Author Contributions
Conceptualization, C.B.J.; Formal analyses, C.B.J., Y.X., P.V., D.Y., Y.K. and Y.R.; Investigation, Y.R., Y.X., D.Y., L.Z., J.S., P.V., Y.K., J.P. and E.H.; Writing, Original Draft; C.B.J., J.S., Y.R., C.B.S.-S., S.T. and P.V.; Writing, Reviewing and Editing, C.B.J., Y.R., C.B.S.-S., P.V., S.T., Y.X. and E.F.; Visualization, D.Y., Y.K., Y.X., Y.R. and C.B.J.; Resources, C.B.J.; Project Administration, C.B.J. All authors have read and agreed to the published version of the manuscript.
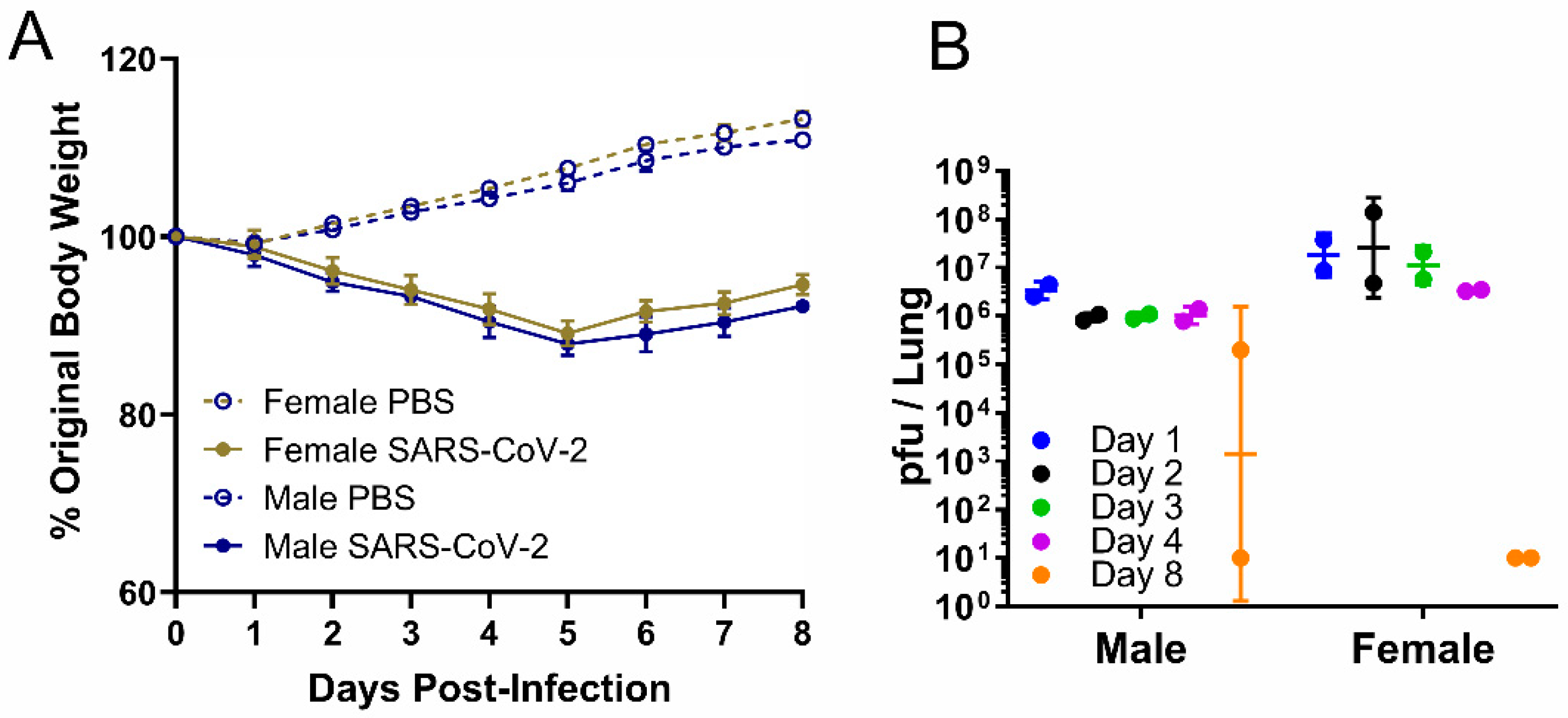
Figure 1.
Weight and viral load in male or female hamsters intranasally infected with PBS or SARS-CoV-2 at a high dose, 2 × 105 pfu. In the serial sacrifice study of SARS-CoV-2-infected hamsters, (A) body weight change (data are presented as mean ± SEM) was measured daily from 0 to 8 dpi, and (B) viral load was measured by plaque assay in lungs of male and female hamsters at five time points (n = 2). Data are presented as geometric mean ± geometric SD.
Figure 1.
Weight and viral load in male or female hamsters intranasally infected with PBS or SARS-CoV-2 at a high dose, 2 × 105 pfu. In the serial sacrifice study of SARS-CoV-2-infected hamsters, (A) body weight change (data are presented as mean ± SEM) was measured daily from 0 to 8 dpi, and (B) viral load was measured by plaque assay in lungs of male and female hamsters at five time points (n = 2). Data are presented as geometric mean ± geometric SD.
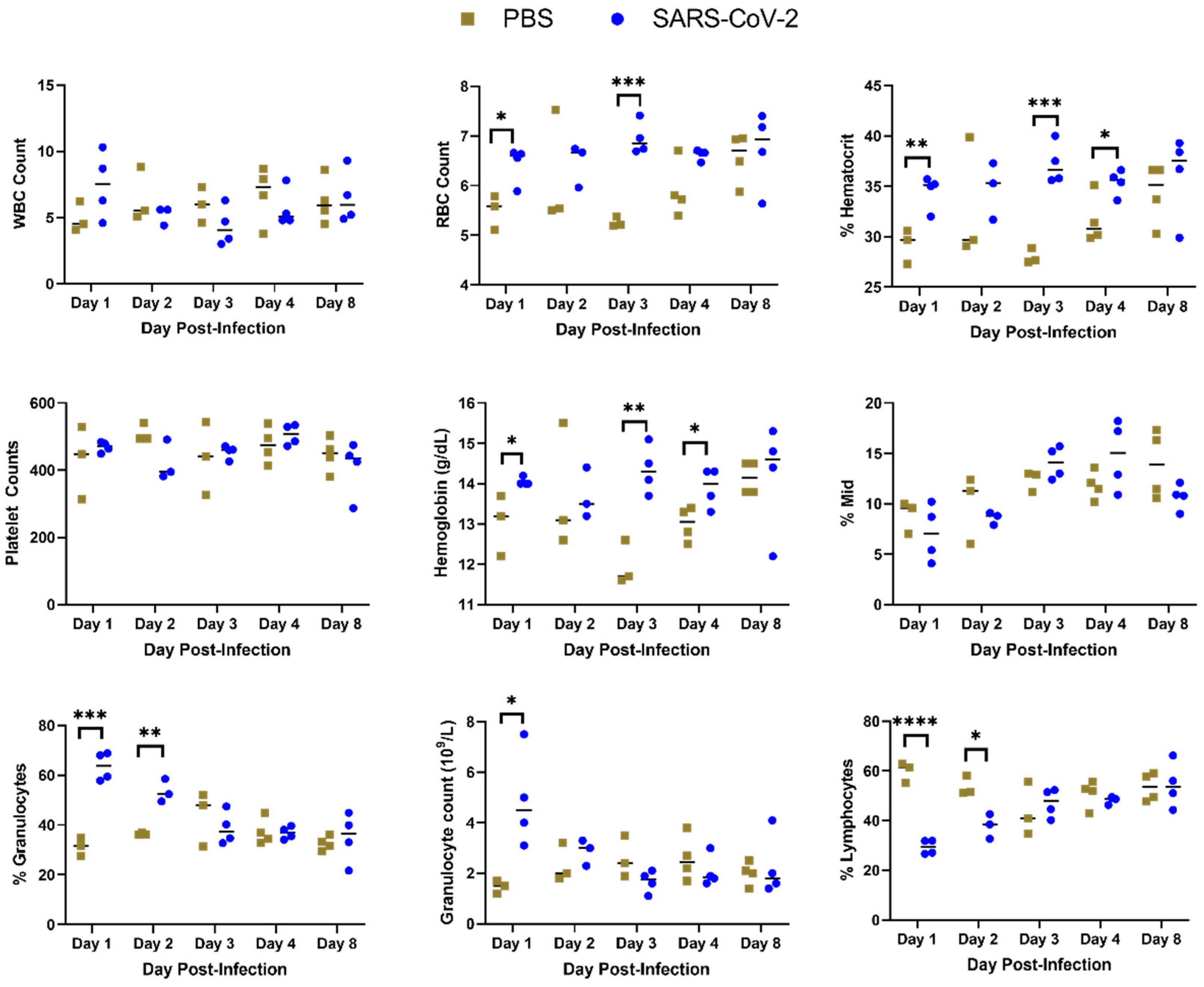
Figure 2.
