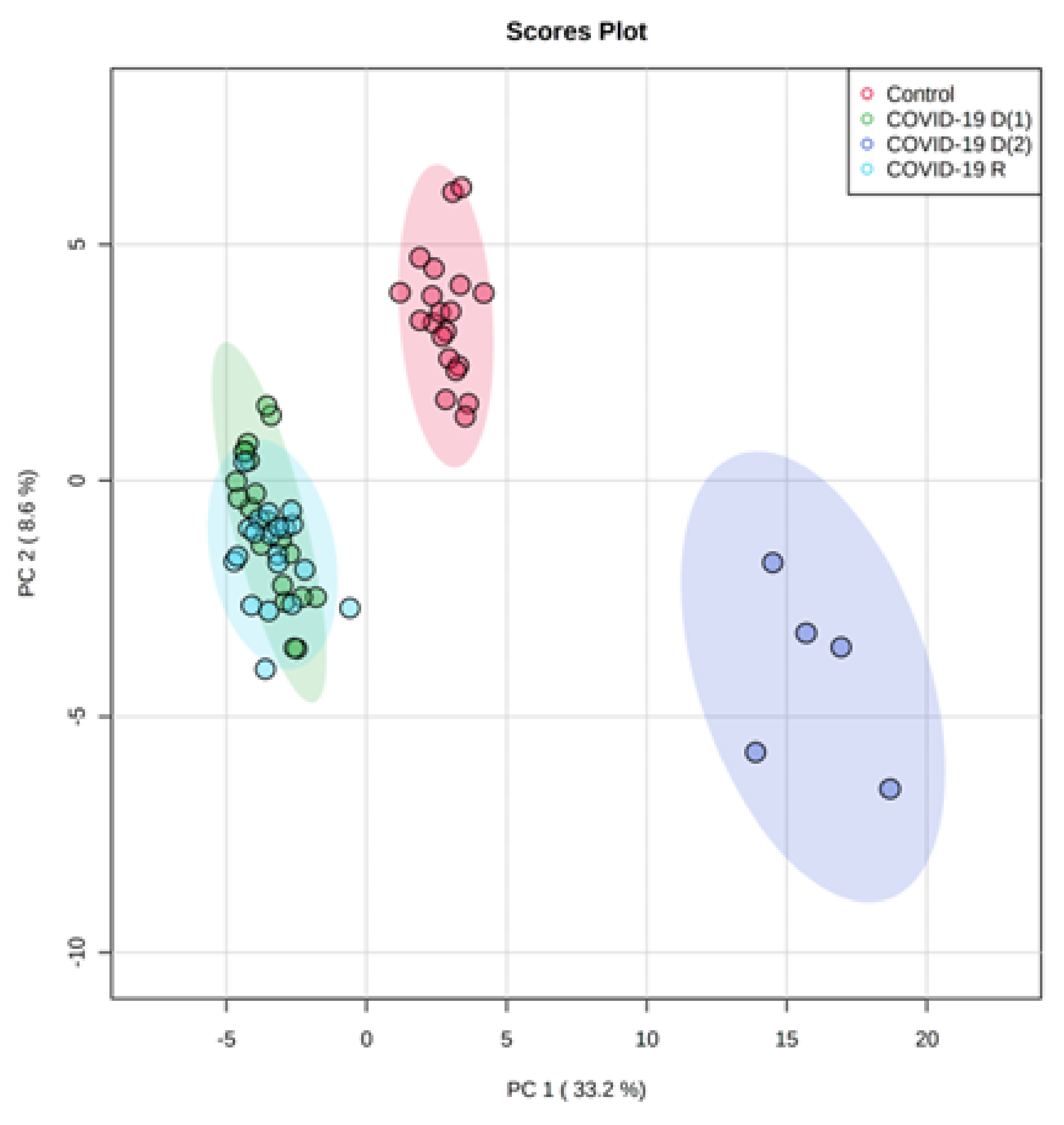
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuiken, T.; Fouchier, R.A.M.; Schutten, M.; Rimmelzwaan, G.F.; van Amerongen, G.; van Riel, D.; Laman, J.D.; de Jong, T.; van Doornum, G.; Lim, W.; et al. Newly Discovered Coronavirus as the Primary Cause of Severe Acute Respiratory Syndrome. Lancet 2003, 362, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int (accessed on 29 March 2022).
