
Comparative Evaluation of Immune Responses and Protection of Chitosan Nanoparticles and Oil-Emulsion Adjuvants in Avian Coronavirus Inactivated Vaccines in Chickens

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus
2.2. Preparations of Vaccines
2.2.1. AvCoV-CS
2.2.2. AvCoV-O
2.2.3. H120 Attenuated Vaccine (L)
2.3. In Vivo Vaccine Evaluation
2.3.1. Microscopic Alterations
2.3.2. RNA Extraction
2.3.3. Quantification of AvCoV Load
2.3.4. Quantification of the Expression of Cell-Mediated Immune (CMI) Response Genes
2.3.5. Evaluation of Mucosal and Systemic Anti-AvCoV Antibody Responses
2.4. Statistical Analysis
3. Results
3.1. Microscopic Alterations
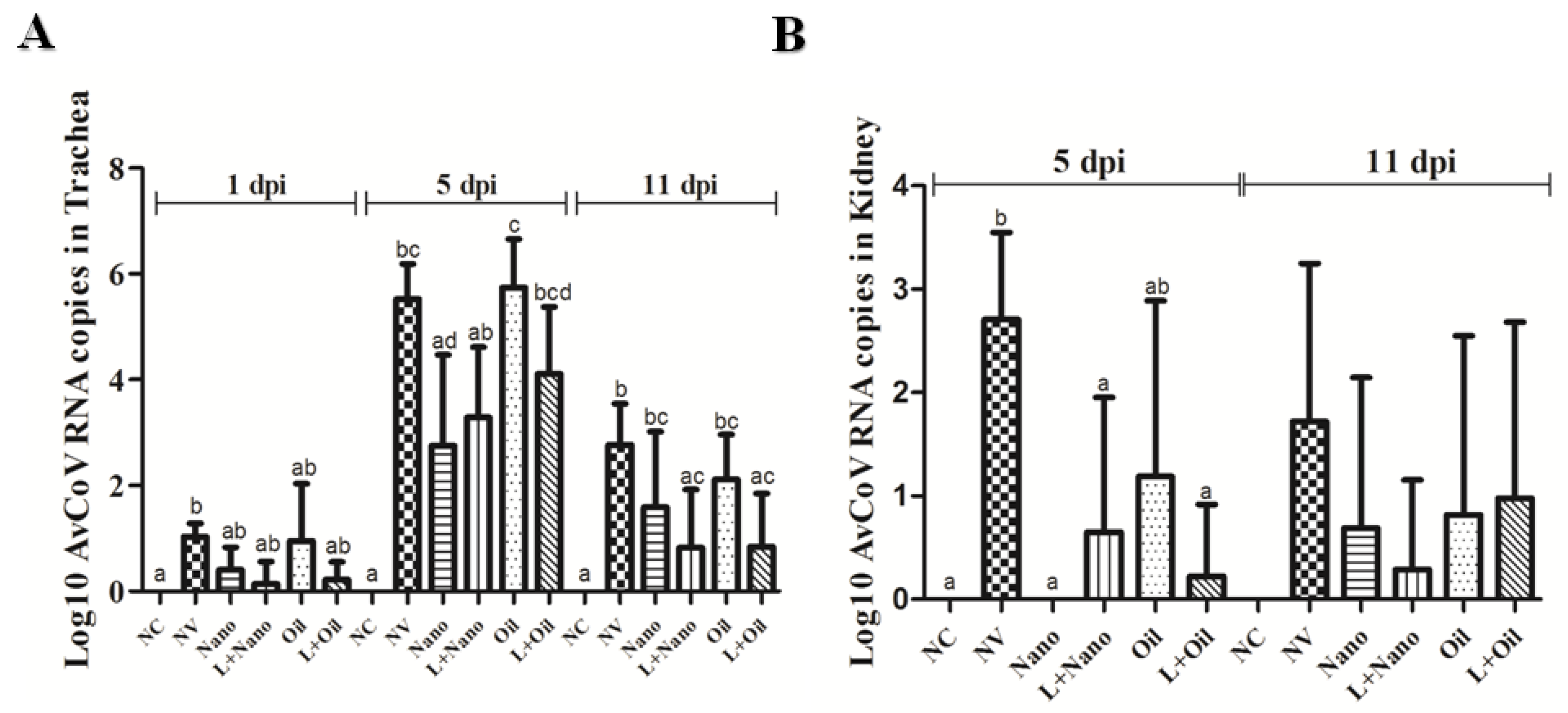
3.2. Quantification of AvCoV Load in Tissue Samples
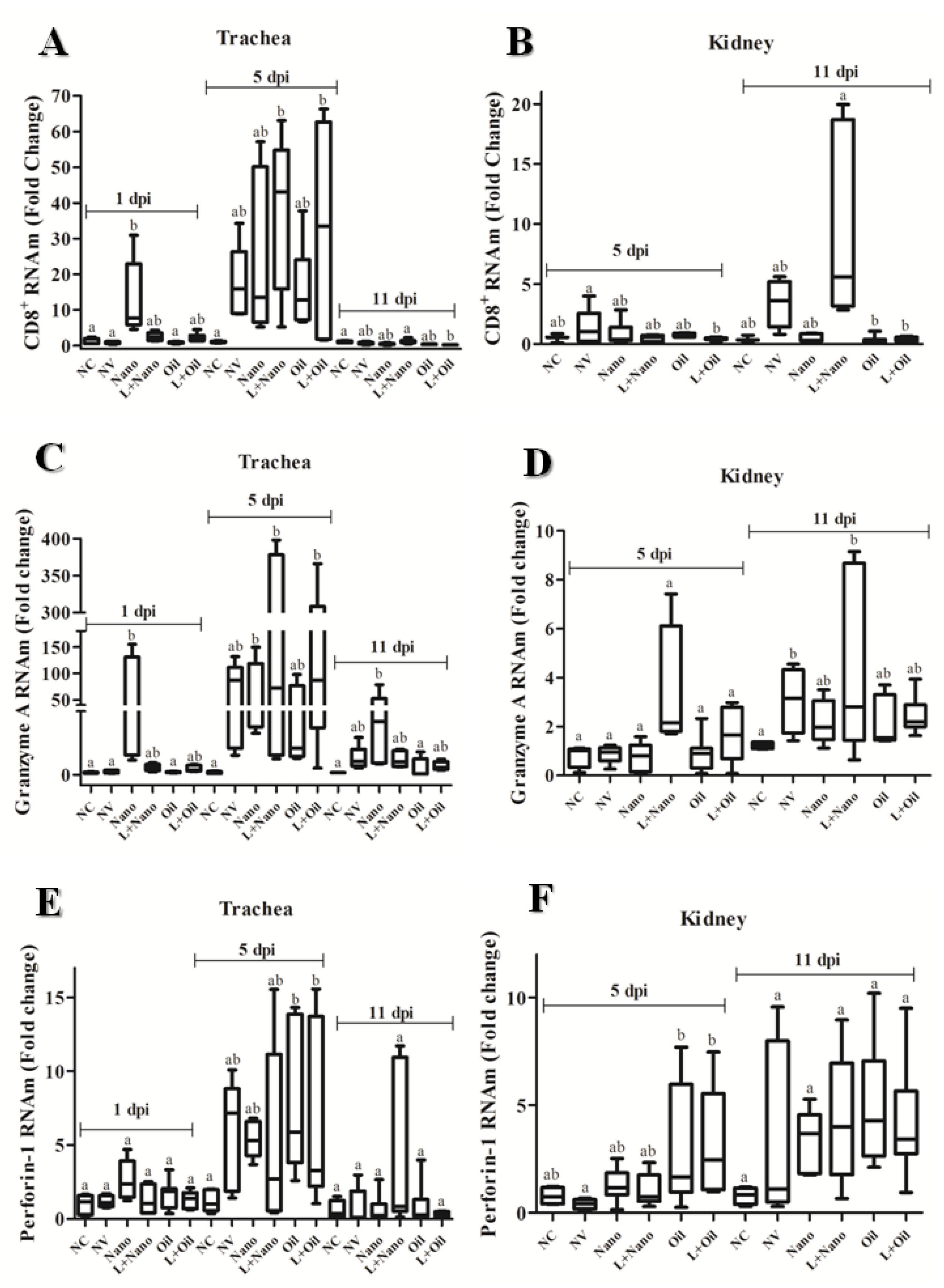
3.3. CMI Responses
3.4. Evaluation of Mucosal and Systemic Anti-AvCoV Antibody Responses
3.5. Correlation between Parameters of the Immune Response against AvCoV Induced by Vaccines and the Pathological Changes Caused by This Virus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chhabra, R.; Chantrey, J.; Ganapathy, K. Immune Responses to Virulent and Vaccine Strains of Infectious Bronchitis Viruses in Chickens. Viral Immunol. 2015, 28, 478–488. [Google Scholar] [CrossRef]
- Chhabra, R.; Forrester, A.; Lemiere, S.; Awad, F.; Chantrey, J.; Ganapathy, K. Mucosal, cellular, and humoral immune responses induced by different live infectious bronchitis virus vaccination regimes and protection conferred against infectious bronchitis virus Q1 strain. Clin. Vaccine Immunol. 2015, 22, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Okino, C.H.; Alessi, A.C.; Montassier, M.D.F.S.; Rosa, A.J.D.M.; Wang, X.; Montassier, H.J. Humoral and cell-mediated immune responses to different doses of attenuated vaccine against avian infectious bronchitis virus. Viral Immunol. 2013, 26, 259–267. [Google Scholar] [CrossRef]
- Al-Rasheed, M.; Ball, C.; Ganapathy, K. Route of infectious bronchitis virus vaccination determines the type and magnitude of immune responses in table egg laying hens. Vet. Res. 2021, 52, 139. [Google Scholar] [CrossRef]
- Jordan, B. Vaccination against infectious bronchitis virus: A continuous challenge. Vet. Microbiol. 2017, 206, 137–143. [Google Scholar] [CrossRef]
- Guzmán, M.; Hidalgo, H. Live Attenuated Infectious Bronchitis Virus Vaccines in Poultry: Modifying Local Viral Populations Dynamics. Animals 2020, 10, 2058. [Google Scholar] [CrossRef]
