
D,L-Lysine-Acetylsalicylate + Glycine (LASAG) Reduces SARS-CoV-2 Replication and Shows an Additive Effect with Remdesivir

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
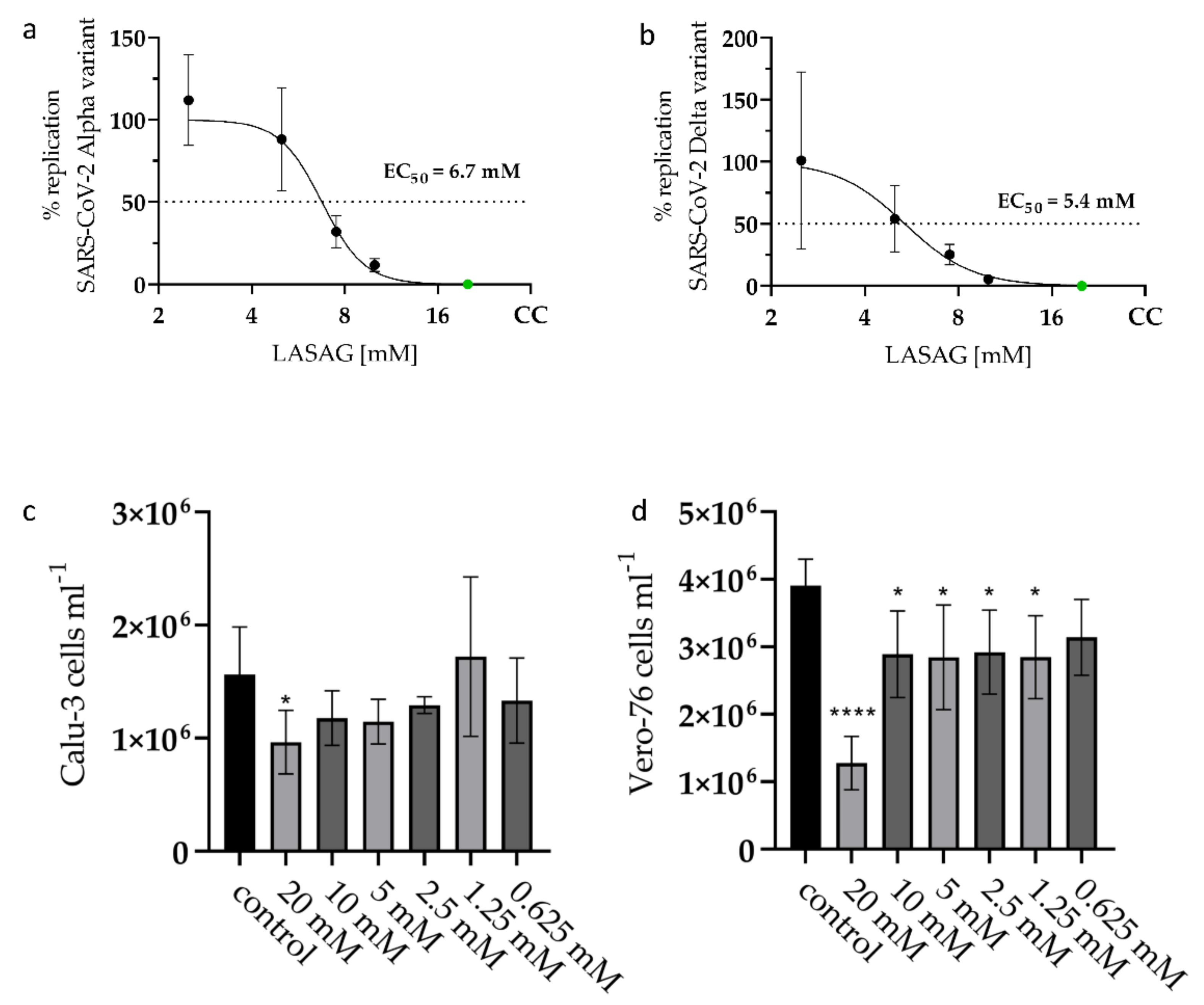
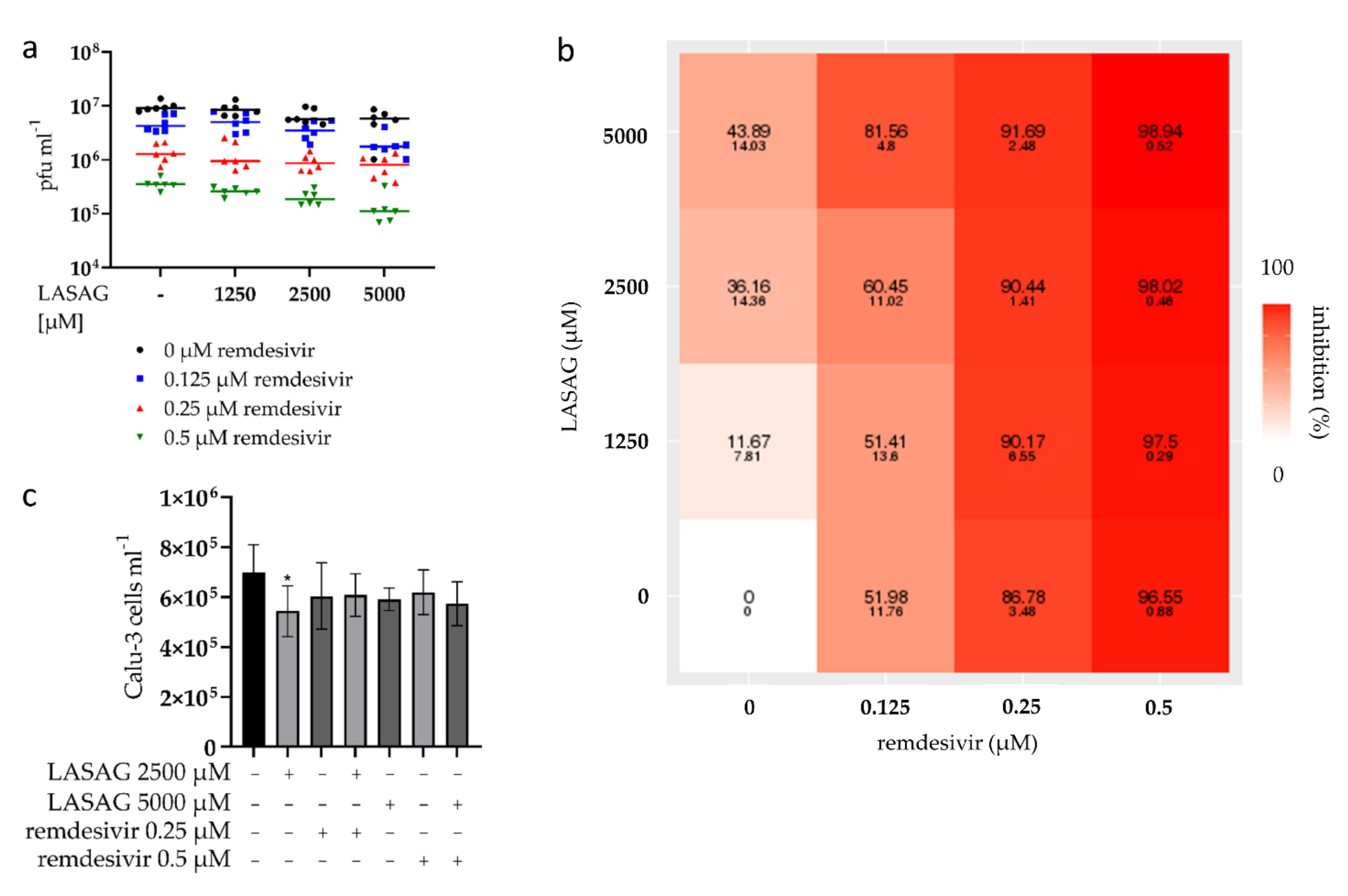
2.1. LASAG Inhibits SARS-CoV-2 Replication at Non-Toxic Concentrations In Vitro
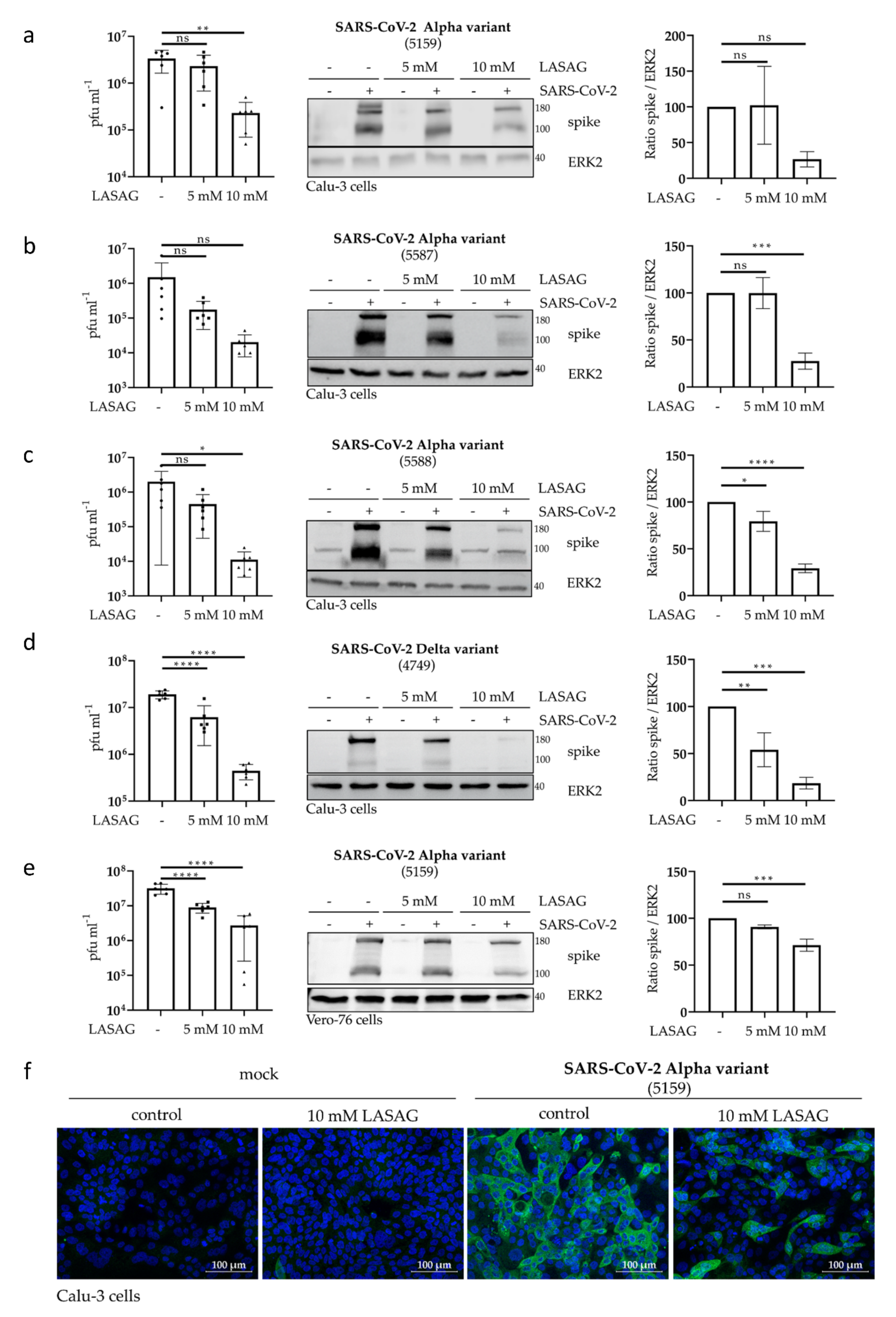
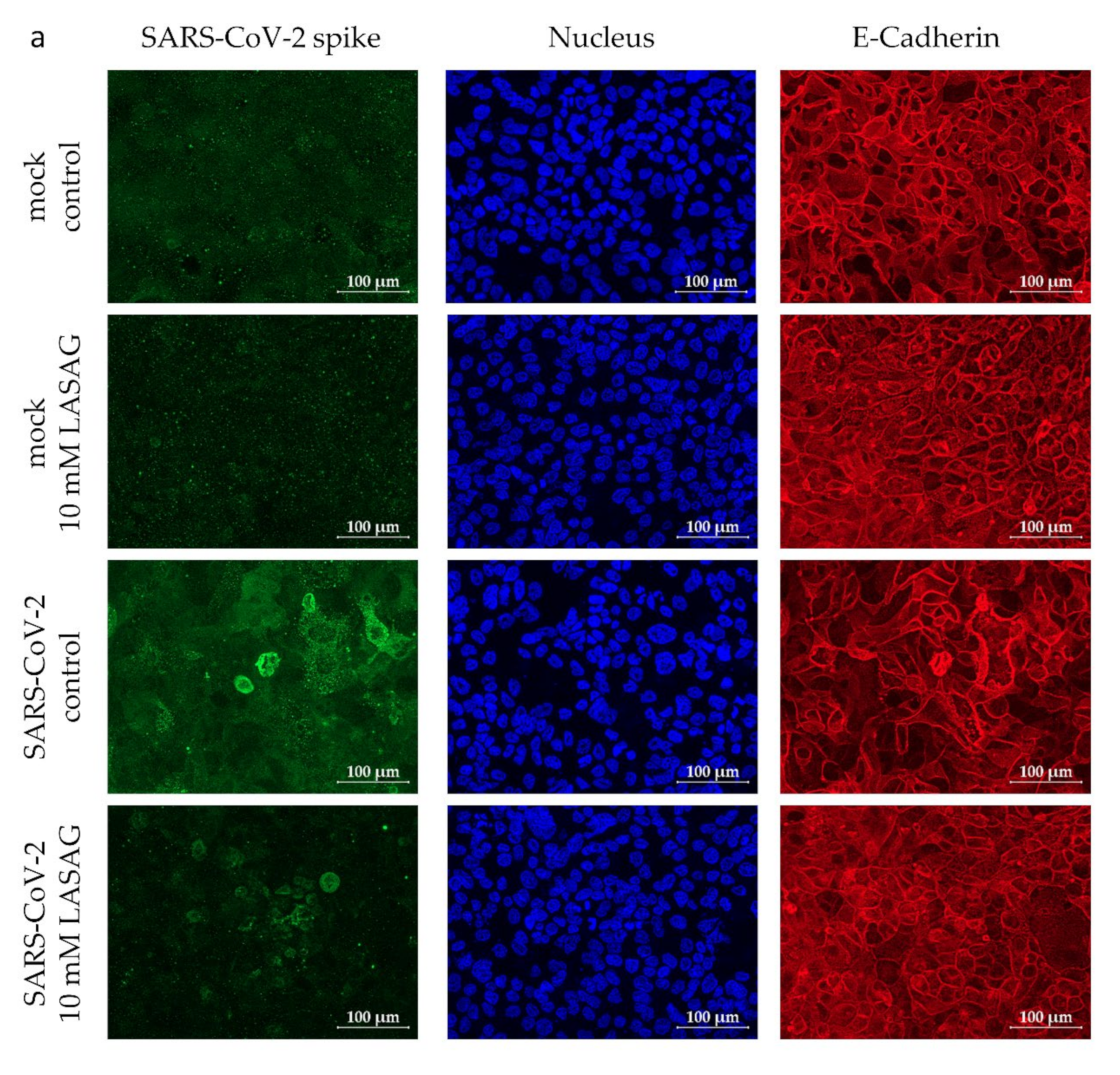
2.2. In the Presence of LASAG, the Replication of Different SARS-CoV-2 Variants Is Reduced In Vitro
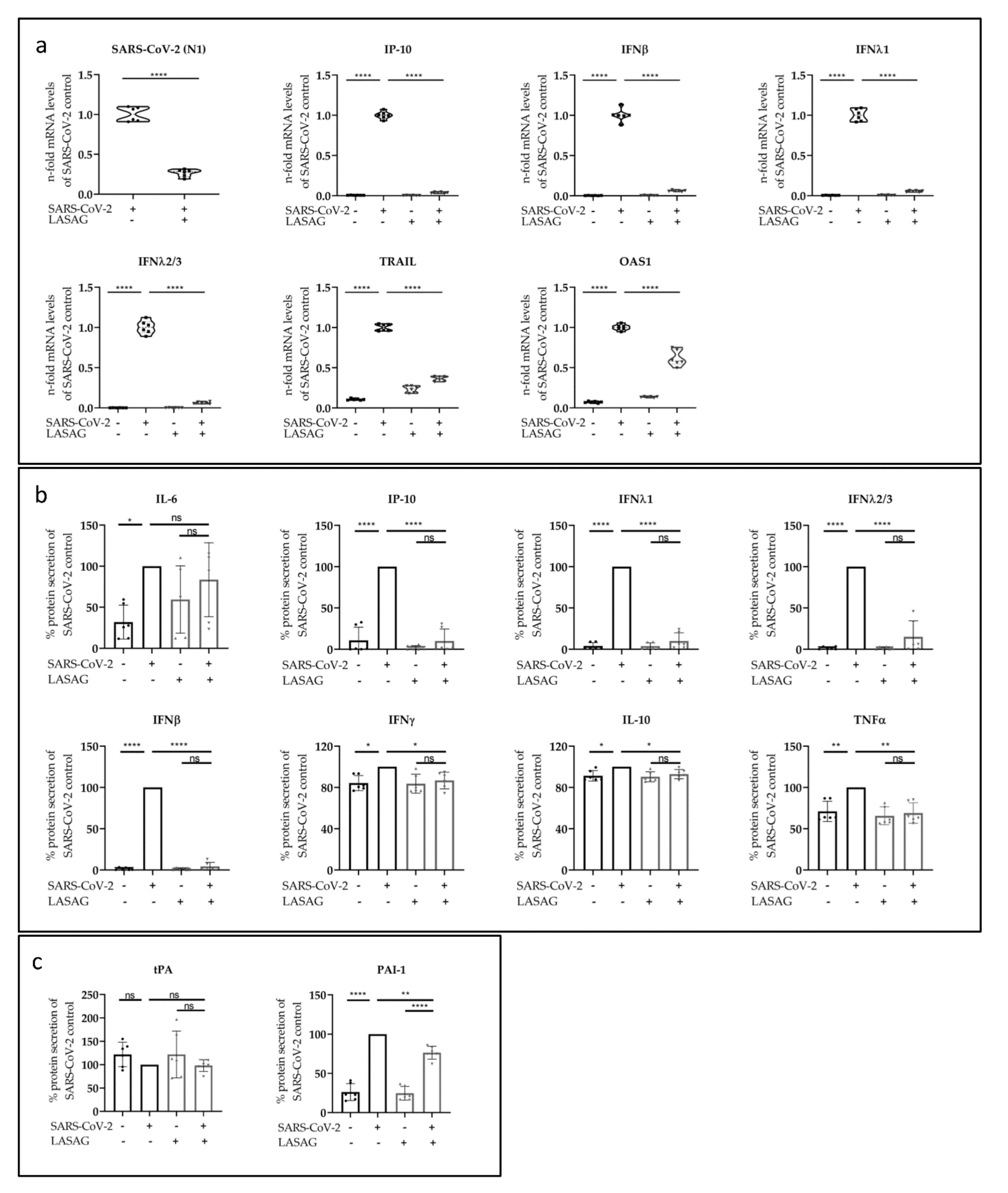
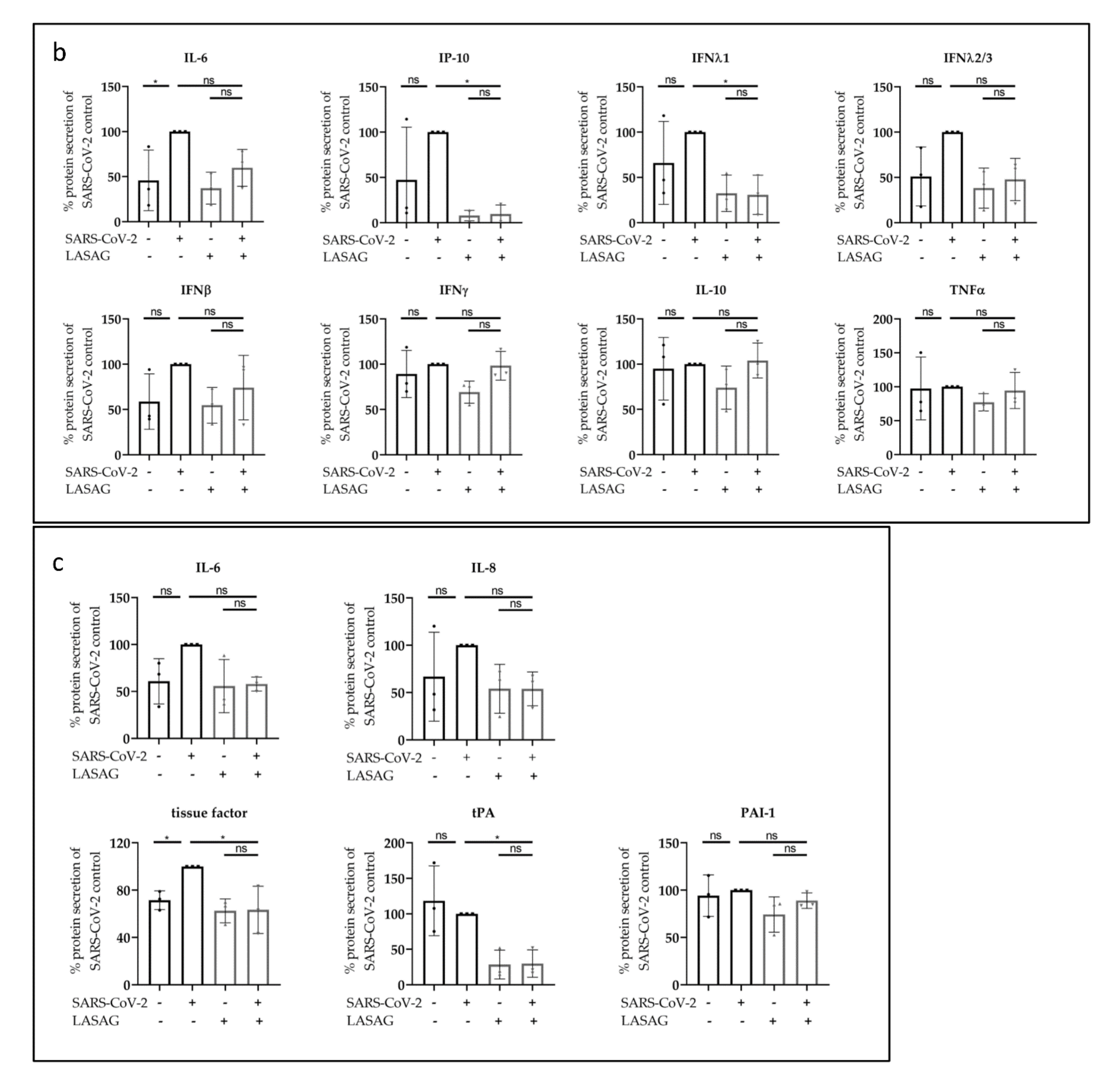
2.3. LASAG Reduces SARS-CoV-2-Induced Cytokine and Chemokine Production In Vitro
2.4. LASAG Reduces Cytokine and Chemokine Production of SARS-CoV-2-Infected Epithelial Cells within the Chip Model
2.5. SARS-CoV-2 Replication Is Strongly Inhibited upon Combinatory Treatment with LASAG and Remdesivir, Which Shows an Additive Inhibitory Effect In Vitro
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Cytotoxicity and Virus Infection
4.2. Viruses and Standard Plaque Assay
4.3. Immunfluorescence Microscopy
4.4. Western Blot Analysis
4.5. Detection of mRNA Expression by Using qRT-PCR
4.6. Flow Cytometry Analyses