Complete blood count from hamsters infected with SARS-CoV-2 or sham inoculated (PBS). Blood was drawn from hamsters on 1, 2, 3, 4 or 8 dpi and a complete blood count was performed. (*: p < 0.05, **: p < 0.01; ***: p < 0.001, ****: p < 0.0001.)
Figure 2.
Complete blood count from hamsters infected with SARS-CoV-2 or sham inoculated (PBS). Blood was drawn from hamsters on 1, 2, 3, 4 or 8 dpi and a complete blood count was performed. (*: p < 0.05, **: p < 0.01; ***: p < 0.001, ****: p < 0.0001.)
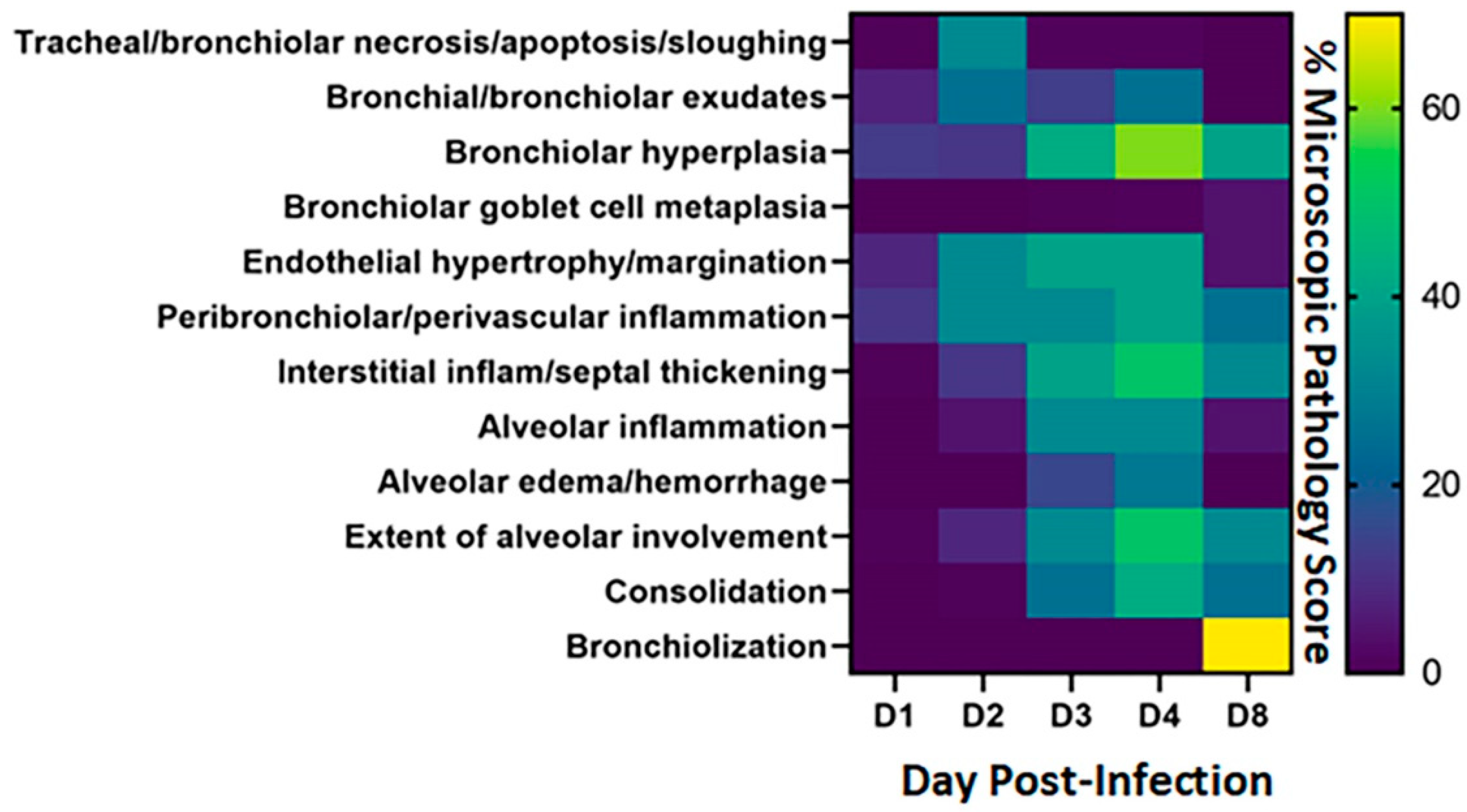
Figure 3.
Heatmap of microscopic pathology of Syrian hamster lungs with SARS-CoV-2. The severity scores for each category of microscopic pathology are illustrated using a heatmap. For each of the days examined, each box represents the average score from four hamsters for scores for these categories: tracheal/bronchiolar necrosis/apoptosis/sloughing, bronchial/bronchiolar exudates, bronchiolar hyperplasia, bronchiolar goblet cell metaplasia, endothelial hypertrophy/margination, peribronchiolar/perivascular inflammation, interstitial inflammation-septal thickening, alveolar inflammation, alveolar edema/hemorrhage, extent of alveolar involvement, consolidation and bronchiolization. Semi-quantitative scores used 0–1 = within normal limits, 2–14 = Minimal: Rare or inconspicuous lesions; 15–39 = Mild: Multifocal or small, focal, or widely separated, but conspicuous lesions; 40–79 = Moderate: Multifocal, prominent lesions; 80–99 = Marked: Extensive to coalescing lesions or areas of inflammation with some loss of structure and 100 = Severe: Diffuse lesion with effacement of normal structure. The scale to the right of the heat map reflects the scoring ranges from 0 to 70%.