- Ennaji, Y.; Khataby, K.; Ennaji, M.M. Infectious Bronchitis Virus in Poultry: Molecular Epidemiology and Factors Leading to the Emergence and Reemergence of Novel Strains of Infectious Bronchitis Virus. In Emerging and Reemerging Viral Pathogens; Elsevier: Amsterdam, The Netherlands, 2020; pp. 31–44. [Google Scholar]
- Legnardi, M.; Tucciarone, C.M.; Franzo, G.; Cecchinato, M. Infectious Bronchitis Virus Evolution, Diagnosis and Control. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, A.; Gopalakrishnamurthy, T.R.; Sivaseelan, S.; Balasubramaniam, G.A.; Johnson Rajeswar, J. Evaluation of an inactivated vaccine for nephropathogenic infectious bronchitis virus. Vet. World 2013, 6, 134–138. [Google Scholar] [CrossRef]
- Jackwood, M.W.; De Wit, S. Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; Wiley: Hoboken, NJ, USA, 2020; ISBN 9781119371168. [Google Scholar]
- Aucouturier, J.; Dupuis, L.; Ganne, V. Adjuvants designed for veterinary and human vaccines. Vaccine 2001, 19, 2666–2672. [Google Scholar] [CrossRef]
- Chandrasekar, S.S.; Kingstad-Bakke, B.; Wu, C.-W.; Suresh, M.; Talaat, A.M. A Novel Mucosal Adjuvant System for Immunization against Avian Coronavirus Causing Infectious Bronchitis. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Sawaengsak, C.; Mori, Y.; Yamanishi, K.; Mitrevej, A.; Sinchaipanid, N. Chitosan nanoparticle encapsulated hemagglutinin-split influenza virus mucosal vaccine. AAPS PharmSciTech 2014, 15, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yin, Z.; Liu, N.; Yang, T.; Wang, J.; Bu, Z.; Wu, D. DNA–chitosan nanoparticles improve DNA vaccine-elicited immunity against Newcastle disease virus through shuttling chicken interleukin-2 gene. J. Microencapsul. 2010, 27, 693–702. [Google Scholar] [CrossRef]
- Zhao, K.; Chen, G.; Shi, X.M.; Gao, T.T.; Li, W.; Zhao, Y.; Zhang, F.Q.; Wu, J.; Cui, X.; Wang, Y.F. Preparation and Efficacy of a Live Newcastle Disease Virus Vaccine Encapsulated in Chitosan Nanoparticles. PLoS ONE 2012, 7, e53314. [Google Scholar] [CrossRef] [Green Version]
- Renu, S.; Markazi, A.D.; Dhakal, S.; Lakshmanappa, Y.S.; Shanmugasundaram, R.; Selvaraj, R.K.; Renukaradhya, G.J. Oral deliverable mucoadhesive Chitosan-Salmonella subunit nanovaccine for layer chickens. Int. J. Nanomed. 2020, 15, 761–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaikabo, A.A.; AbdulKarim, S.M.; Abas, F. Evaluation of the efficacy of chitosan nanoparticles loaded ΦKAZ14 bacteriophage in the biological control of colibacillosis in chickens. Poult. Sci. 2017, 96, 295–302. [Google Scholar] [CrossRef]