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paules, C.I.; Marston, H.D.; Fauci, A.S. Coronavirus Infections—More Than Just the Common Cold. JAMA 2020, 323, 707–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Consortium, C.-G.U.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Beasley, R.; Hills, T.; Kearns, N. Asthma and COVID-19: Preconceptions about Predisposition. Am. J. Respir. Crit. Care Med. 2021, 203, 799–801. [Google Scholar] [CrossRef] [PubMed]
- Bansal, M. Cardiovascular disease and COVID-19. Diabetes Metab. Syndr. 2020, 14, 247–250. [Google Scholar] [CrossRef]
- Zhou, Y.; Chi, J.; Lv, W.; Wang, Y. Obesity and diabetes as high-risk factors for severe coronavirus disease 2019 (COVID-19). Diabetes Metab. Res. Rev. 2021, 37, e3377. [Google Scholar] [CrossRef]
- Grant, M.C.; Geoghegan, L.; Arbyn, M.; Mohammed, Z.; McGuinness, L.; Clarke, E.L.; Wade, R.G. The prevalence of symptoms in 24,410 adults infected by the novel coronavirus (SARS-CoV-2; COVID-19): A systematic review and meta-analysis of 148 studies from 9 countries. PLoS ONE 2020, 15, e0234765. [Google Scholar] [CrossRef]
- Gautret, P.; Million, M.; Jarrot, P.A.; Camoin-Jau, L.; Colson, P.; Fenollar, F.; Leone, M.; La Scola, B.; Devaux, C.; Gaubert, J.Y.; et al. Natural history of COVID-19 and therapeutic options. Expert Rev. Clin. Immunol. 2020, 16, 1159–1184. [Google Scholar] [CrossRef]
- Dos Santos, W.G. Natural history of COVID-19 and current knowledge on treatment therapeutic options. Biomed. Pharmacother. 2020, 129, 110493. [Google Scholar] [CrossRef]