Figure 3.
Heatmap of microscopic pathology of Syrian hamster lungs with SARS-CoV-2. The severity scores for each category of microscopic pathology are illustrated using a heatmap. For each of the days examined, each box represents the average score from four hamsters for scores for these categories: tracheal/bronchiolar necrosis/apoptosis/sloughing, bronchial/bronchiolar exudates, bronchiolar hyperplasia, bronchiolar goblet cell metaplasia, endothelial hypertrophy/margination, peribronchiolar/perivascular inflammation, interstitial inflammation-septal thickening, alveolar inflammation, alveolar edema/hemorrhage, extent of alveolar involvement, consolidation and bronchiolization. Semi-quantitative scores used 0–1 = within normal limits, 2–14 = Minimal: Rare or inconspicuous lesions; 15–39 = Mild: Multifocal or small, focal, or widely separated, but conspicuous lesions; 40–79 = Moderate: Multifocal, prominent lesions; 80–99 = Marked: Extensive to coalescing lesions or areas of inflammation with some loss of structure and 100 = Severe: Diffuse lesion with effacement of normal structure. The scale to the right of the heat map reflects the scoring ranges from 0 to 70%.
![Viruses 14 01403 g003]()
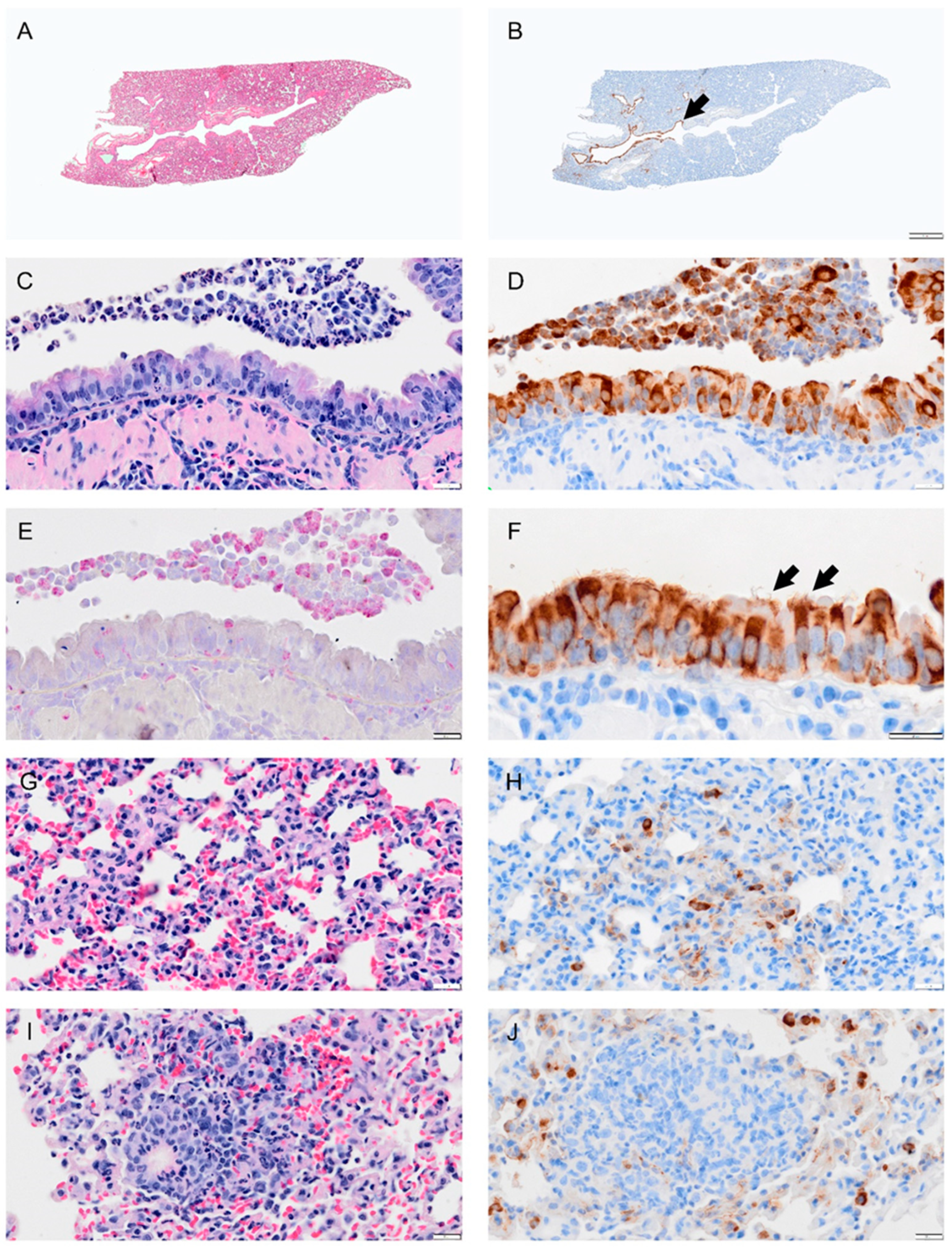
Figure 4.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 1 dpi with SARS-CoV-2. (A) Lower magnification of a H&E-stained section showed no marked change at 1 dpi. (B) Immunohistochemistry (IHC) of SARS-CoV-2 in lung section shows distribution of the virus in the bronchial epithelium of primary and secondary (segmental) bronchi (brown color arrowhead) and in some scattered areas of alveolar tissues. (C) H&E-stained section of bronchus showing inflammatory cell infiltration in bronchial wall (mucosa and submucosa) and cell debris (neutrophils, macrophages, necrotic epithelial cells) in lumen. (D) Adjacent serial sections of panel (C) show abundant infected epithelial cells in mucosa and debris. (E) Adjacent serial section with NACE stain shows neutrophils in debris, epithelium, and submucosal layer. (F) Higher magnification of virus IHC shows disorganized bronchial epithelial cells with disorganized cilia, see arrow. (G) H&E-stained section of alveolar tissue showing intravascular neutrophils and congestion. (H) Adjacent section of panel (G) stained for virus by IHC shows virus antigen concentrated in type 2 alveolar cells, with small amounts lining alveolar surfaces. (I) Section showing inflammatory cells in areas of alveolar collapse, representing early consolidation. (J) Viral antigen concentrated within type 2 pneumocytes surrounding consolidation area. Bar: (A,B) = 1 mm; (C–J) = 20 µm.
Figure 4.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 1 dpi with SARS-CoV-2. (A) Lower magnification of a H&E-stained section showed no marked change at 1 dpi. (B) Immunohistochemistry (IHC) of SARS-CoV-2 in lung section shows distribution of the virus in the bronchial epithelium of primary and secondary (segmental) bronchi (brown color arrowhead) and in some scattered areas of alveolar tissues. (C) H&E-stained section of bronchus showing inflammatory cell infiltration in bronchial wall (mucosa and submucosa) and cell debris (neutrophils, macrophages, necrotic epithelial cells) in lumen. (D) Adjacent serial sections of panel (C) show abundant infected epithelial cells in mucosa and debris. (E) Adjacent serial section with NACE stain shows neutrophils in debris, epithelium, and submucosal layer. (F) Higher magnification of virus IHC shows disorganized bronchial epithelial cells with disorganized cilia, see arrow. (G) H&E-stained section of alveolar tissue showing intravascular neutrophils and congestion. (H) Adjacent section of panel (G) stained for virus by IHC shows virus antigen concentrated in type 2 alveolar cells, with small amounts lining alveolar surfaces. (I) Section showing inflammatory cells in areas of alveolar collapse, representing early consolidation. (J) Viral antigen concentrated within type 2 pneumocytes surrounding consolidation area. Bar: (A,B) = 1 mm; (C–J) = 20 µm.