- Zhao, K.; Rong, G.; Hao, Y.; Yu, L.; Kang, H.; Wang, X.; Wang, X.; Jin, Z.; Ren, Z.; Li, Z. IgA response and protection following nasal vaccination of chickens with Newcastle disease virus DNA vaccine nanoencapsulated with Ag@SiO2 hollow nanoparticles. Sci. Rep. 2016, 6, 25720. [Google Scholar] [CrossRef] [Green Version]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Omar, A.R.; Moeini, H.; Khadkodaei, S.; Wei, T.S.; Keong, Y.S.; Abba, Y.; Anka, I.A. Development and immunogenic potentials of chitosan-saponin encapsulated DNA vaccine against avian infectious bronchitis coronavirus. Microb. Pathog. 2020, 149, 104560. [Google Scholar] [CrossRef] [PubMed]
- Lopes, P.D.; Okino, C.H.; Fernando, F.S.; Pavani, C.; Casagrande, V.M.; Lopez, R.F.V.; Montassier, M.D.F.S.; Montassier, H.J. Inactivated infectious bronchitis virus vaccine encapsulated in chitosan nanoparticles induces mucosal immune responses and effective protection against challenge. Vaccine 2018, 36, 2630–2636. [Google Scholar] [CrossRef] [PubMed]
- Klimka, A.; Michels, L.; Glowalla, E.; Tosetti, B.; Kr??nke, M.; Krut, O. Montanide ISA 71 VG is Advantageous to Freund’s Adjuvant in Immunization Against, S. aureus Infection of Mice. Scand. J. Immunol. 2015, 81, 291–297. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Lillehoj, E.P.; Bertrand, F.; Dupuis, L.; Deville, S. Montanide™ ISA 71 VG adjuvant enhances antibody and cell-mediated immune responses to profilin subunit antigen vaccination and promotes protection against Eimeria acervulina and Eimeria tenella. Exp. Parasitol. 2011, 127, 178–183. [Google Scholar] [CrossRef]
- Lone, N.A.; Spackman, E.; Kapczynski, D. Immunologic evaluation of 10 different adjuvants for use in vaccines for chickens against highly pathogenic avian influenza virus. Vaccine 2017, 35, 3401–3408. [Google Scholar] [CrossRef]
- Santos, R.M.; Fernando, F.S.; Montassier, M.D.F.S.; Silva, K.R.; Lopes, P.D.; Pavani, C.; Borzi, M.M.; Okino, C.H.; Montassier, H.J. Memory immune responses and protection of chickens against a nephropathogenic infectious bronchitis virus strain by combining live heterologous and inactivated homologous vaccines. J. Vet. Med. Sci. 2019, 81, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.I.; Kim, D.K.; Lillehoj, H.S.; Lee, S.H.; Lee, K.W.; Bertrand, F.; Dupuis, L.; Deville, S.; Ben Arous, J.; Lillehoj, E.P. Evaluation of MontanideTM ISA 71 VG Adjuvant during Profilin Vaccination against Experimental Coccidiosis. PLoS ONE 2013, 8, e59786. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Zhang, Y.; Zhang, X.; Li, W.; Shi, C.; Guo, C.; Dai, C.; Chen, Q.; Jin, Z.; Zhao, Y.; et al. Preparation and efficacy of newcastle disease virus dna vaccine encapsulated in chitosan nanoparticles. Int. J. Nanomedicine 2014, 9, 389–402. [Google Scholar] [CrossRef] [Green Version]
- Owen, R.L.; Cowen, B.S.; Hattel, A.L.; Naqi, S.A.; Wilson, R.A. Detection of viral antigen following exposure of one-day-old chickens to the Holland 52 strain of infectious bronchitis virus. Avian Pathol. 1991, 20, 663–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, J.S. Avian infectious bronchitis: The protection afforded by an inactivated virus vaccine. Vet. Rec. 1969, 85, 378–381. [Google Scholar] [CrossRef]
- Chen, B.Y.; Hosi, S.; Nunoya, T.; Itakura, C. Histopathology and immunohistochemistry of renal lesions due to infectious bronchitis virus in chicks. Avian Pathol. 1996, 25, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Fernando, F.S.; Okino, C.H.; Silva, K.R.; Fernandes, C.C.; Gonçalves, M.C.M.; Montassier, M.F.S.; Vasconcelos, R.O.; Montassier, H.J. Increased expression of Interleukin-6 related to nephritis in chickens challenged with an Avian infectious bronchitis virus variant. Pesqui. Vet. Bras. 2015, 35, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Cook, J.K.; Otsuki, K.; Huggins, M.B.; Frazier, J.A. Comparative study of respiratory lesions in two chicken lines of different susceptibility infected with infectious bronchitis virus: Histology, ultrastructure and immunohistochemistry. Avian Pathol. 1991, 20, 241–257. [Google Scholar] [CrossRef]
- Okino, C.H.; Mores, M.A.Z.; Trevisol, I.M.; Coldebella, A.; Montassier, H.J.; Brentano, L. Early immune responses and development of pathogenesis of avian infectious bronchitis viruses with different virulence profiles. PLoS ONE 2017, 12, e0172275. [Google Scholar] [CrossRef] [PubMed]