- Cui, S.; Chen, S.; Li, X.; Liu, S.; Wang, F. Prevalence of venous thromboembolism in patients with severe novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 1421–1424. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.A.M.P.J.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Incidence of thrombotic complications in critically ill ICU patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef]
- Al-Samkari, H.; Leaf, R.S.K.; Dzik, W.H.; Carlson, J.C.T.; Fogerty, A.E.; Waheed, A.; Goodarzi, K.; Bendapudi, P.K.; Bornikova, L.; Gupta, S.; et al. COVID-19 and coagulation: Bleeding and thrombotic manifestations of SARS-CoV-2 infection. Blood 2020, 136, 489–500. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Deinhardt-Emmer, S.; Wittschieber, D.; Sanft, J.; Kleemann, S.; Elschner, S.; Haupt, K.F.; Vau, V.; Haring, C.; Rodel, J.; Henke, A.; et al. Early postmortem mapping of SARS-CoV-2 RNA in patients with COVID-19 and the correlation with tissue damage. eLife 2021, 10, e60361. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Talevi, A.; Bellera, C.L. Challenges and opportunities with drug repurposing: Finding strategies to find alternative uses of therapeutics. Expert Opin. Drug Discov. 2020, 15, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Scavone, C.; Mascolo, A.; Rafaniello, C.; Sportiello, L.; Trama, U.; Zoccoli, A.; Bernardi, F.F.; Racagni, G.; Berrino, L.; Castaldo, G.; et al. Therapeutic strategies to fight COVID-19: Which is the status artis? Br. J. Pharmacol. 2021, 179, 2128–2148. [Google Scholar] [CrossRef]
- Van de Veerdonk, F.L.; Giamarellos-Bourboulis, E.; Pickkers, P.; Derde, L.; Leavis, H.; van Crevel, R.; Engel, J.J.; Wiersinga, W.J.; Vlaar, A.P.J.; Shankar-Hari, M.; et al. A guide to immunotherapy for COVID-19. Nat. Med. 2022, 28, 39–50. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of Covid-19—Final Report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Ansems, K.; Grundeis, F.; Dahms, K.; Mikolajewska, A.; Thieme, V.; Piechotta, V.; Metzendorf, M.I.; Stegemann, M.; Benstoem, C.; Fichtner, F. Remdesivir for the treatment of COVID-19. Cochrane Database Syst. Rev. 2021, 8, CD014962. [Google Scholar] [CrossRef]
- Fischer, W.A.; Eron, J.J., Jr.; Holman, W.; Cohen, M.S.; Fang, L.; Szewczyk, L.J.; Sheahan, T.P.; Baric, R.; Mollan, K.R.; Wolfe, C.R.; et al. A phase 2a clinical trial of molnupiravir in patients with COVID-19 shows accelerated SARS-CoV-2 RNA clearance and elimination of infectious virus. Sci. Transl. Med. 2022, 14, eabl7430. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, Y.; Huang, J.; Yin, P.; Cheng, Z.; Wu, J.; Chen, S.; Zhang, Y.; Chen, B.; Lu, M.; et al. Favipiravir Versus Arbidol for Clinical Recovery Rate in Moderate and Severe Adult COVID-19 Patients: A Prospective, Multicenter, Open-Label, Randomized Controlled Clinical Trial. Front. Pharmacol. 