![Viruses 14 01403 g004]()
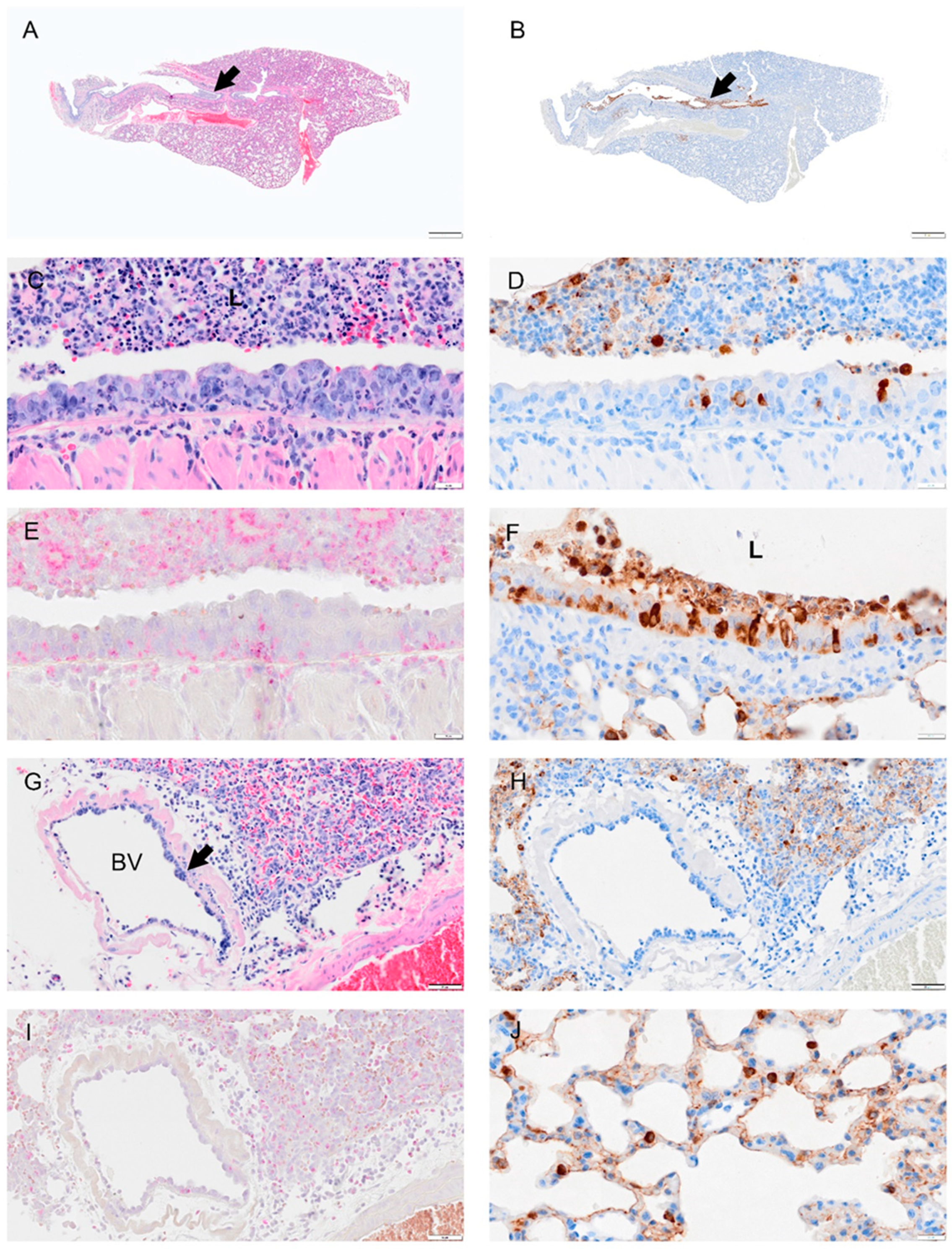
Figure 5.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 2 dpi with SARS-CoV-2. (A) A low magnification image shows widespread septal thickening and debris in the primary bronchial lumen (arrow). (B) Image of serial section adjacent to (A) showing viral antigen bronchial epithelium and debris. (C) Section showing the bronchus with abundant cell debris (neutrophils, monocytes, necrotic epithelial cells, pyknosis and karyorrhexis, red blood cells) in the bronchial lumen (L). Image also shows damaged epithelial cells (loss of cilia, disruption of cell layer and inflammatory cells in mucosal and submucosal layer. (D) Serial section adjacent to (C) showing scattered viral antigen positive cells in the epithelial layer and in lumenal debris. (E) NACE staining of bronchus which shows many infiltrating neutrophils. (F) Viral antigen in the bronchus, debris of lumen (L) and type 2 alveolar cells. (G) A blood vessel (BV) showed thickened hypercellular endothelium(arrow) and inflammatory cell infiltrates in the vascular tunica media, interstitium, and alveoli. (H) Serial section adjacent to (G) shows viral antigen in the alveolar cells but not in the blood vessel. (I) NACE staining of the blood vessel and surrounding tissues showing neutrophil infiltration. (J) Section shows viral antigen in type 2 alveolar cells and thickened septa. Bar: (A,B) = 1 mm; (C–F) = 20 µm; (G–I) = 50 µm and (J) = 20 µm.
Figure 5.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 2 dpi with SARS-CoV-2. (A) A low magnification image shows widespread septal thickening and debris in the primary bronchial lumen (arrow). (B) Image of serial section adjacent to (A) showing viral antigen bronchial epithelium and debris. (C) Section showing the bronchus with abundant cell debris (neutrophils, monocytes, necrotic epithelial cells, pyknosis and karyorrhexis, red blood cells) in the bronchial lumen (L). Image also shows damaged epithelial cells (loss of cilia, disruption of cell layer and inflammatory cells in mucosal and submucosal layer. (D) Serial section adjacent to (C) showing scattered viral antigen positive cells in the epithelial layer and in lumenal debris. (E) NACE staining of bronchus which shows many infiltrating neutrophils. (F) Viral antigen in the bronchus, debris of lumen (L) and type 2 alveolar cells. (G) A blood vessel (BV) showed thickened hypercellular endothelium(arrow) and inflammatory cell infiltrates in the vascular tunica media, interstitium, and alveoli. (H) Serial section adjacent to (G) shows viral antigen in the alveolar cells but not in the blood vessel. (I) NACE staining of the blood vessel and surrounding tissues showing neutrophil infiltration. (J) Section shows viral antigen in type 2 alveolar cells and thickened septa. Bar: (A,B) = 1 mm; (C–F) = 20 µm; (G–I) = 50 µm and (J) = 20 µm.