- Shi, H.; Han, X.; Zheng, C. Evolution of CT Manifestations in a Patient Recovered from 2019 Novel Coronavirus (2019-NCoV) Pneumonia in Wuhan, China. Radiology 2020, 295, 20. [Google Scholar] [CrossRef]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical Course and Outcomes of Critically Ill Patients with SARS-CoV-2 Pneumonia in Wuhan, China: A Single-Centered, Retrospective, Observational Study. Lancet. Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.A.; Hulett, M.D.; Poon, I.K. The Phospholipid Code: A Key Component of Dying Cell Recognition, Tumor Progression and Host–Microbe Interactions. Cell Death Differ. 2015, 22, 1893–1905. [Google Scholar] [CrossRef] [Green Version]
- Alagumuthu, M.; Dahiya, D.; Nigam, P.S.; Alagumuthu, M.; Dahiya, D.; Nigam, P.S. Phospholipid—The Dynamic Structure between Living and Non-Living World; a Much Obligatory Supramolecule for Present and Future. AIMSMOLES 2019, 6, 1–19. [Google Scholar] [CrossRef]
- Cai, T.; Yang, F. Phospholipid and Phospholipidomics in Health and Diseases. In Lipidomics in Health & Disease; Springer: Berlin/Heidelberg, Germany, 2018; pp. 177–202. [Google Scholar]
- Du, Y.; Mi, Z.; Xie, Y.; Lu, D.; Zheng, H.; Sun, H.; Zhang, M.; Niu, Y. Insights into the Molecular Basis of Tick-Borne Encephalitis from Multiplatform Metabolomics. PLoS Negl. Trop. Dis. 2021, 15, 0009172. [Google Scholar] [CrossRef]
- Eisfeld, A.J.; Halfmann, P.J.; Wendler, J.P.; Kyle, J.E.; Burnum-Johnson, K.E.; Peralta, Z.; Maemura, T.; Walters, K.B.; Watanabe, T.; Fukuyama, S.; et al. Multi-Platform ’Omics Analysis of Human Ebola Virus Disease Pathogenesis. Cell Host Microbe. 2017, 22, 817–829. [Google Scholar] [CrossRef] [Green Version]
- Kyle, J.E.; Burnum-Johnson, K.E.; Wendler, J.P.; Eisfeld, A.J.; Halfmann, P.J.; Watanabe, T.; Sahr, F.; Smith, R.D.; Kawaoka, Y.; Waters, K.M.; et al. Plasma Lipidome Reveals Critical Illness and Recovery from Human Ebola Virus Disease. Proc. Natl. Acad. Sci. U S A 2019, 116, 3919–3928. [Google Scholar] [CrossRef] [Green Version]
- Žarković, N.; Orehovec, B.; Milković, L.; Baršić, B.; Tatzber, F.; Wonisch, W.; Tarle, M.; Kmet, M.; Mataić, A.; Jakovčević, A.; et al. Preliminary Findings on the Association of the Lipid Peroxidation Product 4-Hydroxynonenal with the Lethal Outcome of Aggressive COVID-19. Antioxidants 2021, 10, 1341. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipides from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bartlett, E.M.; Lewis, D.H. Spectrophotometric Determination of Phosphate Esters in the Presence and Absence of Orthophosphate. Anal. Biochem. 1970, 36, 159–167. [Google Scholar] [CrossRef]
- Łuczaj, W.; Dobrzyńska, I.; Wroński, A.; Domingues, M.R.; Domingues, P.; Skrzydlewska, E. Cannabidiol-Mediated Changes to the Phospholipid Profile of UVB-Irradiated Keratinocytes from Psoriatic Patients. Int. J. Mol. Sci. 2020, 21, 6592. [Google Scholar] [CrossRef]
