
Long-Term Effectiveness of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine on B Cell Compartment: Efficient Recall of SARS-CoV-2-Specific Memory B Cells
 , ,
, ,  , , , ,
, , , ,  , , and
, , and 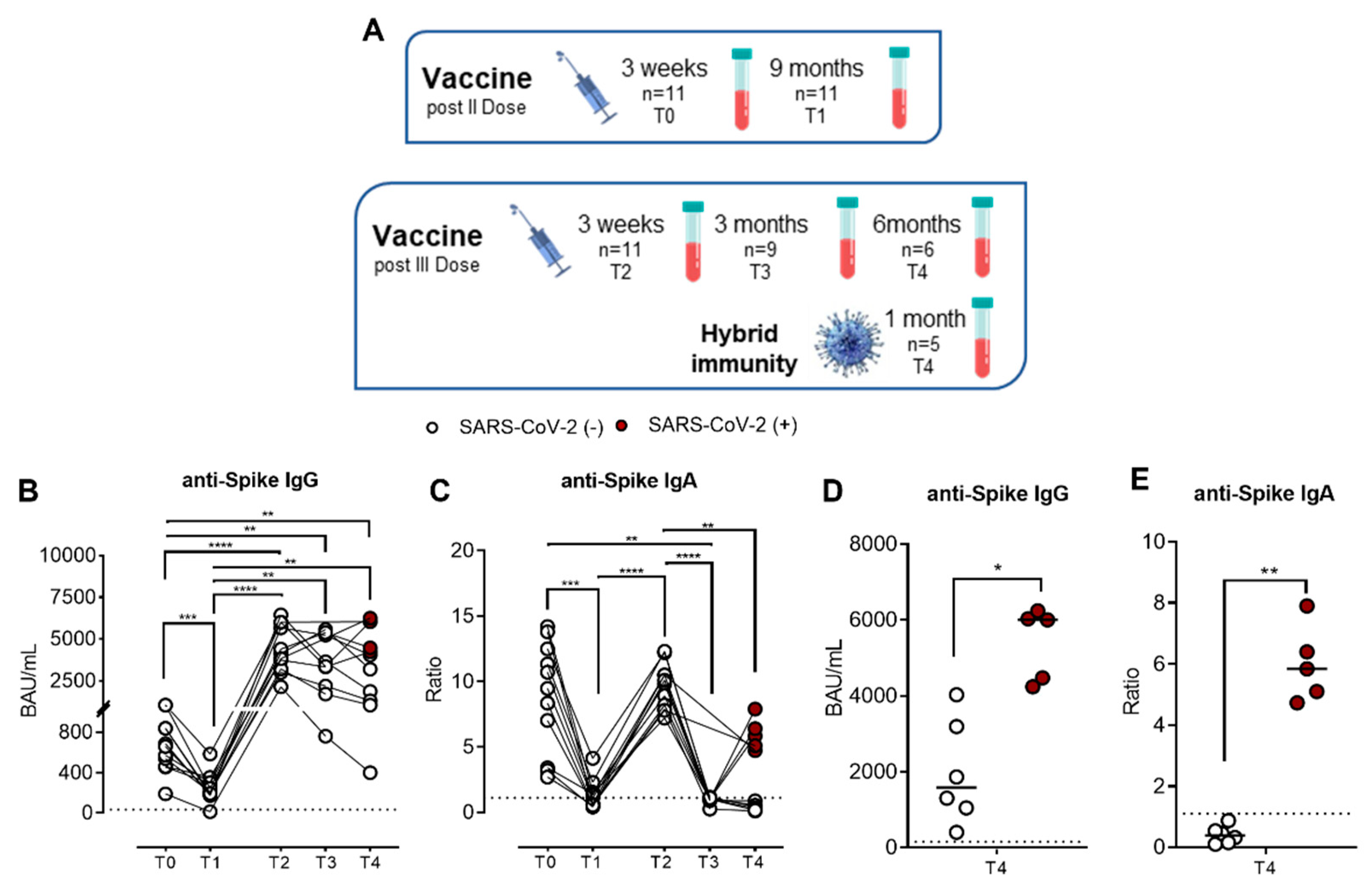{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. SARS-CoV-2-Specific IgG and IgA Kinetics after Pfizer-BioNTech BNT162b2 mRNA Vaccine
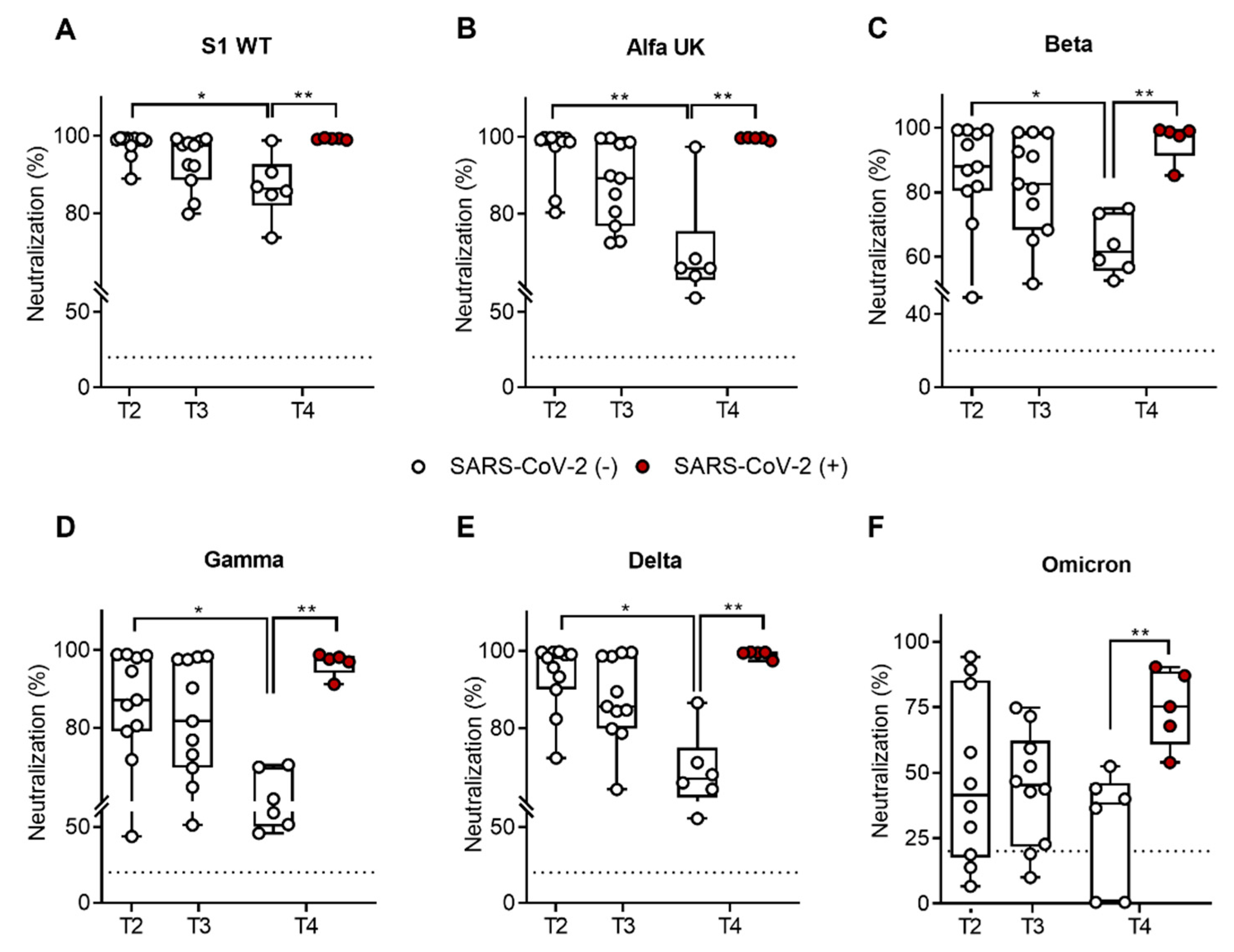
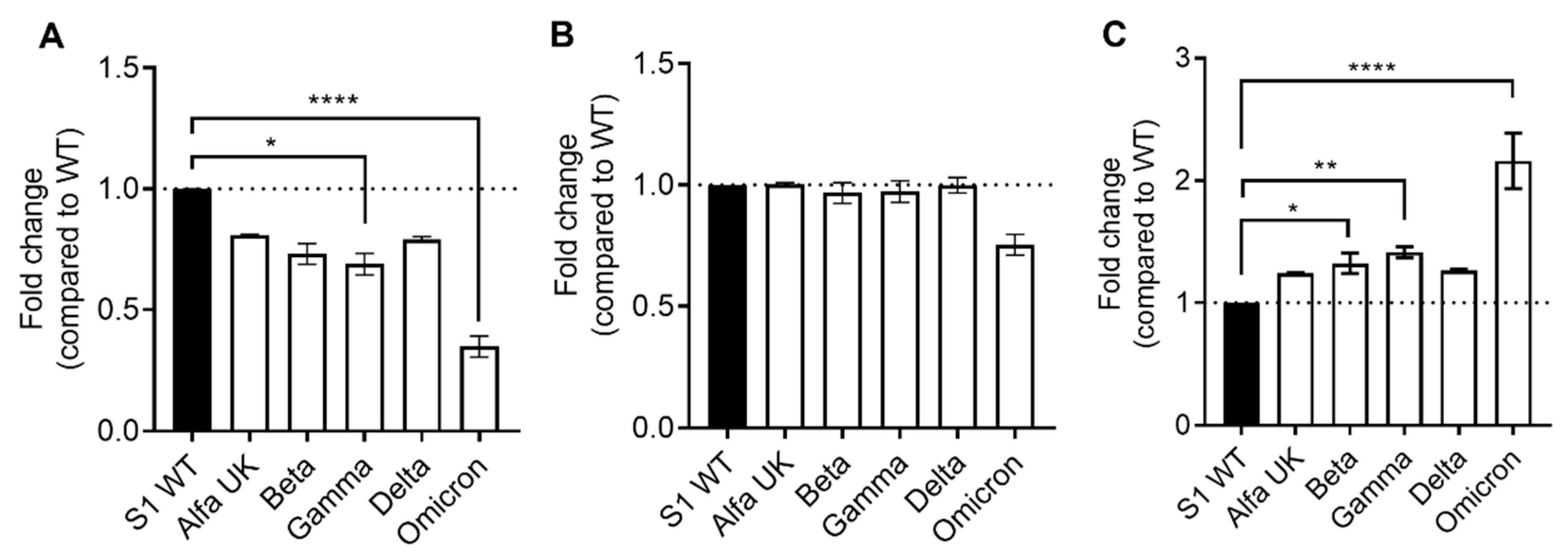
2.2. SARS-CoV-2-Specific IgG Neutralization Ability against SARS-CoV-2 Variants
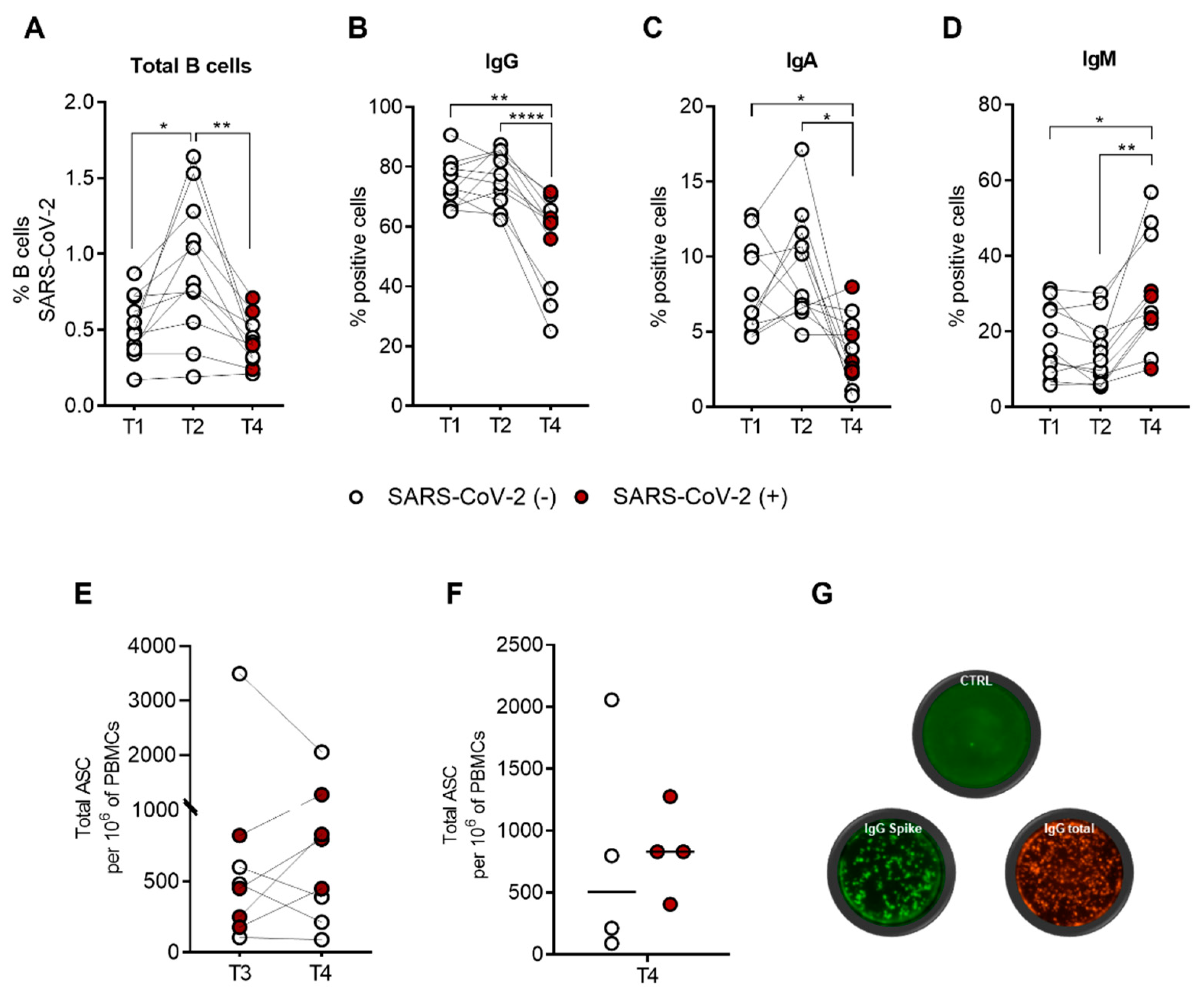
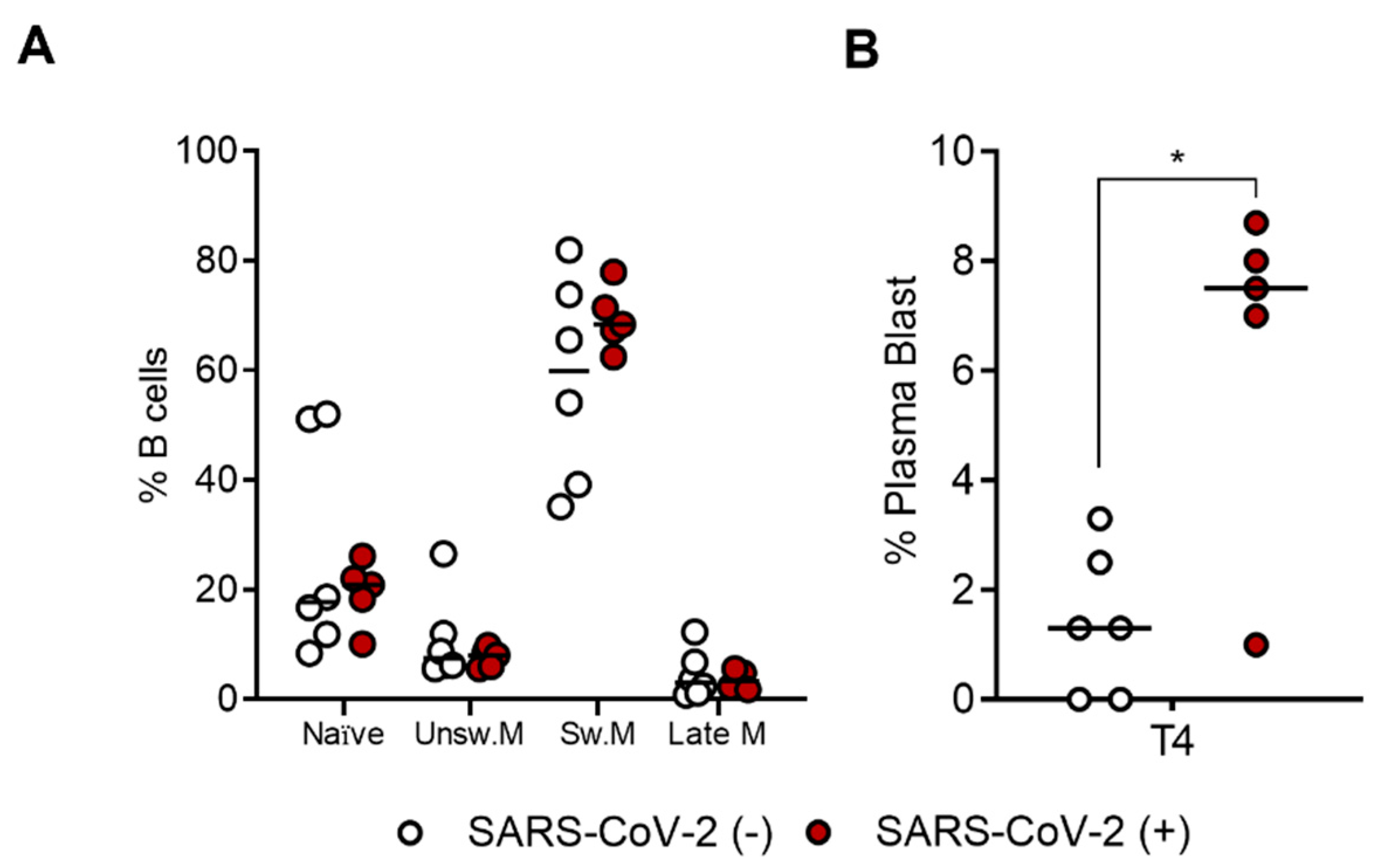
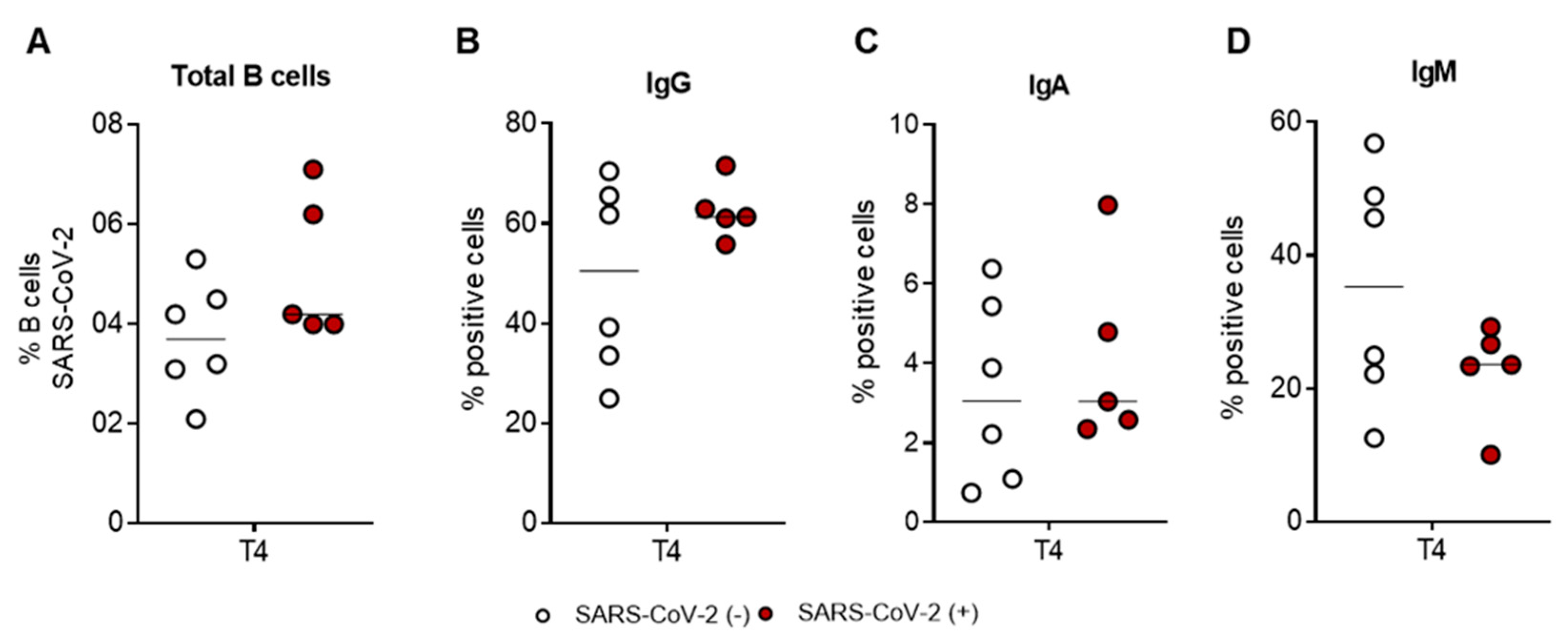
2.3. The SARS-CoV-2-Specific B-Cell Pool Is Stable until Six Months after the Third Dose of the Pfizer-BioNTech BNT162b2 mRNA Vaccine, Preserving Its Effector Function
2.4. SARS-CoV-2-Specific B-Cell Subpopulations Are Enriched in Plasma Blasts in Recovered Subjects
3. Discussion
4. Materials and Methods
4.1. Subjects Enrolled in the Study
4.2. Detection of SARS-CoV-2 Antibodies
4.3. Neutralization Assay
