
Differential Immune Checkpoint and Ig-like V-Type Receptor Profiles in COVID-19: Associations with Severity and Treatment

 , ,
, ,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Recruitment and Study Design
2.2. Immunophenotyping
2.3. Plasma and PBMCs Isolation
2.4. Cytokines, Soluble Immune-Checkpoints and Thrombosis Markers Quantification
2.5. Statistical Analysis
3. Results
3.1. Parameters at Admission Associated to COVID-19 Severity
3.1.1. Patients’ Clinical Characteristics and Routine Laboratory Markers
3.1.2. Soluble Plasma Markers
3.1.3. Immune Cell Subpopulations and Their Immune Checkpoint Expression
3.2. Parameters at Admission Associated with Secondary Infections in COVID-19 Patients
3.2.1. Patients’ Clinical Characteristics and Routine Laboratory Markers at Admission Associated with Secondary Infections
3.2.2. Soluble Plasma Markers at Admission Associated with Secondary Infections
3.2.3. Immune Cell Subpopulations and Their Immune-Checkpoint Expression at Admission Associated with Secondary Infections
3.3. Immunological Profile Changes after Secondary Infections in COVID-19 Patients
3.4. Immunological Profile Changes after Corticosteroid Treatment in COVID-19 Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group. Association between Administration of Systemic Corticosteroids and Mortality among Critically Ill Patients with COVID-19: A Meta-Analysis. JAMA 2020, 324, 1330–1341. [Google Scholar] [CrossRef] [PubMed]
- RECOVERY Collaborative Group; Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; et al. Dexamethasone in Hospitalized Patients with COVID-19—Preliminary Report. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef]
- De Bruyn, A.; Verellen, S.; Bruckers, L.; Geebelen, L.; Callebaut, I.; De Pauw, I.; Stessel, B.; Dubois, J. Secondary Infection in COVID-19 Critically Ill Patients: A Retrospective Single-Center Evaluation. BMC Infect. Dis. 2022, 22, 207. [Google Scholar] [CrossRef] [PubMed]
- Suarez-de-la-Rica, A.; Serrano, P.; De-la-Oliva, R.; Sánchez-Díaz, P.; Molinero, P.; Falces-Romero, I.; Ferrando, C.; Rello, J.; Maseda, E. Secondary Infections in Mechanically Ventilated Patients with COVID-19: An Overlooked Matter? Rev. Esp. Quim. 2021, 34, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Bellinvia, S.; Edwards, C.J.; Schisano, M.; Banfi, P.; Fallico, M.; Murabito, P. The Unleashing of the Immune System in COVID-19 and Sepsis: The Calm before the Storm? Inflamm. Res. 2020, 69, 757–763. [Google Scholar] [CrossRef]
- Avendaño-Ortiz, J.; Lozano-Rodríguez, R.; Martín-Quirós, A.; Maroun-Eid, C.; Terrón-Arcos, V.; Montalbán-Hernández, K.; Valentín, J.; del Val, E.M.; García-Garrido, M.A.; del Balzo-Castillo, Á.; et al. SARS-CoV-2 Proteins Induce Endotoxin Tolerance Hallmarks: A Demonstration in Patients with COVID-19. J. Immunol. 2021, 207, 162–174. [Google Scholar] [CrossRef]