- Łuczaj, W.; Moniuszko-Malinowska, A.; Domingues, P.; Domingues, M.R.; Gindzienska-Sieskiewicz, E.; Skrzydlewska, E. Plasma Lipidomic Profile Signature of Rheumatoid Arthritis versus Lyme Arthritis Patients. Arch. Biochem. Biophys. 2018, 654, 105–114. [Google Scholar] [CrossRef]
- Kulmacz, R.J.; Wang, L.H. Comparison of Hydroperoxide Initiator Requirements for the Cyclooxygenase Activities of Prostaglandin H Synthase-1 and -2. J. Biol. Chem. 1995, 270, 24019–24023. [Google Scholar] [CrossRef] [Green Version]
- Nikolic-Heitzler, V.; Rabuzin, F.; Tatzber, F.; Vrkic, N.; Bulj, N.; Borovic, S.; Wonisch, W.; Sunko, B.M.; Zarkovic, N. Persistent Oxidative Stress after Myocardial Infarction Treated by Percutaneous Coronary Intervention. Tohoku J. Exp. Med. 2006, 210, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Reiger, J.; Tatzber, F.; Stettner, H.; Wonisch, W. Antibody against Low Density Lipoproteins (OLAb), Procalcitonin and Neopterin as Prognostic Marker during Sepsis and Systemic Inflammatory Response Syndrome (SIRS). Free Radic. Biol. Med. 2018, 124, 573–574. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Zarkovic, N.; Jakovcevic, A.; Mataic, A.; Jaganjac, M.; Vukovic, T.; Waeg, G.; Zarkovic, K. Post-Mortem Findings of Inflammatory Cells and the Association of 4-Hydroxynonenal with Systemic Vascular and Oxidative Stress in Lethal COVID-19. Cells 2022, 11, 444. [Google Scholar] [CrossRef]
- Arita, M. Mediator Lipidomics in Acute Inflammation and Resolution. J. Biochem. 2012, 152, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, K.; Yang, L.; Liu, R.; Chu, Y.; Qin, X.; Yang, P.; Yu, H. Lipid Metabolism in Inflammation-Related Diseases. Analyst 2018, 143, 4526–4536. [Google Scholar] [CrossRef]
- Wang, X.; Nijman, R.; Camuzeaux, S.; Sands, C.; Jackson, H.; Kaforou, M.; Emonts, M.; Herberg, J.A.; Maconochie, I.; Carrol, E.D.; et al. Plasma Lipid Profiles Discriminate Bacterial from Viral Infection in Febrile Children. Sci. Rep. 2019, 9, 17714. [Google Scholar] [CrossRef] [Green Version]
- Barberis, E.; Timo, S.; Amede, E.; Vanella, V.V.; Puricelli, C.; Cappellano, G.; Raineri, D.; Cittone, M.G.; Rizzi, E.; Pedrinelli, A.R.; et al. Large-Scale Plasma Analysis Revealed New Mechanisms and Molecules Associated with the Host Response to SARS-CoV-2. Int. J. Mol. Sci. 2020, 21, 8623. [Google Scholar] [CrossRef]
- Kaur, G.; Ji, X.; Rahman, I. SARS-CoV2 Infection Alters Tryptophan Catabolism and Phospholipid Metabolism. Metabolites 2021, 11, 659. [Google Scholar] [CrossRef]
- Ireland, R.; Schwarz, B.; Nardone, G.; Wehrly, T.D.; Broeckling, C.D.; Chiramel, A.I.; Best, S.M.; Bosio, C.M. Unique Francisella Phosphatidylethanolamine Acts as a Potent Anti-Inflammatory Lipid. J. Innate. Immun. 2018, 10, 291–305. [Google Scholar] [CrossRef]
- Mazidimoradi, A.; Alemzadeh, E.; Alemzadeh, E.; Salehiniya, H. The Effect of Polyunsaturated Fatty Acids on the Severity and Mortality of COVID Patients: A Systematic Review. Life Sci. 2022, 299, 120489. [Google Scholar] [CrossRef]