- Chousalkar, K.K.; Cheetham, B.F.; Roberts, J.R. LNA probe-based real-time RT-PCR for the detection of infectious bronchitis virus from the oviduct of unvaccinated and vaccinated laying hens. J. Virol. Methods 2009, 155, 67–71. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Bronzoni, R.V.M.; Fatima, M.; Montassier, S.; Pereira, G.T.; Gama, N.M.S.Q.; Sakai, V.; Montassier, H.J. Detection of infectious bronchitis virus and specific anti- viral antibodies using a Concanavalin A-Sandwich-ELISA. Viral Immunol. 2005, 18, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhuiyan, M.S.A.; Amin, Z.; Bakar, A.M.S.A.; Saallah, S.; Yusuf, N.H.M.; Shaarani, S.M.; Siddiquee, S. Factor Influences for Diagnosis and Vaccination of Avian Infectious Bronchitis Virus (Gammacoronavirus) in Chickens. Vet. Sci. 2021, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Rosa, A.J.M.; Chen, D.-G.; Wang, X. Molecular mechanisms of primary and secondary mucosal immunity using avian infectious bronchitis virus as a model system. Vet. Immunol. Immunopathol. 2008, 121, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Senevirathne, A.; Hewawaduge, C.; Kim, J.; Lee, J.H. Intranasally administered protein coated chitosan nanoparticles encapsulating influenza H9N2 HA2 and M2e mRNA molecules elicit protective immunity against avian influenza viruses in chickens. Vet. Res. 2020, 51, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renu, S.; Markazi, A.D.; Dhakal, S.; Lakshmanappa, Y.S.; Gourapura, S.R.; Shanmugasundaram, R.; Senapati, S.; Narasimhan, B.; Selvaraj, R.K.; Renukaradhya, G.J. Surface engineered polyanhydride-based oral Salmonella subunit nanovaccine for poultry. Int. J. Nanomedicine 2018, 13, 8195. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.Z.; Wen, Y.C.; Lin, S.Y.; Chen, T.C.; Chen, H.W. Anti-influenza protective efficacy of a h6 virus-like particle in chickens. Vaccines 2020, 13, 465. [Google Scholar] [CrossRef]
- Pipkin, M.E.; Lieberman, J. Delivering the kiss of death: Progress on understanding how perforin works. Curr. Opin. Immunol. 2007, 19, 301–308. [Google Scholar] [CrossRef]
- Gurjar, R.S.; Gulley, S.L.; van Ginkel, F.W. Cell-mediated immune responses in the head-associated lymphoid tissues induced to a live attenuated avian coronavirus vaccine. Dev. Comp. Immunol. 2013, 41, 715–722. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | 1st Day of Age | 14th Day of Age | 31st Day Of Age |
|---|---|---|---|
| NC | - | Nanoparticles with no virus | DMEM culture medium |
| NV | - | - | Challenged with AvCoV/Brazil/PR05 strain |
| Nano | - | Inactivated AvCoV-CS vaccine | Challenged with AvCoV/Brazil/PR05 strain |