- López-Collazo, E.; Avendaño-Ortiz, J.; Martín-Quirós, A.; Aguirre, L.A. Immune Response and COVID-19: A Mirror Image of Sepsis. Int. J. Biol. Sci. 2020, 16, 2479–2489. [Google Scholar] [CrossRef]
- Shafran, N.; Shafran, I.; Ben-Zvi, H.; Sofer, S.; Sheena, L.; Krause, I.; Shlomai, A.; Goldberg, E.; Sklan, E.H. Secondary Bacterial Infection in COVID-19 Patients Is a Stronger Predictor for Death Compared to Influenza Patients. Sci. Rep. 2021, 11, 12703. [Google Scholar] [CrossRef]
- Liu, Y.; Pang, Y.; Hu, Z.; Wu, M.; Wang, C.; Feng, Z.; Mao, C.; Tan, Y.; Liu, Y.; Chen, L.; et al. Thymosin Alpha 1 (Tα1) Reduces the Mortality of Severe COVID-19 by Restoration of Lymphocytopenia and Reversion of Exhausted T Cells. Clin. Infect. Dis. 2020, 71, 2150–2157. [Google Scholar] [CrossRef]
- Zhou, R.; To, K.K.-W.; Wong, Y.-C.; Liu, L.; Zhou, B.; Li, X.; Huang, H.; Mo, Y.; Luk, T.-Y.; Lau, T.T.-K.; et al. Acute SARS-CoV-2 Infection Impairs Dendritic Cell and T Cell Responses. Immunity 2020, 53, 864–877.e5. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.-E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000.e3. [Google Scholar] [CrossRef] [PubMed]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An Immune-Based Biomarker Signature Is Associated with Mortality in COVID-19 Patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef] [PubMed]
- Avendaño-Ortiz, J.; Lozano-Rodríguez, R.; Martín-Quirós, A.; Terrón, V.; Maroun-Eid, C.; Montalbán-Hernández, K.; Valentín-Quiroga, J.; García-Garrido, M.Á.; del Val, E.M.; del Balzo-Castillo, Á.; et al. The Immune Checkpoints Storm in COVID-19: Role as Severity Markers at Emergency Department Admission. Clin. Transl. Med. 2021, 11, e573. [Google Scholar] [CrossRef] [PubMed]
- Hue, S.; Beldi-Ferchiou, A.; Bendib, I.; Surenaud, M.; Fourati, S.; Frapard, T.; Rivoal, S.; Razazi, K.; Carteaux, G.; Delfau-Larue, M.-H.; et al. Uncontrolled Innate and Impaired Adaptive Immune Responses in Patients with COVID-19 ARDS. Am. J. Respir. Crit. Care Med. 2020, 202, 1509–1519. [Google Scholar] [CrossRef]
- Avendaño-Ortiz, J.; Lozano-Rodríguez, R.; Martín-Quirós, A.; Maroun-Eid, C.; Terrón, V.; Valentín, J.; Montalbán-Hernández, K.; Ruiz de la Bastida, F.; García-Garrido, M.A.; Cubillos-Zapata, C.; et al. Proteins from SARS-CoV-2 Reduce T Cell Proliferation: A Mirror Image of Sepsis. Heliyon 2020, 6, e05635. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [Green Version]