- Gallo, C.G.; Fiorino, S.; Posabella, G.; Antonacci, D.; Tropeano, A.; Pausini, E.; Pausini, C.; Guarniero, T.; Hong, W.; Giampieri, E.; et al. The Function of Specialized Pro-Resolving Endogenous Lipid Mediators, Vitamins, and Other Micronutrients in the Control of the Inflammatory Processes: Possible Role in Patients with SARS-CoV-2 Related Infection. Prostaglandins Other Lipid Mediat. 2022, 159, 106619. [Google Scholar] [CrossRef]
- Liu, W.; Porter, N.A.; Schneider, C.; Brash, A.R.; Yin, H. Formation of 4-hydroxynonenal from cardiolipin oxidation: Intramolecular peroxyl radical addition and decomposition. Free Radic. Biol. Med. 2011, 50, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.; Tallman, K.A.; Porter, N.A.; Brash, A.R. Two Distinct Pathways of Formation of 4-Hydroxynonenal: Mechanisms of nonenzymatic transformation of the 9- and 13-hydroperoxides of linoleic acid to 4-hydroxyalkenals*. J. Bio. Chem. 2001, 276, 20831–20838. [Google Scholar] [CrossRef]
- Łuczaj, W.; Gęgotek, A.; Skrzydlewska, E. Antioxidants and HNE in Redox Homeostasis. Free Radic. Biol. Med. 2017, 111, 87–101. [Google Scholar] [CrossRef]
- Shoeb, M.; Ansari, N.H.; Srivastava, S.K.; Ramana, K.V. 4-Hydroxynonenal in the Pathogenesis and Progression of Human Diseases. Curr. Med. Chem. 2014, 21, 230–237. [Google Scholar] [CrossRef]
- Kumagai, T.; Usami, H.; Matsukawa, N.; Nakashima, F.; Chikazawa, M.; Shibata, T.; Noguchi, N.; Uchida, K. Functional Interaction between Cyclooxygenase-2 and P53 in Response to an Endogenous Electrophile. Redox. Biol. 2014, 4, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Rouzer, C.A.; Marnett, L.J. Cyclooxygenases: Structural and Functional Insights. J. Lipid. Res. 2009, 50, S29–S34. [Google Scholar] [CrossRef] [Green Version]
- Argañaraz, G.A.; Palmeira, J.d.F.; Argañaraz, E.R. Phosphatidylserine inside out: A Possible Underlying Mechanism in the Inflammation and Coagulation Abnormalities of COVID-19. Cell Commun. Signal. 2020, 18, 190. [Google Scholar] [CrossRef]
- Zhang, L.; Richard, A.S.; Jackson, C.B.; Ojha, A.; Choe, H. Phosphatidylethanolamine and Phosphatidylserine Synergize to Enhance GAS6/AXL-Mediated Virus Infection and Efferocytosis. J. Virol. 2020, 95, e02079-20. [Google Scholar] [CrossRef]
- Meertens, L.; Carnec, X.; Lecoin, M.P.; Ramdasi, R.; Guivel-Benhassine, F.; Lew, E.; Lemke, G.; Schwartz, O.; Amara, A. The TIM and TAM Families of Phosphatidylserine Receptors Mediate Dengue Virus Entry. Cell Host Microbe 2012, 12, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Moller-Tank, S.; Kondratowicz, A.S.; Davey, R.A.; Rennert, P.D.; Maury, W. Role of the Phosphatidylserine Receptor TIM-1 in Enveloped-Virus Entry. J. Virol. 2013, 87, 8327–8341. [Google Scholar] [CrossRef] [Green Version]
- Stark, K.; Massberg, S. Interplay between Inflammation and Thrombosis in Cardiovascular Pathology. Nat. Rev. Cardiol. 2021, 18, 666–682. [Google Scholar] [CrossRef]