| L+Nano | Attenuated H120 strain vaccine | Inactivated AvCoV-CS vaccine | Challenged with AvCoV/Brazil/PR05 strain |
| Oil | - | Inactivated AvCoV-O vaccine | Challenged with AvCoV/Brazil/PR05 strain |
| L+Oil | Attenuated H120 strain vaccine | Inactivated AvCoV-O vaccine | Challenged with AvCoV/Brazil/PR05 strain |
| Group | Trachea | Kidney |
|---|---|---|
| NC | 0.0 ± 0.0 a | 0.0 ± 0.0 a |
| NV | 1.241 ± 1.331 bd | 0.9778 ± 0.9412 b |
| Nano | 0.8000 ± 0.9467 bc | 0.2222 ± 0.5520 ac |
| L+Nano | 0.5526 ± 0.8390 c | 0.5167 ± 0.7700 c |
| Oil | 1.575 ± 1.156 d | 0.8889 ± 0.9000 b |
| L+Oil | 0.9375 ± 1.194 bc | 0.5556 ± 0.9801 bc |
| IgA | IgG | CD8β | Granzyme | Perforin | |
|---|---|---|---|---|---|
| Viral load 5 dpi (T) | −0.499 * | −0.366 | −0.323 | −0.553 * | −0.147 |
| Viral load 11 dpi (T) | −0.413 * | −0.487 * | −0.245 | −0.551 * | - |
| Viral load 5 dpi (K) | - | −0.533 * | - | - | −0.508 |
| Viral load 11 dpi (K) | - | −0.181 | - | −0.517 * | - |
| IgA | IgG | CD8β | Granzyme | Perforin | |
|---|---|---|---|---|---|
| Viral load 5 dpi (T) | - | −0.148 | −0.058 | - | - |
| Viral load 11 dpi (T) | - | −0.419 * | - | - | - |
| Viral load 5 dpi (K) | - | −0.438 * | - | −0.122 | −0.339 |
| Viral load 11 dpi (K) | - | −0.018 | −0.310 | −0.024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, P.D.; Okino, C.H.; Fernando, F.S.; Pavani, C.; Mariguela, V.C.; Montassier, M.d.F.S.; Montassier, H.J. Comparative Evaluation of Immune Responses and Protection of Chitosan Nanoparticles and Oil-Emulsion Adjuvants in Avian Coronavirus Inactivated Vaccines in Chickens. Vaccines 2021, 9, 1457. https://doi.org/10.3390/vaccines9121457
Lopes PD, Okino CH, Fernando FS, Pavani C, Mariguela VC, Montassier MdFS, Montassier HJ. Comparative Evaluation of Immune Responses and Protection of Chitosan Nanoparticles and Oil-Emulsion Adjuvants in Avian Coronavirus Inactivated Vaccines in Chickens. Vaccines. 2021; 9(12):1457. https://doi.org/10.3390/vaccines9121457
Chicago/Turabian StyleLopes, Priscila Diniz, Cintia Hiromi Okino, Filipe Santos Fernando, Caren Pavani, Viviane Casagrande Mariguela, Maria de Fátima Silva Montassier, and Hélio José Montassier. 2021. "Comparative Evaluation of Immune Responses and Protection of Chitosan Nanoparticles and Oil-Emulsion Adjuvants in Avian Coronavirus Inactivated Vaccines in Chickens" Vaccines 9, no. 12: 1457. https://doi.org/10.3390/vaccines9121457