- Bolandi, N.; Derakhshani, A.; Hemmat, N.; Baghbanzadeh, A.; Asadzadeh, Z.; Afrashteh Nour, M.; Brunetti, O.; Bernardini, R.; Silvestris, N.; Baradaran, B. The Positive and Negative Immunoregulatory Role of B7 Family: Promising Novel Targets in Gastric Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 10719. [Google Scholar] [CrossRef]
- Li, Y.-M.; Liu, Z.-Y.; Li, Z.-C.; Wang, J.-C.; Yu, J.-M.; Yang, H.-J.; Chen, Z.-N.; Tang, J. Alterations of the Immunologic Co-Stimulator B7 and TNFR Families Correlate with Hepatocellular Carcinoma Prognosis and Metastasis by Inactivating STAT3. Int. J. Mol. Sci. 2019, 20, 156. [Google Scholar] [CrossRef] [Green Version]
- Pascolutti, R.; Sun, X.; Kao, J.; Maute, R.; Ring, A.M.; Bowman, G.R.; Kruse, A.C. Structure and Dynamics of PD-L1 and an Ultra High-Affinity PD-1 Receptor Mutant. Structure 2016, 24, 1719–1728. [Google Scholar] [CrossRef] [Green Version]
- Vigdorovich, V.; Ramagopal, U.A.; Lázár-Molnár, E.; Sylvestre, E.; Lee, J.S.; Hofmeyer, K.A.; Zang, X.; Nathenson, S.G.; Almo, S.C. Structure and T-Cell Inhibition Properties of B7 Family Member, B7-H3. Structure 2013, 21, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.; Wang, Y.; Wu, X.; Han, J.; Li, G.; Hua, M.; Han, K.; Zhang, H.; Li, A.; Zeng, H. Storm of Soluble Immune Checkpoints Associated with Disease Severity of COVID-19. Signal Transduct. Target. Ther. 2020, 5, 192. [Google Scholar] [CrossRef] [PubMed]
- Martín-Quirós, A.; Maroun-Eid, C.; Avendaño-Ortiz, J.; Lozano-Rodríguez, R.; Valentín Quiroga, J.; Terrón, V.; Montalbán-Hernández, K.; García-Garrido, M.A.; Muñoz del Val, E.; del Balzo-Castillo, Á.; et al. Potential Role of the Galectin-9/TIM-3 Axis in the Disparate Progression of SARS-CoV-2 in a Married Couple: A Case Report. BMH 2021, 6, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Galan, L.; Ruiz, A.; Martinez-Espinosa, K.; Aguilar-Duran, H.; Torres, M.; Falfan-Valencia, R.; Pérez-Rubio, G.; Selman, M.; Buendia-Roldan, I. Circulating Levels of PD-L1, TIM-3 and MMP-7 Are Promising Biomarkers to Differentiate COVID-19 Patients That Require Invasive Mechanical Ventilation. Biomolecules 2022, 12, 445. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Zhao, C.; Ozarina, M.; Zhao, X.; Yang, J.; Chen, H. Targeting Immune Checkpoints in Lung Cancer: Current Landscape and Future Prospects. Clin. Drug Investig. 2019, 39, 341–353. [Google Scholar] [CrossRef]
- Shi, T.; Ma, Y.; Yu, L.; Jiang, J.; Shen, S.; Hou, Y.; Wang, T. Cancer Immunotherapy: A Focus on the Regulation of Immune Checkpoints. Int. J. Mol. Sci. 2018, 19, 1389. [Google Scholar] [CrossRef] [Green Version]
- Park, L.M.; Lannigan, J.; Jaimes, M.C. OMIP-069: Forty-Color Full Spectrum Flow Cytometry Panel for Deep Immunophenotyping of Major Cell Subsets in Human Peripheral Blood. Cytom. Part A 2020, 97, 1044–1051. [Google Scholar] [CrossRef]