2021, 12, 683296. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Yang, M.; Liu, D.; Chen, J.; Shu, D.; Xia, J.; Liao, X.; Gu, Y.; Cai, Q.; Yang, Y.; et al. Experimental Treatment with Favipiravir for COVID-19: An Open-Label Control Study. Engineering 2020, 6, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Lamb, Y.N. Nirmatrelvir Plus Ritonavir: First Approval. Drugs 2022, 82, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhao, Y.; Zhang, F.; Wang, Q.; Li, T.; Liu, Z.; Wang, J.; Qin, Y.; Zhang, X.; Yan, X.; et al. The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The Perspectives of clinical immunologists from China. Clin. Immunol. 2020, 214, 108393. [Google Scholar] [CrossRef]
- Luo, P.; Liu, Y.; Qiu, L.; Liu, X.; Liu, D.; Li, J. Tocilizumab treatment in COVID-19: A single center experience. J. Med. Virol. 2020, 92, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef]
- Deeks, E.D. Casirivimab/Imdevimab: First Approval. Drugs 2021, 81, 2047–2055. [Google Scholar] [CrossRef]
- Syed, Y.Y. Regdanvimab: First Approval. Drugs 2021, 81, 2133–2137. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Compound Summary Aspirin. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Aspirin (accessed on 16 March 2022).