![Viruses 14 01403 g005]()
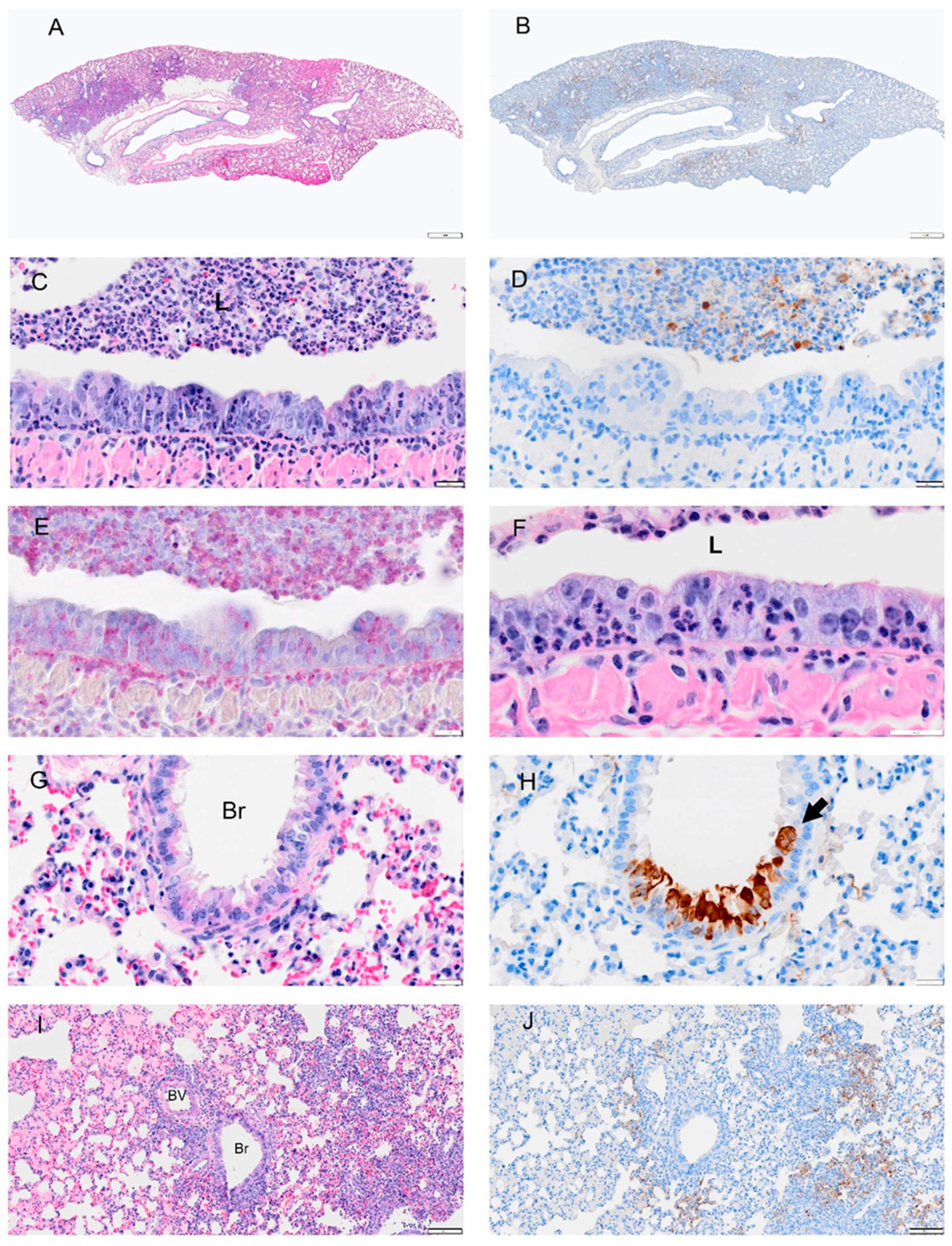
Figure 6.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 3 dpi with SARS-CoV-2. (A) Low magnification of a section with multifocal areas consolidation and hemorrhage. (B) An adjacent serial section of (A) shows viral antigen is concentrated at the margin of consolidated areas. (C) Higher magnification of bronchus shows debris with necrotic neutrophils, epithelial cells, and macrophages in bronchial lumen (L), abundant neutrophils within epithelial layer and submucosa, hyperplastic epithelial cells, and decreased cilia. (D) Adjacent serial section of (C) showing absence of viral antigen in the bronchial epithelium and but some remaining in luminal debris. (E) An adjacent serial section of (C,D) shows abundant neutrophils in the luminal debris, and within the epithelial and submucosal layers (visualized using NACE stain). (F) High magnification of the bronchial epithelium shows damaged mucosal epithelium with absent or very short cilia; neutrophils were the primary type of inflammatory cells. (G) H&E-stained section shows degenerated epithelial cells in small airways (Br). (H) An adjacent serial section to (G) shows viral antigen in damaged epithelial cells (arrow). (I) H&E-stained section shows hyperplasia of bronchiolar epithelium (Br), intra-alveolar inflammatory cell infiltrates (right side of image), and serous exudates filling alveolar spaces (left side of image). (J) Adjacent serial section of (I) shows viral antigen tends to be concentrated at the periphery of consolidated areas. Scale bars: (A,B) = 1 mm; (C–H) = 20 µm; (I,J) = 100 µm.
Figure 6.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 3 dpi with SARS-CoV-2. (A) Low magnification of a section with multifocal areas consolidation and hemorrhage. (B) An adjacent serial section of (A) shows viral antigen is concentrated at the margin of consolidated areas. (C) Higher magnification of bronchus shows debris with necrotic neutrophils, epithelial cells, and macrophages in bronchial lumen (L), abundant neutrophils within epithelial layer and submucosa, hyperplastic epithelial cells, and decreased cilia. (D) Adjacent serial section of (C) showing absence of viral antigen in the bronchial epithelium and but some remaining in luminal debris. (E) An adjacent serial section of (C,D) shows abundant neutrophils in the luminal debris, and within the epithelial and submucosal layers (visualized using NACE stain). (F) High magnification of the bronchial epithelium shows damaged mucosal epithelium with absent or very short cilia; neutrophils were the primary type of inflammatory cells. (G) H&E-stained section shows degenerated epithelial cells in small airways (Br). (H) An adjacent serial section to (G) shows viral antigen in damaged epithelial cells (arrow). (I) H&E-stained section shows hyperplasia of bronchiolar epithelium (Br), intra-alveolar inflammatory cell infiltrates (right side of image), and serous exudates filling alveolar spaces (left side of image). (J) Adjacent serial section of (I) shows viral antigen tends to be concentrated at the periphery of consolidated areas. Scale bars: (A,B) = 1 mm; (C–H) = 20 µm; (I,J) = 100 µm.
![Viruses 14 01403 g006]()
Figure 7.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at day 4 post-infection with SARS-CoV-2. (A) Section of the lung shows more extensive consolidation. (B) Image showing overall reduction in viral antigen (not present in bronchi). (C) Higher magnification of bronchus shows decreased debris in bronchial lumen, abundant neutrophil infiltration in hyperplastic (and disorganized) epithelium and submucosa. (D) A serial section adjacent to (C) shows minimal viral antigen in epithelial layer and debris. (E) An adjacent serial section to (C,D), shows neutrophils in debris, epithelial and sub mucosa layer following NACE staining. (F) Virus antigen is still present in some bronchiolar epithelial cells. (G) H&E-stained section which shows hyperplastic bronchial epithelium with neutrophil infiltration, debris in lumen, necrosis (Br) and inflammatory cells infiltrates in lung parenchyma, a blood vessel with thickened endothelium (BV) and perivascular edema. (H) An adjacent serial section to (G) shows small pockets of viral antigen in pulmonary parenchyma (consolidation) but not in bronchial epithelium. (I) H&E-stained lung tissue shows perivascular (BV) and peri-bronchial inflammatory cell infiltration, alveolar exudates, and alveolar epithelial hyperplasia with mitosis. (J) A BV with proliferated endothelial cells Monocytes and eosinophils, and fibroblasts (undergoing mitosis) are distributed around blood vessels. Necrotic lung tissues show damaged alveolar walls and inflammatory cell infiltrates in consolidation area. Bar: (A,B) = 1 mm; (C–F) = 20 µm; (G,H,J) = 100 µm; (I) = 50 µm.