- Niewold, P.; Ashhurst, T.M.; Smith, A.L.; King, N.J.C. Evaluating Spectral Cytometry for Immune Profiling in Viral Disease. Cytom. Part A 2020, 97, 1165–1179. [Google Scholar] [CrossRef]
- Iba, T.; Connors, J.M.; Levy, J.H. The Coagulopathy, Endotheliopathy, and Vasculitis of COVID-19. Inflamm. Res. 2020, 69, 1181–1189. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, M.; Zhang, S.; Xia, P.; Cao, W.; Jiang, W.; Chen, H.; Ding, X.; Zhao, H.; Zhang, H.; et al. Coagulopathy and Antiphospholipid Antibodies in Patients with COVID-19. N. Engl. J. Med. 2020, 382, e38. [Google Scholar] [CrossRef]
- Malas, M.B.; Naazie, I.N.; Elsayed, N.; Mathlouthi, A.; Marmor, R.; Clary, B. Thromboembolism Risk of COVID-19 Is High and Associated with a Higher Risk of Mortality: A Systematic Review and Meta-Analysis. EClinicalMedicine 2020, 29, 100639. [Google Scholar] [CrossRef]
- Bhatt, P.J.; Shiau, S.; Brunetti, L.; Xie, Y.; Solanki, K.; Khalid, S.; Mohayya, S.; Au, P.H.; Pham, C.; Uprety, P.; et al. Risk Factors and Outcomes of Hospitalized Patients with Severe COVID-19 and Secondary Bloodstream Infections: A Multicenter, Case-Control Study. Clin. Infect. Dis. 2020, 72, e995–e1003. [Google Scholar] [CrossRef] [PubMed]
- Carlet, J.; Payen, D.; Opal, S.M. Steroids for Sepsis and ARDS: This Eternal Controversy Remains with COVID-19. Lancet 2020, 396, e61–e62. [Google Scholar] [CrossRef]
- Boumaza, A.; Gay, L.; Mezouar, S.; Bestion, E.; Diallo, A.B.; Michel, M.; Desnues, B.; Raoult, D.; La Scola, B.; Halfon, P.; et al. Monocytes and Macrophages, Targets of SARS-CoV-2: The Clue for COVID-19 Immunoparalysis. J. Infect. Dis. 2021, 224, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Torres-Flores, J.M.; Silva-Ayala, D.; Espinoza, M.A.; López, S.; Arias, C.F. The Tight Junction Protein JAM-A Functions as Coreceptor for Rotavirus Entry into MA104 Cells. Virology 2015, 475, 172–178. [Google Scholar] [CrossRef] [Green Version]
- Mandell, K.J.; Babbin, B.A.; Nusrat, A.; Parkos, C.A. Junctional Adhesion Molecule 1 Regulates Epithelial Cell Morphology through Effects on Β1 Integrins and Rap1 Activity. J. Biol. Chem. 2005, 280, 11665–11674. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Carrette, F.; Barraza, M.L.; Otero, D.C.; Magaña, J.; Bosenberg, M.W.; Swain, S.L.; Bradley, L.M. PSGL-1 Is an Immune Checkpoint Regulator That Promotes T Cell Exhaustion. Immunity 2016, 44, 1190–1203. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Otero, D.C.; Takahashi, A.A.; Bradley, L.M. PSGL-1: A New Player in the Immune Checkpoint Landscape. Trends Immunol. 2017, 38, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, R.; Neubert, E.N.; Stairiker, C.J.; Henriquez, M.L.; Bradley, L.M. PSGL-1 Is a T Cell Intrinsic Inhibitor That Regulates Effector and Memory Differentiation and Responses During Viral Infection. Front. Immunol. 2021, 12, 677824. [Google Scholar] [CrossRef]