- Christiansen, M.; Grove, E.L.; Hvas, A.M. Contemporary Clinical Use of Aspirin: Mechanisms of Action, Current Concepts, Unresolved Questions, and Future Perspectives. Semin. Thromb. Hemost. 2021, 47, 800–814. [Google Scholar] [CrossRef]
- Scheuch, G.; Canisius, S.; Nocker, K.; Hofmann, T.; Naumann, R.; Pleschka, S.; Ludwig, S.; Welte, T.; Planz, O. Targeting intracellular signaling as an antiviral strategy: Aerosolized LASAG for the treatment of influenza in hospitalized patients. Emerg. Microbes Infect. 2018, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Botting, R.M. Mechanism of action of aspirin-like drugs. Semin. Arthritis Rheum. 1997, 26, 2–10. [Google Scholar] [CrossRef]
- Wilden, J.J.; van Kruchten, A.; Gieselmann, L.; Hrincius, E.R.; Deinhardt-Emmer, S.; Haupt, K.F.; Preugschas, H.F.; Niemann, S.; Ludwig, S.; Ehrhardt, C. The influenza replication blocking inhibitor LASAG does not sensitize human epithelial cells for bacterial infections. PLoS ONE 2020, 15, e0233052. [Google Scholar] [CrossRef]
- Müller, C.; Karl, N.; Ziebuhr, J.; Pleschka, S. D, L-lysine acetylsalicylate + glycine Impairs Coronavirus Replication. J. Antivir. Antiretrovir. 2016, 8, 4. [Google Scholar] [CrossRef]
- Rübsamen-Schaeff, H.; Buschmann, H.; Mannhold, R.; Holenz, J. New Drug Development for Known and Emerging Viruses; Wiley: Hoboken, NJ, USA, 2022. [Google Scholar]
- Mazur, I.; Wurzer, W.J.; Ehrhardt, C.; Pleschka, S.; Puthavathana, P.; Silberzahn, T.; Wolff, T.; Planz, O.; Ludwig, S. Acetylsalicylic acid (ASA) blocks influenza virus propagation via its NF-kappaB-inhibiting activity. Cell Microbiol. 2007, 9, 1683–1694. [Google Scholar] [CrossRef]
- Droebner, K.; Haasbach, E.; Dudek, S.E.; Scheuch, G.; Nocker, K.; Canisius, S.; Ehrhardt, C.; von Degenfeld, G.; Ludwig, S.; Planz, O. Pharmacodynamics, Pharmacokinetics, and Antiviral Activity of BAY 81-8781, a Novel NF-kappaB Inhibiting Anti-influenza Drug. Front. Microbiol. 2017, 8, 2130. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.H.; Khanna, A.K.; Kethireddy, S.; Yamane, D.; Levine, A.; Jackson, A.M.; McCurdy, M.T.; Tabatabai, A.; Kumar, G.; Park, P.; et al. Aspirin Use Is Associated with Decreased Mechanical Ventilation, Intensive Care Unit Admission, and In-Hospital Mortality in Hospitalized Patients with Coronavirus Disease 2019. Anesth. Analg. 2021, 132, 930–941. [Google Scholar] [CrossRef]
- Sisinni, A.; Rossi, L.; Battista, A.; Poletti, E.; Battista, F.; Battista, R.A.; Malagoli, A.; Biagi, A.; Zanni, A.; Sticozzi, C.; et al. Pre-admission acetylsalicylic acid therapy and impact on in-hospital outcome in COVID-19 patients: The ASA-CARE study. Int. J. Cardiol. 2021, 344, 240–245. [Google Scholar] [CrossRef]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef]
- Morris, T.; Stables, M.; Hobbs, A.; de Souza, P.; Colville-Nash, P.; Warner, T.; Newson, J.; Bellingan, G.; Gilroy, D.W. Effects of low-dose aspirin on acute inflammatory responses in humans. J. Immunol. 2009, 183, 2089–2096. [Google Scholar] [CrossRef]
- FitzGerald, E.S.; Chen, Y.; Fitzgerald, K.A.; Jamieson, A.M. Lung Epithelial Cell Transcriptional Regulation as a Factor in COVID-19-associated Coagulopathies. Am. J. Respir. Cell Mol. Biol. 2021, 64, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Deinhardt-Emmer, S.; Böttcher, S.; Häring, C.; Giebeler, L.; Henke, A.; Zell, R.; Jungwirth, J.; Jordan Paul, M.; Werz, O.; Hornung, F.; et al. SARS-CoV-2 Causes Severe Epithelial Inflammation and Barrier Dysfunction. J. Virol. 2021, 95, e00110-21. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Giri, A.K.; Aittokallio, T. SynergyFinder 2.0: Visual analytics of multi-drug combination synergies. Nucleic Acids Res. 2020, 48, W488–W493. [Google Scholar] [CrossRef] [PubMed]
- Siwik, D.; Gajewska, M.; Karon, K.; Pluta, K.; Wondolkowski, M.; Wilimski, R.; Szarpak, L.; Filipiak, K.J.; Gasecka, A. Pleiotropic Effects of Acetylsalicylic Acid after Coronary Artery Bypass Grafting-Beyond Platelet Inhibition. J. Clin. Med. 2021, 10, 2317. [Google Scholar] [CrossRef] [PubMed]
- Schror, K. Aspirin and Reye syndrome: A review of the evidence. Paediatr. Drugs 2007, 9, 195–204. [Google Scholar] [CrossRef]
- Group, R.C. Aspirin in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2022, 399, 143–151. [Google Scholar] [CrossRef]
- Husain, A.; Sayem, M.A.; Kamal, S.M.; Morshed, M.E. Beneficial Effect of Low Dose Aspirin in Adult Patients with COVID-19: A Retrospective Observational Study in Bangladesh. Mymensingh Med. J. 2022, 31, 194–199. [Google Scholar]
- Russo, V.; Di Maio, M.; Attena, E.; Silverio, A.; Scudiero, F.; Celentani, D.; Lodigiani, C.; Di Micco, P. Clinical impact of pre-admission antithrombotic therapy in hospitalized patients with COVID-19: A multicenter observational study. Pharmacol. Res. 2020, 159, 104965. [Google Scholar] [CrossRef]
- Pan, D.; Ip, A.; Zhan, S.; Wasserman, I.; Snyder, D.J.; Agathis, A.Z.; Shamapant, N.; Yang, J.Y.; Pai, A.; Mazumdar, M.; et al. Pre-hospital antiplatelet medication use on COVID-19 disease severity. Heart Lung 2021, 50, 618–621. [Google Scholar] [CrossRef]