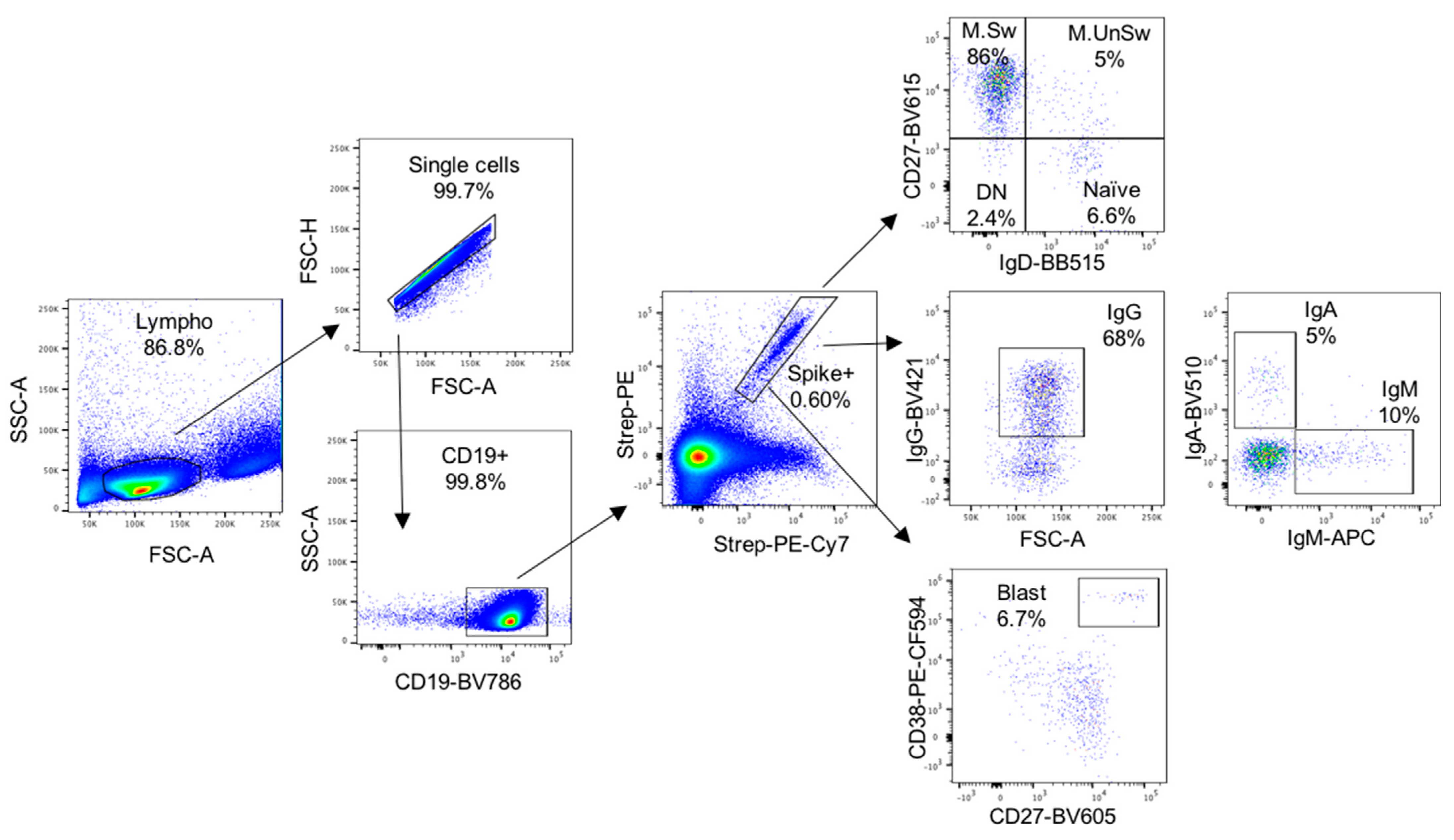
4.4. Isolation and Quantification of SARS-CoV-2-Specific B Cells
4.5. IgG SARS-CoV-2 FluoroSpot Assay
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Amano, M.; Otsu, S.; Maeda, K.; Uemura, Y.; Shimizu, Y.; Omata, K.; Matsuoka, M.; Shimada, S.; Mitsuya, H. Neutralization activity of sera/IgG preparations from fully BNT162b2 vaccinated individuals against SARS-CoV-2 Alpha, Beta, Gamma, Delta, and Kappa variants. Sci. Rep. 2022, 12, 13524. [Google Scholar] [CrossRef] [PubMed]
- Dulovic, A.; Kessel, B.; Harries, M.; Becker, M.; Ortmann, J.; Griesbaum, J.; Jungling, J.; Junker, D.; Hernandez, P.; Gornyk, D.; et al. Comparative Magnitude and Persistence of Humoral SARS-CoV-2 Vaccination Responses in the Adult Population in Germany. Front. Immunol. 2022, 13, 828053. [Google Scholar] [CrossRef] [PubMed]
- Fodor, E.; Olmos Calvo, I.; Kuten-Pella, O.; Hamar, E.; Bukva, M.; Madar, A.; Hornyak, I.; Hinsenkamp, A.; Hetenyi, R.; Foldes, F.; et al. Comparison of immune activation of the COVID vaccines: ChAdOx1, BNT162b2, mRNA-1273, BBIBP-CorV, and Gam-COVID-Vac from serological human samples in Hungary showed higher protection after mRNA-based immunization. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 5297–5306. [Google Scholar] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Derhovanessian, E.; Vogler, I.; Kranz, L.M.; Vormehr, M.; Baum, A.; Pascal, K.; Quandt, J.; Maurus, D.; et al. COVID-19 vaccine BNT162b1 elicits human antibody and TH1 T cell responses. Nature 2020, 586, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef]
- Saciuk, Y.; Kertes, J.; Shamir Stein, N.; Ekka Zohar, A. Effectiveness of a Third Dose of BNT162b2 mRNA Vaccine. J. Infect. Dis. 2022, 225, 30–33. [Google Scholar] [CrossRef]
- Benotmane, I.; Gautier, G.; Perrin, P.; Olagne, J.; Cognard, N.; Fafi-Kremer, S.; Caillard, S. Antibody Response After a Third Dose of the mRNA-1273 SARS-CoV-2 Vaccine in Kidney Transplant Recipients with Minimal Serologic Response to 2 Doses. JAMA 2021, 326, 1063–1065. [Google Scholar] [CrossRef]
- Busa, R.; Sorrentino, M.C.; Russelli, G.; Amico, G.; Miceli, V.; Miele, M.; Di Bella, M.; Timoneri, F.; Gallo, A.; Zito, G.; et al. Specific Anti-SARS-CoV-2 Humoral and Cellular Immune Responses After Booster Dose of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine: Integrated Study of Adaptive Immune System Components. Front. Immunol. 2022, 13, 856657. [Google Scholar] [CrossRef]
- Ducloux, D.; Colladant, M.; Chabannes, M.; Yannaraki, M.; Courivaud, C. Humoral response after 3 doses of the BNT162b2 mRNA COVID-19 vaccine in patients on hemodialysis. Kidney Int. 2021, 100, 702–704. [Google Scholar] [CrossRef]
- Eliakim-Raz, N.; Leibovici-Weisman, Y.; Stemmer, A.; Ness, A.; Awwad, M.; Ghantous, N.; Stemmer, S.M. Antibody Titers Before and After a Third Dose of the SARS-CoV-2 BNT162b2 Vaccine in Adults Aged ≥60 Years. JAMA 2021, 326, 2203–2204. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, J.; Tonnus, W.; Paliege, A.; Rettig, R.; Steglich, A.; Gembardt, F.; Kessel, F.; Kroger, H.; Arndt, P.; Sradnick, J.; et al. Cellular and Humoral Immune Responses After 3 Doses of BNT162b2 mRNA SARS-CoV-2 Vaccine in Kidney Transplant. Transplantation 2021, 105, e267–e269. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, B.J.; Ellebedy, A.H. The germinal centre B cell response to SARS-CoV-2. Nat. Rev. Immunol. 2022, 22, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Rodriguez, R.; Valentin-Quiroga, J.; Avendano-Ortiz, J.; Martin-Quiros, A.; Pascual-Iglesias, A.; Terron-Arcos, V.; Montalban-Hernandez, K.; Casalvilla-Duenas, J.C.; Bergon-Gutierrez, M.; Alcami, J.; et al. Cellular and humoral functional responses after BNT162b2 mRNA vaccination differ longitudinally between naive and subjects recovered from COVID-19. Cell Rep. 2022, 38, 110235. [Google Scholar] [CrossRef] [PubMed]
- McMahan, K.; Yu, J.; Mercado, N.B.; Loos, C.; Tostanoski, L.H.; Chandrashekar, A.; Liu, J.; Peter, L.; Atyeo, C.; Zhu, A.; et al. Correlates of protection against SARS-CoV-2 in rhesus macaques. Nature 2021, 590, 630–634. [Google Scholar] [CrossRef]
- Pusnik, J.; Konig, J.; Mai, K.; Richter, E.; Zorn, J.; Proksch, H.; Schulte, B.; Alter, G.; Streeck, H. Persistent Maintenance of Intermediate Memory B Cells Following SARS-CoV-2 Infection and Vaccination Recall Response. J. Virol. 2022, 96, e0076022. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Bowman, K.A.; Stein, D.; Shin, S.; Ferbas, K.G.; Tobin, N.H.; Mann, C.; Fischinger, S.; Ollmann Saphire, E.; Lauffenburger, D.; Rimoin, A.W.; et al. Hybrid Immunity Shifts the Fc-Effector Quality of SARS-CoV-2 mRNA Vaccine-Induced Immunity. mBio 2022, 13, e0164722. [Google Scholar] [CrossRef]
- Ferrara, P.; Ponticelli, D.; Magliuolo, R.; Borrelli, M.; Schiavone, B.; Mantovani, L.G. Time-Varying Effect of Hybrid Immunity on the Risk of Breakthrough Infection after Booster Dose of mRNA COVID-19 Vaccine: The MOSAICO Study. Vaccines 2022, 10, 1353. [Google Scholar] [CrossRef]
- Kajanova, I.; Grossmannova, K.; Jelenska, L.; Lukacikova, L.; Radikova, Z.; Knutova, N.; Nahlikova, J.; Belisova, M.; Pastorekova, S.; Kopacek, J. Seroprevalence of SARS-CoV-2 antibodies in the county town of Slovakia—A pilot study from the Trencin city. Acta Virol. 2022, 66, 228–237. [Google Scholar] [CrossRef]