Figure 7.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at day 4 post-infection with SARS-CoV-2. (A) Section of the lung shows more extensive consolidation. (B) Image showing overall reduction in viral antigen (not present in bronchi). (C) Higher magnification of bronchus shows decreased debris in bronchial lumen, abundant neutrophil infiltration in hyperplastic (and disorganized) epithelium and submucosa. (D) A serial section adjacent to (C) shows minimal viral antigen in epithelial layer and debris. (E) An adjacent serial section to (C,D), shows neutrophils in debris, epithelial and sub mucosa layer following NACE staining. (F) Virus antigen is still present in some bronchiolar epithelial cells. (G) H&E-stained section which shows hyperplastic bronchial epithelium with neutrophil infiltration, debris in lumen, necrosis (Br) and inflammatory cells infiltrates in lung parenchyma, a blood vessel with thickened endothelium (BV) and perivascular edema. (H) An adjacent serial section to (G) shows small pockets of viral antigen in pulmonary parenchyma (consolidation) but not in bronchial epithelium. (I) H&E-stained lung tissue shows perivascular (BV) and peri-bronchial inflammatory cell infiltration, alveolar exudates, and alveolar epithelial hyperplasia with mitosis. (J) A BV with proliferated endothelial cells Monocytes and eosinophils, and fibroblasts (undergoing mitosis) are distributed around blood vessels. Necrotic lung tissues show damaged alveolar walls and inflammatory cell infiltrates in consolidation area. Bar: (A,B) = 1 mm; (C–F) = 20 µm; (G,H,J) = 100 µm; (I) = 50 µm.
![Viruses 14 01403 g007]()
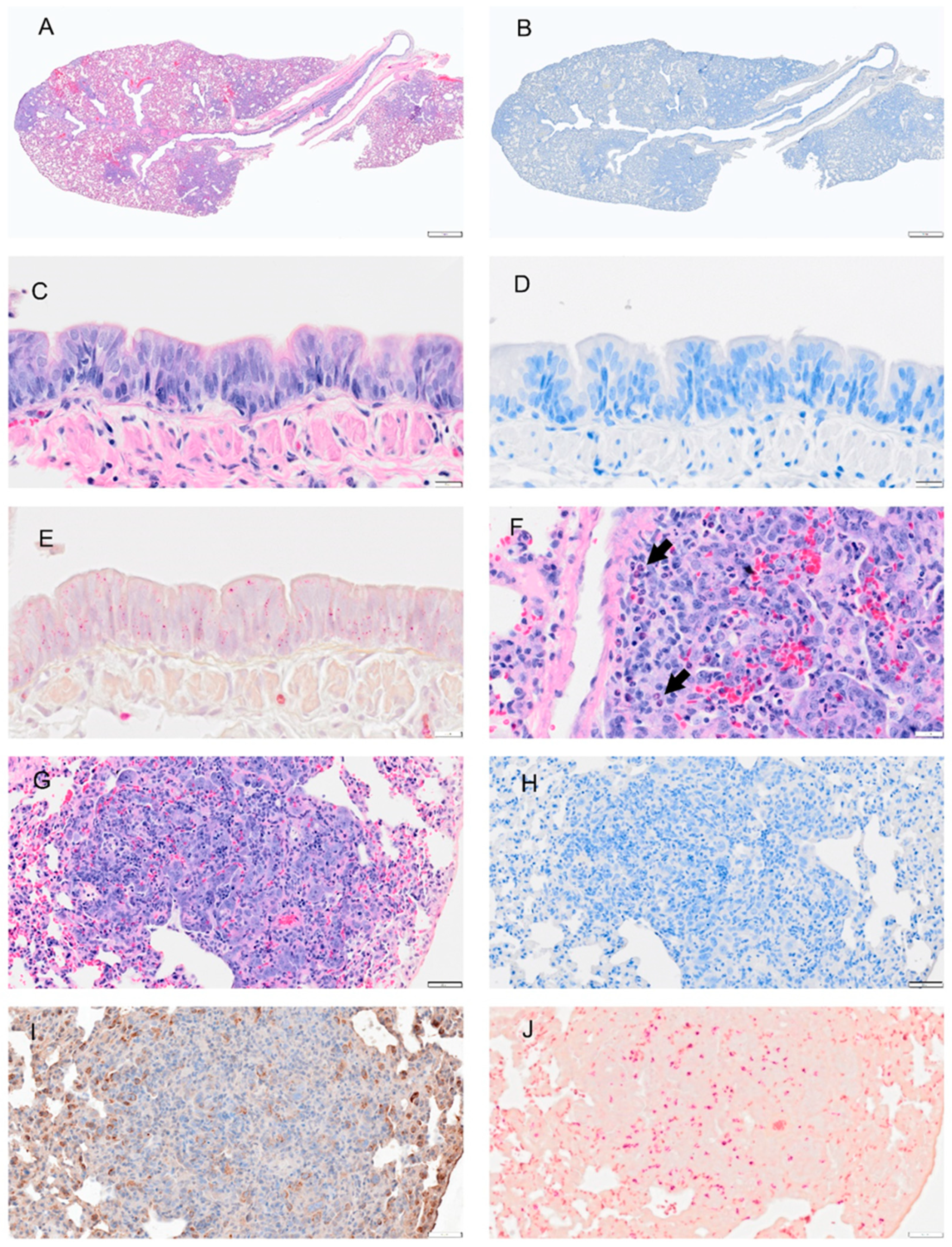
Figure 8.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 8 dpi with SARS-CoV-2. (A) H&E section shows well demarcated areas of consolidation and multifocal hemorrhage. (B) A representative section of the lung was subjected to IHC for the discovery of viral antigens. No staining for viral antigen was observed. (C) A higher magnification image of bronchial epithelial cells shows hyperplasia and few inflammatory cells (D). No viral staining was detected in adjacent serial section to (C). (E) A representative image stained with NACE stain shows no neutrophils in mucosa or submucosa. (F) H&E imaging shows increased eosinophils and neutrophils in the interstitial tissues (arrow) of the lung. Alveolar epithelial are hypertrophic and hyperplastic. (G) Representative section of the lung showing alveolar bronchiolization and inflammatory cell infiltration. (H) An adjacent section of (G) probed by IHC for viral antigen. No viral antigen was detected. (I) An adjacent serial section to (G/H) probed for prosurfactant protein C. IHC shows positive signals in cytoplasm of proliferating type 2 alveolar cells. (J) An adjacent serial section to (G/H) stained with NACE stain and shows neutrophils in these foci. Scalebar: (A,B) = 1 mm; (C–F) = 20 µm; (G–J) = 50 µm.
Figure 8.