- Kim, I.; Yoon, S.; Kim, M.; Lee, H.; Park, S.; Kim, W.; Lee, S. Aspirin Is Related to Worse Clinical Outcomes of COVID-19. Medicina 2021, 57, 931. [Google Scholar] [CrossRef]
- Vahedian-Azimi, A.; Rahimibashar, F.; Najafi, A.; Kidde, J.; Shahriary, A.; Shojaei, S.; Pourhoseingholi, M.A.; Jamialahmadi, T.; Sahebkar, A. Associastion of In-hospital Use of Statins, Aspirin, and Renin-Angiotensin-Aldosterone Inhibitors with Mortality and ICU Admission Due to COVID-19. Adv. Exp. Med. Biol. 2021, 1327, 205–214. [Google Scholar] [CrossRef]
- Mura, C.; Preissner, S.; Nahles, S.; Heiland, M.; Bourne, P.E.; Preissner, R. Real-world evidence for improved outcomes with histamine antagonists and aspirin in 22,560 COVID-19 patients. Signal Transduct. Target. Ther. 2021, 6, 267. [Google Scholar] [CrossRef]
- Ehrhardt, C.; Ruckle, A.; Hrincius, E.R.; Haasbach, E.; Anhlan, D.; Ahmann, K.; Banning, C.; Reiling, S.J.; Kuhn, J.; Strobl, S.; et al. The NF-kappaB inhibitor SC75741 efficiently blocks influenza virus propagation and confers a high barrier for development of viral resistance. Cell Microbiol. 2013, 15, 1198–1211. [Google Scholar] [CrossRef]
- Li, Q.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-kappaB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [Green Version]
- Hariharan, A.; Hakeem, A.R.; Radhakrishnan, S.; Reddy, M.S.; Rela, M. The Role and Therapeutic Potential of NF-kappa-B Pathway in Severe COVID-19 Patients. Inflammopharmacology 2021, 29, 91–100. [Google Scholar] [CrossRef]
- Temple, A.R. Acute and chronic effects of aspirin toxicity and their treatment. Arch. Intern. Med. 1981, 141, 364–369. [Google Scholar] [CrossRef]
- Nagelschmitz, J.; Blunck, M.; Kraetzschmar, J.; Ludwig, M.; Wensing, G.; Hohlfeld, T. Pharmacokinetics and pharmacodynamics of acetylsalicylic acid after intravenous and oral administration to healthy volunteers. Clin. Pharmacol. 2014, 6, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Bianco, S.; Vaghi, A.; Robuschi, M.; Refini, R.M.; Pieroni, M.G.; Sestini, P. Steroid-sparing effect of inhaled lysine acetylsalicylate and furosemide in high-dose beclomethasone-dependent asthma. J. Allergy Clin. Immunol. 1995, 95, 937–943. [Google Scholar] [CrossRef]
- Wang, X.; Sacramento, C.Q.; Jockusch, S.; Chaves, O.A.; Tao, C.; Fintelman-Rodrigues, N.; Chien, M.; Temerozo, J.R.; Li, X.; Kumar, S.; et al. Combination of antiviral drugs inhibits SARS-CoV-2 polymerase and exonuclease and demonstrates COVID-19 therapeutic potential in viral cell culture. Commun. Biol. 2022, 5, 154. [Google Scholar] [CrossRef]
- Jeffreys, L.N.; Pennington, S.H.; Duggan, J.; Caygill, C.H.; Lopeman, R.C.; Breen, A.F.; Jinks, J.B.; Ardrey, A.; Donnellan, S.; Patterson, E.I.; et al. Remdesivir-ivermectin combination displays synergistic interaction with improved in vitro activity against SARS-CoV-2. Int. J. Antimicrob. Agents 2022, 59, 106542. [Google Scholar] [CrossRef] [PubMed]
- Ianevski, A.; Yao, R.; Zusinaite, E.; Lello, L.S.; Wang, S.; Jo, E.; Yang, J.; Ravlo, E.; Wang, W.; Lysvand, H.; et al. Synergistic Interferon-Alpha-Based Combinations for Treatment of SARS-CoV-2 and Other Viral Infections. Viruses 2021, 13, 2489. [Google Scholar] [CrossRef] [PubMed]
- Iba, T.; Levy, J.H.; Levi, M.; Connors, J.M.; Thachil, J. Coagulopathy of Coronavirus Disease 2019. Crit. Care Med. 2020, 48, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Fraisse, M.; Logre, E.; Pajot, O.; Mentec, H.; Plantefeve, G.; Contou, D. Thrombotic and hemorrhagic events in critically ill COVID-19 patients: A French monocenter retrospective study. Crit. Care 2020, 24, 275. [Google Scholar] [CrossRef] [PubMed]
- Mackman, N. Role of tissue factor in hemostasis, thrombosis, and vascular development. Arter. Thromb. Vasc. Biol. 2004, 24, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Canas, C.A.; Canas, F.; Bautista-Vargas, M.; Bonilla-Abadia, F. Role of Tissue Factor in the Pathogenesis of COVID-19 and the Possible Ways to Inhibit It. Clin. Appl. Thromb. Hemost. 2021, 27, 10760296211003983. [Google Scholar] [CrossRef] [PubMed]
- DiNicolantonio, J.J.; McCarty, M. Thrombotic complications of COVID-19 may reflect an upregulation of endothelial tissue factor expression that is contingent on activation of endosomal NADPH oxidase. Open Heart 2020, 7, e001337. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Vargas, M.; Bonilla-Abadia, F.; Canas, C.A. Potential role for tissue factor in the pathogenesis of hypercoagulability associated with in COVID-19. J. Thromb. Thrombolysis 2020, 50, 479–483. [Google Scholar] [CrossRef]
- Mackman, N.; Antoniak, S.; Wolberg, A.S.; Kasthuri, R.; Key, N.S. Coagulation Abnormalities and Thrombosis in Patients Infected with SARS-CoV-2 and Other Pandemic Viruses. Arter. Thromb. Vasc. Biol. 2020, 40, 2033–2044. [Google Scholar] [CrossRef]