- André, S.; Picard, M.; Cezar, R.; Roux-Dalvai, F.; Alleaume-Butaux, A.; Soundaramourty, C.; Cruz, A.S.; Mendes-Frias, A.; Gotti, C.; Leclercq, M.; et al. T Cell Apoptosis Characterizes Severe COVID-19 Disease. Cell. Death Differ. 2022. ahead of print. [Google Scholar] [CrossRef]
- Bellesi, S.; Metafuni, E.; Hohaus, S.; Maiolo, E.; Marchionni, F.; D’Innocenzo, S.; La Sorda, M.; Ferraironi, M.; Ramundo, F.; Fantoni, M.; et al. Increased CD95 (Fas) and PD-1 Expression in Peripheral Blood T Lymphocytes in COVID-19 Patients. Br. J. Haematol. 2020, 191, 207–211. [Google Scholar] [CrossRef]
- Newton, K.; Dixit, V.M.; Kayagaki, N. Dying Cells Fan the Flames of Inflammation. Science 2021, 374, 1076–1080. [Google Scholar] [CrossRef] [PubMed]
- Pascutti, M.F.; Geerman, S.; Slot, E.; van Gisbergen, K.P.J.M.; Boon, L.; Arens, R.; van Lier, R.A.W.; Wolkers, M.C.; Nolte, M.A. Enhanced CD8 T Cell Responses through GITR-Mediated Costimulation Resolve Chronic Viral Infection. PLoS Pathog. 2015, 11, e1004675. [Google Scholar] [CrossRef] [PubMed]
- Buechele, C.; Baessler, T.; Wirths, S.; Schmohl, J.U.; Schmiedel, B.J.; Salih, H.R. Glucocorticoid-Induced TNFR-Related Protein (GITR) Ligand Modulates Cytokine Release and NK Cell Reactivity in Chronic Lymphocytic Leukemia (CLL). Leukemia 2012, 26, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Okuno, T.; Yamamoto, M.; Pasterkamp, R.J.; Takegahara, N.; Takamatsu, H.; Kitao, T.; Takagi, J.; Rennert, P.D.; Kolodkin, A.L.; et al. Semaphorin 7A Initiates T-Cell-Mediated Inflammatory Responses through A1β1 Integrin. Nature 2007, 446, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Yeo, J.; Ko, M.; Lee, D.-H.; Park, Y.; Jin, H.-S. TIGIT/CD226 Axis Regulates Anti-Tumor Immunity. Pharmaceuticals 2021, 14, 200. [Google Scholar] [CrossRef]
- Huang, Z.; Qi, G.; Miller, J.S.; Zheng, S.G. CD226: An Emerging Role in Immunologic Diseases. Front. Cell. Dev. Biol. 2020, 8, 564. [Google Scholar] [CrossRef]
- Braun, M.; Aguilera, A.R.; Sundarrajan, A.; Corvino, D.; Stannard, K.; Krumeich, S.; Das, I.; Lima, L.G.; Meza Guzman, L.G.; Li, K.; et al. CD155 on Tumor Cells Drives Resistance to Immunotherapy by Inducing the Degradation of the Activating Receptor CD226 in CD8+ T Cells. Immunity 2020, 53, 805–823.e15. [Google Scholar] [CrossRef]
- Weulersse, M.; Asrir, A.; Pichler, A.C.; Lemaitre, L.; Braun, M.; Carrié, N.; Joubert, M.-V.; Moine, M.L.; Souto, L.D.; Gaud, G.; et al. Eomes-Dependent Loss of the Co-Activating Receptor CD226 Restrains CD8+ T Cell Anti-Tumor Functions and Limits the Efficacy of Cancer Immunotherapy. Immunity 2020, 53, 824–839.e10. [Google Scholar] [CrossRef]