Selected pathology and immunohistochemistry of lungs from Syrian hamsters at 8 dpi with SARS-CoV-2. (A) H&E section shows well demarcated areas of consolidation and multifocal hemorrhage. (B) A representative section of the lung was subjected to IHC for the discovery of viral antigens. No staining for viral antigen was observed. (C) A higher magnification image of bronchial epithelial cells shows hyperplasia and few inflammatory cells (D). No viral staining was detected in adjacent serial section to (C). (E) A representative image stained with NACE stain shows no neutrophils in mucosa or submucosa. (F) H&E imaging shows increased eosinophils and neutrophils in the interstitial tissues (arrow) of the lung. Alveolar epithelial are hypertrophic and hyperplastic. (G) Representative section of the lung showing alveolar bronchiolization and inflammatory cell infiltration. (H) An adjacent section of (G) probed by IHC for viral antigen. No viral antigen was detected. (I) An adjacent serial section to (G/H) probed for prosurfactant protein C. IHC shows positive signals in cytoplasm of proliferating type 2 alveolar cells. (J) An adjacent serial section to (G/H) stained with NACE stain and shows neutrophils in these foci. Scalebar: (A,B) = 1 mm; (C–F) = 20 µm; (G–J) = 50 µm.
![Viruses 14 01403 g008]()
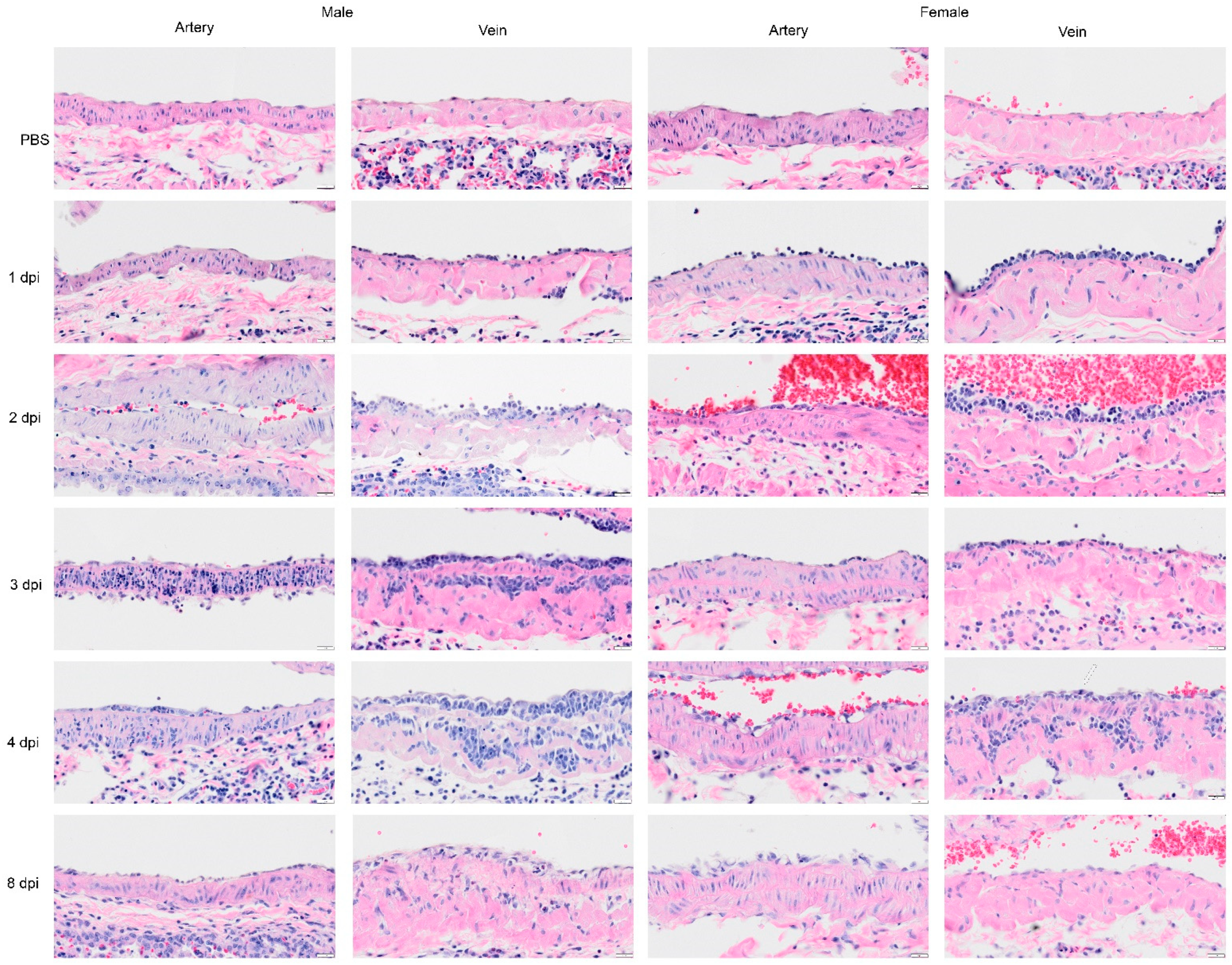
Figure 9.
Selected H&E images of the large pulmonary blood vessels from Syrian hamster following infection with SARS-CoV-2 on 1, 2, 3, 4, and 8 dpi. In the sham inoculated group (PBS), the main pulmonary artery and veins display a normal character in each layer of the blood vessel. In subsequent rows, representative images of pulmonary artery and veins are presented for 1, 2, 3, 4, and 8 dpi. The pulmonary artery at 1 dpi is normal, although in one female hamster, a few leucocytes were attached to the tunica intima. At 1 dpi, the image of the pulmonary vein shows leucocytes attached to the wall of pulmonary veins. At 2 dpi, blood vessels show an increased level of inflammatory cells or leucocytes penetrating through endothelial cells, and tunica intima, and appeared to be greater in the vein. The image on 3 dpi shows neutrophils that have penetrated through tunica media of pulmonary artery (male) or less severe (female). In the image of the pulmonary veins, the inflammatory cells have clotted in the tunica media. The image on 4 dpi shows inflammatory cells passing through an intact endothelial cell layer of the artery (male) although some vacuoles are present in endothelial cells and in muscle cells (female). The image of the vein shows the infiltration and proliferation of mononuclear cells. Mitosis was also noted in the endothelial cells (arrow in female vein implies proliferation of endothelial cells). We also noted groups of increased cells with single nuclei in the tunica media (smooth muscle cells). By 8 dpi, there was a decrease in the level of inflammatory cells infiltrating the blood vessels (recovered), and a higher density of endothelial cells in tunica intima. Scalebar = 20 μm.
Figure 9.
Selected H&E images of the large pulmonary blood vessels from Syrian hamster following infection with SARS-CoV-2 on 1, 2, 3, 4, and 8 dpi. In the sham inoculated group (PBS), the main pulmonary artery and veins display a normal character in each layer of the blood vessel. In subsequent rows, representative images of pulmonary artery and veins are presented for 1, 2, 3, 4, and 8 dpi. The pulmonary artery at 1 dpi is normal, although in one female hamster, a few leucocytes were attached to the tunica intima. At 1 dpi, the image of the pulmonary vein shows leucocytes attached to the wall of pulmonary veins. At 2 dpi, blood vessels show an increased level of inflammatory cells or leucocytes penetrating through endothelial cells, and tunica intima, and appeared to be greater in the vein. The image on 3 dpi shows neutrophils that have penetrated through tunica media of pulmonary artery (male) or less severe (female). In the image of the pulmonary veins, the inflammatory cells have clotted in the tunica media. The image on 4 dpi shows inflammatory cells passing through an intact endothelial cell layer of the artery (male) although some vacuoles are present in endothelial cells and in muscle cells (female). The image of the vein shows the infiltration and proliferation of mononuclear cells. Mitosis was also noted in the endothelial cells (arrow in female vein implies proliferation of endothelial cells). We also noted groups of increased cells with single nuclei in the tunica media (smooth muscle cells). By 8 dpi, there was a decrease in the level of inflammatory cells infiltrating the blood vessels (recovered), and a higher density of endothelial cells in tunica intima. Scalebar = 20 μm.