- Poggiali, E.; Zaino, D.; Immovilli, P.; Rovero, L.; Losi, G.; Dacrema, A.; Nuccetelli, M.; Vadacca, G.B.; Guidetti, D.; Vercelli, A.; et al. Lactate Dehydrogenase and C-Reactive Protein as Predictors of Respiratory Failure in COVID-19 Patients. Clin. Chim. Acta. 2020, 509, 135–138. [Google Scholar] [CrossRef]
- AbouYabis, A.N.; Bell, G.T. Hemolytic Anemia Complicating COVID-19 Infection. J. Hematol. 2021, 10, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Nair, L.J.; Regukumar, A.; Baalamurugan, K.T. COVID-19-Associated Severe Autoimmune Hemolytic Anemia: A Rare Case Report. Saudi. J. Med. Med. Sci. 2021, 9, 276–279. [Google Scholar] [CrossRef] [PubMed]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.; Eichbaum, Q. COVID-19 Associated with Severe Autoimmune Hemolytic Anemia. Transfusion 2021, 61, 635–640. [Google Scholar] [CrossRef]
- Lazarian, G.; Quinquenel, A.; Bellal, M.; Siavellis, J.; Jacquy, C.; Re, D.; Merabet, F.; Mekinian, A.; Braun, T.; Damaj, G.; et al. Autoimmune Haemolytic Anaemia Associated with COVID-19 Infection. Br. J. Haematol. 2020, 190, 29–31. [Google Scholar] [CrossRef]
- Lopez, C.; Kim, J.; Pandey, A.; Huang, T.; DeLoughery, T.G. Simultaneous Onset of COVID-19 and Autoimmune Haemolytic Anaemia. Br. J. Haematol. 2020, 190, 31–32. [Google Scholar] [CrossRef]
- Wahlster, L.; Weichert-Leahey, N.; Trissal, M.; Grace, R.F.; Sankaran, V.G. COVID-19 Presenting with Autoimmune Hemolytic Anemia in the Setting of Underlying Immune Dysregulation. Pediatr. Blood Cancer 2020, 67, e28382. [Google Scholar] [CrossRef]
- Barcellini, W.; Fattizzo, B. Clinical Applications of Hemolytic Markers in the Differential Diagnosis and Management of Hemolytic Anemia. Dis. Markers 2015, 2015, 635670. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participants | Number of Participants | Sex (Female/Male) | Age Range (Average) | CRP [mg/L] | LDH [U/L] | Oxygen Saturation [%] |
|---|---|---|---|---|---|---|
| Control [healthy subjects] | 20 | 12/8 | 33−56 (44) | 0.00−5.00 | 140−280 | > 95% |
| COVID-19 D [COVID-19 deceased patients] | 20 | 12/8 | 58−85 (73) | 132.3 ± 54.8 | 488.4−156.5 | 82.35 ± 14.05 |
| COVID-19 R [COVID-19 recovered patients] | 20 | 12/8 | 54−85 (65) | 114.5 ± 61.8 | 355.0 ± 147.5 | 90.21 ± 6.05 |
| Participants | Number of Participants | Sex (Female/Male) | Age Range (Average) | CRP [mg/L] | LDH [U/L] | Oxygen Saturation [%] |
|---|---|---|---|---|---|---|
| COVID-19 D1 [COVID-19 deceased patients subgroup 1] | 15 | 9/6 | 58–85 (72) | 168.2 ± 73.1 | 300.3 ± 145.9 | 81.4 ± 15.5 |
| COVID-19 D2 [COVID-19 deceased patients subgroup 2] | 5 | 3/2 | 64–80 (74) | 96.4 ± 36.6 | 676.5 ± 167.0 | 83.3 ± 12.6 |
| PL Class | PL Specie | Phospholipid Molecular Species | Log2 (Fold-Change) | |||||
|---|---|---|---|---|---|---|---|---|
| COVID-19 R vs. Control | COVID-19 D(1) vs. Control | COVID-19 D(2) vs. Control | COVID-19 D(2) vs. COVID-19 D(1) | COVID-19 D(1) vs. COVID-19 R | COVID-19 D(2) vs. COVID-19 R | |||