- Duan, S.; Paulson, J.C. Siglecs as Immune Cell Checkpoints in Disease. Annu. Rev. Immunol. 2020, 38, 365–395. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.; Sari-Ak, D.; Bagga, T. Siglecs as Therapeutic Targets in Cancer. Biology 2021, 10, 1178. [Google Scholar] [CrossRef]
- Fehervari, Z. Targeting Cancer by Siglecs. Nat. Immunol. 2018, 19, 1148. [Google Scholar] [CrossRef] [PubMed]
- van Houtum, E.J.H.; Büll, C.; Cornelissen, L.A.M.; Adema, G.J. Siglec Signaling in the Tumor Microenvironment. Front. Immunol. 2021, 12, 790317. [Google Scholar] [CrossRef] [PubMed]
- Vuchkovska, A.; Iwashima, M. Siglec 5-a Novel Checkpoint Receptor in T Cells. J. Immunol. 2020, 204, 238–248. [Google Scholar]
- Lozano-Rodriguez, R.; Avendano-Ortiz, J.; Montalban-Hernandez, K.; Ruiz-Rodriguez, J.C.; Ferrer, R.; Martin-Quiros, A.; Maroun-Eid, C.; Gonzalez-Lopez, J.J.; Fabrega, A.; Terron, V.; et al. SIGLEC5: An Immune Checkpoint Ligand in Sepsis. medRxiv 2020. [Google Scholar] [CrossRef]
- Montalbán-Hernández, K.; Cantero-Cid, R.; Lozano-Rodríguez, R.; Pascual-Iglesias, A.; Avendaño-Ortiz, J.; Casalvilla-Dueñas, J.C.; Bonel Pérez, G.C.; Guevara, J.; Marcano, C.; Barragán, C.; et al. Soluble SIGLEC5: A New Prognosis Marker in Colorectal Cancer Patients. Cancers 2021, 13, 3896. [Google Scholar] [CrossRef]
- Tsai, T.-Y.; Huang, M.-T.; Sung, P.-S.; Peng, C.-Y.; Tao, M.-H.; Yang, H.-I.; Chang, W.-C.; Yang, A.-S.; Yu, C.-M.; Lin, Y.-P.; et al. SIGLEC-3 (CD33) Serves as an Immune Checkpoint Receptor for HBV Infection. J. Clin. Investig. 2021, 131, 141965. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, P.S.; Rezaei, N. Immune Checkpoint Inhibition in COVID-19: Risks and Benefits. Expert Opin. Biol. Ther. 2021, 21, 1173–1179. [Google Scholar] [CrossRef]


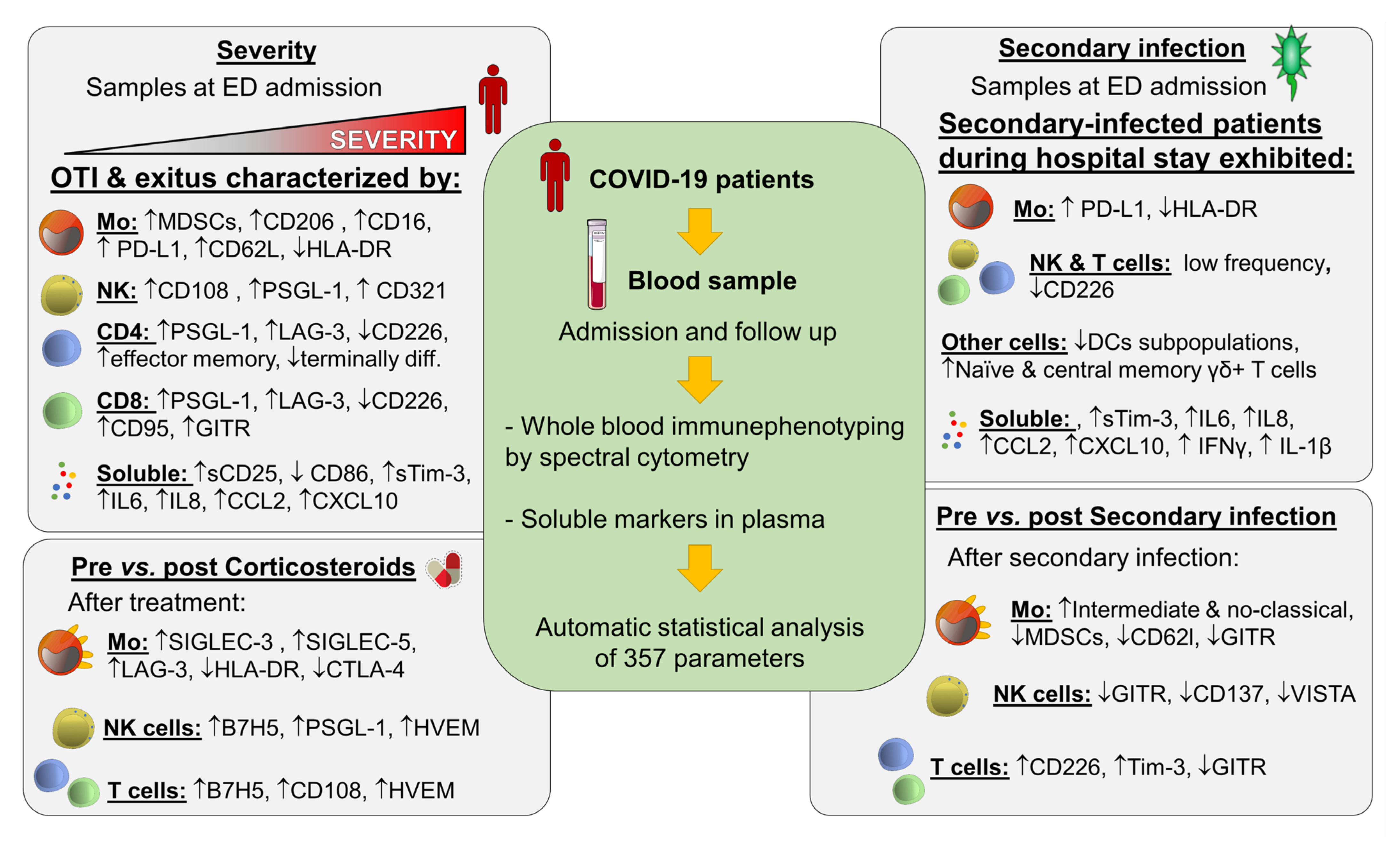{kind=link}
| Population | No Hospitalized (n = 22) | Hospitalized with No O2 (n = 10) | O2 Requirement (n = 23) | OrotrachealIntubation (n = 13) | Exitus (n = 19) | p-Value | |
|---|---|---|---|---|---|---|---|
| Monocytes (MΦ) | m-MDSCs (% of total leukocytes) | 0.95 (0.53–1.37) | 0.68 (0.6–1.36) | 0.74 (0.49–1.73) | 2.61 (0.93–3.74) | 1.55 (0.91–3.63) | 0.04 * |
| CD16 | 47,765 (42,068–51,544) | 50,548 (47,680–51,714) | 47,259 (41,927–56,258) | 54,050 (48,199–72,597) | 58,286 (50,114–68,235) | 0.04 * | |
| CD137 | 12,375 (10,704–14,070) | 15,033 (14,701–17,356) | 14,905 (12,882–16,091) | 15,963 (13,351–21,575) | 14,529 (10,466–16,733) | 0.03 * | |
| CD206 | 8009 (7246–8668) | 8722 (7906–9937) | 8492 (7200–9525) | 9503 (8633–12,995) | 9191 (7480–10,997) | 0.03 * | |