![Viruses 14 01403 g009]()
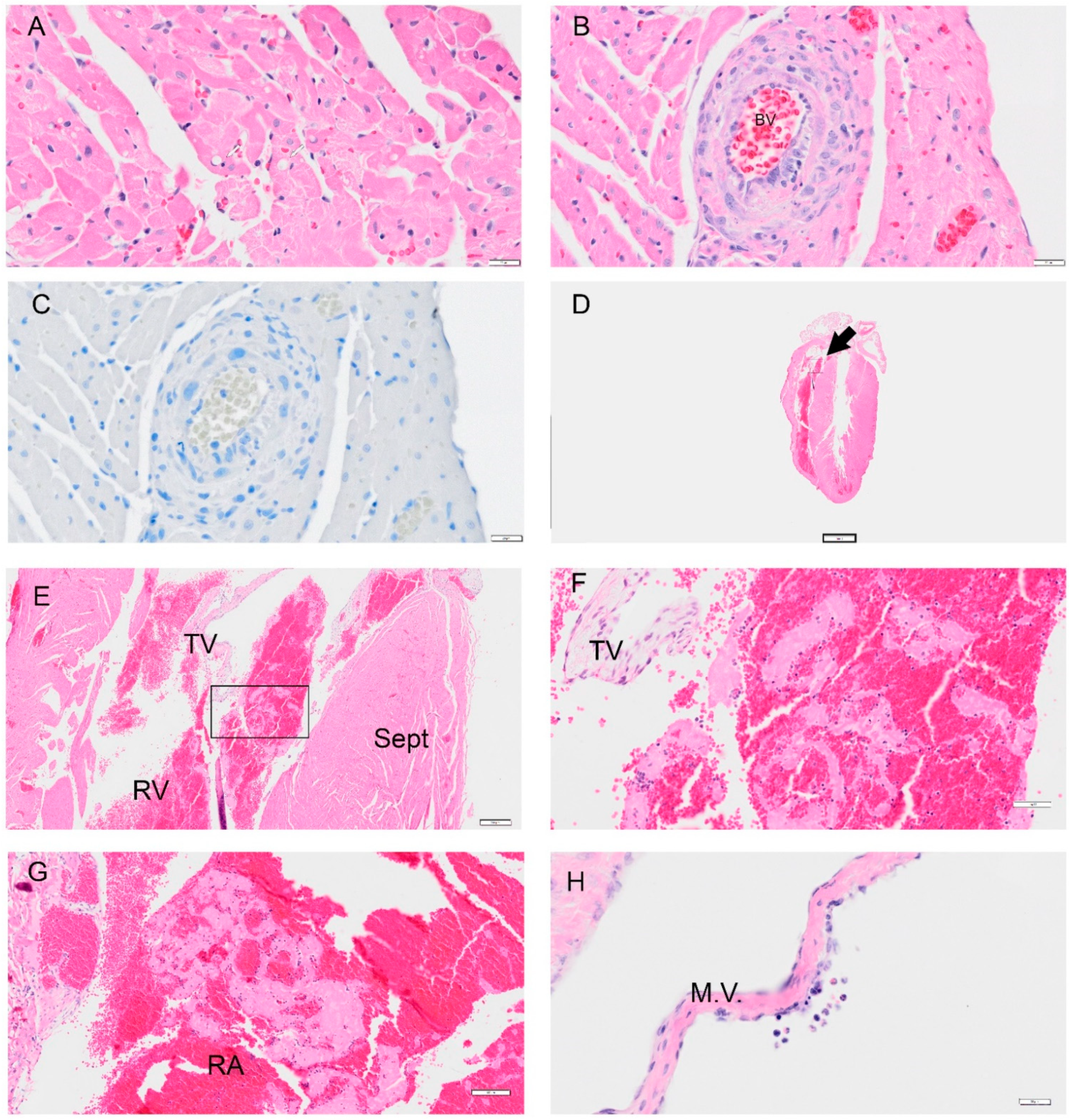
Figure 10.
Selected pathology and immunohistochemistry of the heart from the Syrian hamster following infection with SARS-CoV-2. Representative images of pathology are presented. (A) An image at 1 dpi from the right ventricle showed mild degeneration of cardiomyocytes. (B) An image from 1 dpi shows an abnormal muscle layer in the coronary artery. (C) A representative image of the heart probed for viral antigen by IHC Viral N protein was not detected in over 80 slides. (D) A low magnification image of the heart at 8 dpi in a female hamster with an arrow showing a thrombus (0.4×). (E) An image of a right ventricle with a platelet and fibrin aggregate (4×). (F) Higher magnification of (E) which in the right ventricle with white blood cells (WBC) trapped in fibrin and red blood cells (RBC); close to the tricuspid valve (TV). (G) An image showing a thrombus in the right atrium (RA) with similar findings in the right ventricle in (F). (H) Image showing endothelial cell abnormality with hyperplasia of the endothelial cells. Image shows WBC (mononuclear cells and eosinophils) around a damaged area on the mitral valve (M.V.) (female 8 dpi). Abbreviations: RV: right ventricle; Sept: septum; TV: tricuspid valve. Scalebar: (A–C,H) = 20 μm; (D): = 2 mm, (E) = 200 μm; (F) = 50 μm, (G) = 100 μm.
Figure 10.
Selected pathology and immunohistochemistry of the heart from the Syrian hamster following infection with SARS-CoV-2. Representative images of pathology are presented. (A) An image at 1 dpi from the right ventricle showed mild degeneration of cardiomyocytes. (B) An image from 1 dpi shows an abnormal muscle layer in the coronary artery. (C) A representative image of the heart probed for viral antigen by IHC Viral N protein was not detected in over 80 slides. (D) A low magnification image of the heart at 8 dpi in a female hamster with an arrow showing a thrombus (0.4×). (E) An image of a right ventricle with a platelet and fibrin aggregate (4×). (F) Higher magnification of (E) which in the right ventricle with white blood cells (WBC) trapped in fibrin and red blood cells (RBC); close to the tricuspid valve (TV). (G) An image showing a thrombus in the right atrium (RA) with similar findings in the right ventricle in (F). (H) Image showing endothelial cell abnormality with hyperplasia of the endothelial cells. Image shows WBC (mononuclear cells and eosinophils) around a damaged area on the mitral valve (M.V.) (female 8 dpi). Abbreviations: RV: right ventricle; Sept: septum; TV: tricuspid valve. Scalebar: (A–C,H) = 20 μm; (D): = 2 mm, (E) = 200 μm; (F) = 50 μm, (G) = 100 μm.
![Viruses 14 01403 g010]()


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