| PS | PS(36:2) | PS(18:0/18:2) | 5.18 ↑ | 4.27 ↑ | - | 3.52 ↓ | - | 4.43 ↓ |
| PE | PE(40:6) | PE(18:0/22:6) | 3.64 ↑ | 3.29 ↑ | 2.46 ↑ | - | - | 1.19 ↓ |
| PC | PC(36:2) | PC(18:0/18:2) | 3.06 ↑ | 2.67 ↑ | 2.70 ↓ | 5.37 ↓ | - | 5.75 ↓ |
| PE | PE(40:5) | PE(18:0/22:5) | 3.15 ↑ | 3.22 ↑ | - | - | - | 4.02 ↓ |
| PS | PS(38:2) | PS(20:0/18:2) | 1.28 ↑ | 1.31 ↑ | 2.14 ↓ | 3.45 ↓ | - | 3.42 ↓ |
| PE | PE(36:4) | PE(16:0/20:4) | 1.71 ↑ | 1.71 ↑ | 8.16 ↓ | 9.86 ↓ | - | 9.87 ↓ |
| PE | PE(38:3) | PE(20:0/18:3) | 1.79 ↑ | 1.63 ↑ | 6.65 ↓ | 8.28 ↓ | - | 8.44 ↓ |
| PE | PE(38:2) | PE(20:0/18:2) | 1.83 ↑ | 1.74 ↑ | 5.67 ↓ | 7.41 ↓ | - | 7.49 ↓ |
| PE | PE(40:2) | PE(18:0/22:2) | 2.42 ↑ | 2.38 ↑ | 5.11 ↓ | 7.49 ↓ | - | 7.53 ↓ |
| PE | PE(40:3) | PE(20:1/18:2) | 1.54 ↑ | 2.92 ↑ | 7.96 ↓ | 10.88 ↓ | - | 11.09 ↓ |
| PE | PE(36:1) | PE(18:0/18:1) | 3.57 ↑ | 3.24 ↑ | 7.61 ↓ | 10.85 ↓ | - | 11.18 ↓ |
| PI | PI(36:2) | PI(18:0/18:2) | - | 1.29 ↑ | 2.45 ↓ | 3.74 ↓ | - | 3.42 ↓ |
| PE | PE(34:3) | PE(16:1/18:2) | 1.45 ↑ | 1.02 ↑ | 7.48 ↓ | 8.50 ↓ | - | 8.93 ↓ |
| PE | PE(36:3) | PE(18:1/18:2) | 1.32 ↑ | 1.27 ↑ | 6.66 ↓ | 7.93 ↓ | - | 7.98 ↓ |
| PE | PE(38:1) | PE(20:0/18:1) | 1.34 ↑ | 1.11 ↑ | 5.51 ↓ | 6.62 ↓ | - | 6.85 ↓ |
| PE | PE(40:4) | PE(20:0/20:4) | 2.32 ↑ | 2.30 ↑ | 8.39 ↓ | 10.69 ↓ | - | 10.71 ↓ |
| PE | PE(34:2) | PE(16:0/18:2) | 1.44 ↑ | 1.86 ↑ | 8.86 ↓ | 10.72 ↓ | - | 10.30 ↓ |
| PE | PE(38:4) | PE(18:0/20:4) | 1.43 ↑ | 1.77 ↑ | 9.39 ↓ | 11.17 ↓ | - | 10.82 ↓ |
| PE | PE(38:7) | PE(16:1/22:6) | 1.27 ↑ | 1.31 ↑ | 5.67 ↓ | 6.98 ↓ | - | 6.95 ↓ |
| PE | PE(40:1) | PE(18:0/22:1) | 1.71 ↑ | 1.55 ↑ | 5.46 ↓ | 7.04 ↓ | - | 7.16 ↓ |
| Groups | COX 1 [nmol/min/mL] | COX 2 [nmol/min/mL] |
|---|---|---|
| Control | 14.59 ± 4.99 | 9.90 ± 1.90 |
| COVID-19 D1 | 18.42 ± 3.61 ab | 14.23 ± 1.12 b |
| COVID-19 D2 | 14.74 ± 2.74 | 5.71 ± 0.46 abc |
| COVID-19 R | 24.52 ± 3.22 a | 19.53 ± 4.14 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žarković, N.; Orehovec, B.; Baršić, B.; Tarle, M.; Kmet, M.; Lukšić, I.; Tatzber, F.; Wonisch, W.; Skrzydlewska, E.; Łuczaj, W. Lipidomics Revealed Plasma Phospholipid Profile Differences between Deceased and Recovered COVID-19 Patients. Biomolecules 2022, 12, 1488. https://doi.org/10.3390/biom12101488
Žarković N, Orehovec B, Baršić B, Tarle M, Kmet M, Lukšić I, Tatzber F, Wonisch W, Skrzydlewska E, Łuczaj W. Lipidomics Revealed Plasma Phospholipid Profile Differences between Deceased and Recovered COVID-19 Patients. Biomolecules. 2022; 12(10):1488. https://doi.org/10.3390/biom12101488
Chicago/Turabian StyleŽarković, Neven, Biserka Orehovec, Bruno Baršić, Marko Tarle, Marta Kmet, Ivica Lukšić, Franz Tatzber, Willibald Wonisch, Elżbieta Skrzydlewska, and Wojciech Łuczaj. 2022. "Lipidomics Revealed Plasma Phospholipid Profile Differences between Deceased and Recovered COVID-19 Patients" Biomolecules 12, no. 10: 1488. https://doi.org/10.3390/biom12101488