- Suryawanshi, R.; Ott, M. SARS-CoV-2 hybrid immunity: Silver bullet or silver lining? Nat. Rev. Immunol. 2022, 22, 591–592. [Google Scholar] [CrossRef]
- Moore, T.; Hossain, R.; Doores, K.J.; Shankar-Hari, M.; Fear, D.J. SARS-CoV-2-Specific Memory B Cell Responses Are Maintained After Recovery from Natural Infection and Postvaccination. Viral Immunol. 2022, 35, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Lenti, M.V.; Aronico, N.; Pellegrino, I.; Boveri, E.; Giuffrida, P.; Borrelli de Andreis, F.; Morbini, P.; Vanelli, L.; Pasini, A.; Ubezio, C.; et al. Depletion of circulating IgM memory B cells predicts unfavourable outcome in COVID-19. Sci. Rep. 2020, 10, 20836. [Google Scholar] [CrossRef] [PubMed]
- Rossi, C.M.; Lenti, M.V.; Merli, S.; Di Sabatino, A. Role of IgM Memory B Cells and Spleen Function in COVID-19. Front. Immunol. 2022, 13, 889876. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Busà, R.; Miele, M.; Sorrentino, M.C.; Amico, G.; Timoneri, F.; Miceli, V.; Di Bella, M.; Russelli, G.; Gallo, A.; Zito, G.; et al. Long-Term Effectiveness of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine on B Cell Compartment: Efficient Recall of SARS-CoV-2-Specific Memory B Cells. Int. J. Mol. Sci. 2022, 23, 15046. https://doi.org/10.3390/ijms232315046
Busà R, Miele M, Sorrentino MC, Amico G, Timoneri F, Miceli V, Di Bella M, Russelli G, Gallo A, Zito G, et al. Long-Term Effectiveness of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine on B Cell Compartment: Efficient Recall of SARS-CoV-2-Specific Memory B Cells. International Journal of Molecular Sciences. 2022; 23(23):15046. https://doi.org/10.3390/ijms232315046
Chicago/Turabian StyleBusà, Rosalia, Monica Miele, Maria Concetta Sorrentino, Giandomenico Amico, Francesca Timoneri, Vitale Miceli, Mariangela Di Bella, Giovanna Russelli, Alessia Gallo, Giovanni Zito, and et al. 2022. "Long-Term Effectiveness of BNT162b2 Pfizer-BioNTech mRNA-Based Vaccine on B Cell Compartment: Efficient Recall of SARS-CoV-2-Specific Memory B Cells" International Journal of Molecular Sciences 23, no. 23: 15046. https://doi.org/10.3390/ijms232315046