- Salyers, A.K.; Szalony, J.A.; Suleymanov, O.D.; Parlow, J.J.; Wood, R.S.; South, M.S.; Nicholson, N.S. Assessment of bleeding propensity in non-human primates by combination of selective tissue factor/VIIa inhibition and aspirin compared to warfarin and aspirin treatment. Pharmacology 2004, 70, 100–106. [Google Scholar] [CrossRef]
- Roque, M.; Reis, E.D.; Fuster, V.; Padurean, A.; Fallon, J.T.; Taubman, M.B.; Chesebro, J.H.; Badimon, J.J. Inhibition of tissue factor reduces thrombus formation and intimal hyperplasia after porcine coronary angioplasty. J. Am. Coll. Cardiol. 2000, 36, 2303–2310. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.K.; Vaidyula, V.R.; Bagga, S.; Jalagadugula, G.; Gaughan, J.; Wilhite, D.B.; Comerota, A.J. Effect of antiplatelet agents clopidogrel, aspirin, and cilostazol on circulating tissue factor procoagulant activity in patients with peripheral arterial disease. Thromb. Haemost. 2006, 96, 738–743. [Google Scholar] [PubMed]
- Marco, J.; Ariens, R.A.; Fajadet, J.; Bossi, I.M.; Marco, I.; Bernies, M.; Romano, S.M.; Donatelli, F.; Brambilla, G.M.; Somalvico, F.; et al. Effect of aspirin and ticlopidine on plasma tissue factor levels in stable and unstable angina pectoris. Am. J. Cardiol. 2000, 85, 527–531. [Google Scholar] [CrossRef]
- Subrahmanian, S.; Borczuk, A.; Salvatore, S.; Fung, K.M.; Merrill, J.T.; Laurence, J.; Ahamed, J. Tissue factor upregulation is associated with SARS-CoV-2 in the lungs of COVID-19 patients. J. Thromb. Haemost. 2021, 19, 2268–2274. [Google Scholar] [CrossRef]
- Glowacka, P.; Rudnicka, L.; Warszawik-Hendzel, O.; Sikora, M.; Goldust, M.; Gajda, P.; Stochmal, A.; Blicharz, L.; Rakowska, A.; Olszewska, M. The Antiviral Properties of Cyclosporine. Focus on Coronavirus, Hepatitis C Virus, Influenza Virus, and Human Immunodeficiency Virus Infections. Biology 2020, 9, 192. [Google Scholar] [CrossRef]
- Ghasemnejad-Berenji, M.; Pashapour, S.; Sadeghpour, S. Pentoxifylline: A Drug with Antiviral and Anti-Inflammatory Effects to Be Considered in the Treatment of Coronavirus Disease 2019. Med. Princ. Pract. 2021, 30, 98–100. [Google Scholar] [CrossRef]
- Gale, A.J. Continuing education course #2: Current understanding of hemostasis. Toxicol. Pathol. 2011, 39, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Warnock, M.; Harbaugh, A.; Yalavarthi, S.; Gockman, K.; Zuo, M.; Madison, J.A.; Knight, J.S.; Kanthi, Y.; Lawrence, D.A. Plasma tissue plasminogen activator and plasminogen activator inhibitor-1 in hospitalized COVID-19 patients. Sci. Rep. 2021, 11, 1580. [Google Scholar] [CrossRef]
- Levin, R.I.; Harpel, P.C.; Harpel, J.G.; Recht, P.A. Inhibition of tissue plasminogen activator activity by aspirin in vivo and its relationship to levels of tissue plasminogen activator inhibitor antigen, plasminogen activator and their complexes. Blood 1989, 74, 1635–1643. [Google Scholar] [CrossRef] [Green Version]
- Deinhardt-Emmer, S.; Rennert, K.; Schicke, E.; Cseresnyes, Z.; Windolph, M.; Nietzsche, S.; Heller, R.; Siwczak, F.; Haupt, K.F.; Carlstedt, S.; et al. Co-infection with Staphylococcus aureus after primary influenza virus infection leads to damage of the endothelium in a human alveolus-on-a-chip model. Biofabrication 2020, 12, 025012. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence 5′-3′ |
|---|---|
| GAPDH-fwd | CTCTGCTCCTCCTGTTCGAC |
| GAPDH-rev | CAATACGACCAAATCCGTTGAC |
| 2019_nCoV_N1-fwd | GACCCCAAAATCAGCGAAAT |
| 2019_nCoV_N1-rev | TCTGGTTACTGCCAGTTGAATCTG |
| IP-10-fwd | CCAGAATCGAAGGCCATCAA |
| IP-10-rev | TTTCCTTGCTAACTGCTTTCAG |
| IFN-β-fwd | ATGACCAACAAGTGTCTCCTCC |
| IFN-β-rev | GGAATCCAAGCAAGTTGTAGCTC |
| IFN-λ1-fwd | CGCCTTGGAAGAGTCACTCA |
| IFN-λ1-rev | GAAGCCTCAGGTCCCAATTC |
| IFN-λ2/3-fwd | AGTTCCGGGCCTGTATCCAG |
| IFN-λ2/3-rev | GAGCCGGTACAGCCAATGGT |
| TRAIL-fwd | GTCTCTCTGTGTGGCTGTAACTTACG |
| TRAIL-rev | AAACAAGCAATGCCACTTTTGG |
| OAS1-fwd | GATCTCAGAAATACCCCAGCCA |
| OAS1-rev | AGCTACCTCGGAAGCACCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jungwirth, J.; Häring, C.; König, S.; Giebeler, L.; Doshi, H.; Brandt, C.; Deinhardt-Emmer, S.; Löffler, B.; Ehrhardt, C. D,L-Lysine-Acetylsalicylate + Glycine (LASAG) Reduces SARS-CoV-2 Replication and Shows an Additive Effect with Remdesivir. Int. J. Mol. Sci. 2022, 23, 6880. https://doi.org/10.3390/ijms23136880
Jungwirth J, Häring C, König S, Giebeler L, Doshi H, Brandt C, Deinhardt-Emmer S, Löffler B, Ehrhardt C. D,L-Lysine-Acetylsalicylate + Glycine (LASAG) Reduces SARS-CoV-2 Replication and Shows an Additive Effect with Remdesivir. International Journal of Molecular Sciences. 2022; 23(13):6880. https://doi.org/10.3390/ijms23136880
Chicago/Turabian StyleJungwirth, Johannes, Clio Häring, Sarah König, Liane Giebeler, Heena Doshi, Christian Brandt, Stefanie Deinhardt-Emmer, Bettina Löffler, and Christina Ehrhardt. 2022. "D,L-Lysine-Acetylsalicylate + Glycine (LASAG) Reduces SARS-CoV-2 Replication and Shows an Additive Effect with Remdesivir" International Journal of Molecular Sciences 23, no. 13: 6880. https://doi.org/10.3390/ijms23136880