| CD223 | −2832 (−3599–(−2303)) | −998 (−1743–2.1) | −1596 (−2967–(−446)) | −1290 (−2342–1471) | −2497 (−3521–(−661)) | 0.02 * | |
| GITR | 11,878 (8035–15,868) | 17,396 (16,751–18,156) | 16,795 (11,010–19,404) | 19,646 (15,132–27,655) | 16,033 (8000–19,872) | 0.01 * | |
| HLA-DR | 89,341 (67,377–119,767) | 71,207 (58,657–111,503) | 98,626 (53,345–154,812) | 13,075 (116–34,984) | 22,586 (3363–44,757) | <0.01 * | |
| HLA-DR (on classical MΦ) | 41,211 (31,131–54,789) | 32,889 (28,380–61,435) | 44,596 (26,126–65,190) | 17,781 (13,158–22,383) | 25,505 (14,868–31,224) | <0.01 * | |
| PD-L1 (on classical MΦ) | 19,373 (17,394–21,817) | 19,117 (16,622–28,767) | 22,188 (18,085–31,198) | 27,525 (22,930–43,874) | 26,049 (23,386–38,107) | 0.01 * | |
| PD-L1 (on intermediate MΦ) | 13,945 (9906–15,559) | 12,795 (9162–16,247) | 15,600 (13,303–19,005) | 17,768 (14,871–20,041) | 19,883 (15,799–24,578) | 0.04 * | |
| CD62L (on classical MΦ) | 3407 (2024–4631) | 4498 (3424–5702) | 4554 (2798–6036) | 3830 (2057–4506) | 5607 (4083–8203) | 0.048 * | |
| CD62L (on intermediate MΦ) | 2980 (1533–4121) | 3931 (2519–5148) | 3901 (3036–5636) | 2563 (1444–3039) | 4772 (3728–7378) | <0.01 * | |
| CD62L (on non-classic MΦ) | 1402 (−383–3843) | 6089 (4960–9107) | 4895 (1090–6778) | 486 (−118–3492) | 5295 (2142–6421) | <0.01 * | |
| Neutrophils | CD206 | 4754 (3979–4982) | 6591 (5695–7236) | 5698 (4311–7042) | 5441 (4881–8097) | 6266 (5119–8150) | 0.02 * |
| CD226 | 9314 (7735–10,897) | 8814 (5757–10,431) | 8893 (7126–10,495) | 10,498 (8864–13,827) | 11,449 (9576–11,957) | 0.04 * | |
| Tim-3 | −268 (−454–(−123)) | 96 (−248–765) | −56 (−160–280) | −157 (−297–72) | −76 (387–54) | 0.03 * | |
| Dendritic Cells (% of total leukocytes) | Myeloid (CD11c+CD123+) | 0.23 (0.13–0.34) | 0.18 (0.07–0.28) | 0.15 (0.09–0.32) | 0.047 (0.018–0.11) | 0.09 (0.01–0.16) | 0.01 * |
| CD141+ | 0.074 (0.017–0.185) | 0.031 (0.01–0.08) | 0.05 (0.025–0.07) | 0.013 (0.003–0.017) | 0.02 (0.007–0.05) | 0.02 * | |
| NK cells | CD56bright (% of total leukocytes) | 0.15 (0.09–0.27) | 0.11 (0.01–0.14) | 0.09 (0.068–0.12) | 0.04 (0.02–0.09) | 0.05 (0.02–0.09) | 0.01 * |
| CD108 | 1001 (757–1122) | 1409 (1211–1761) | 1450 (993–1637) | 1509 (1123–2554) | 1343 (966–2045) | 0.02 * | |
| CD137 | 5732 (4970–6693) | 7283 (7102–8826) | 7138 (6064–7936) | 7487 (6121–10,070) | 7043 (5112–7680) | 0.03 * | |
| CD162 (PSGL-1) | 5621 (4849–7189) | 6191 (2458–10,102) | 5915 (4413–8319) | 13,321 (9722–16,325) | 7689 (5892–11,417) | <0.01 * | |
| CD270 (HVEM) | 37,940 (32,249–41,290) | 41,220 (40,686–42,794) | 41,617 (40,485–44,314) | 40,851 (36,233–47,783) | 41,951 (37,003–48,630) | 0.03 * | |
| CD321 | 13,624 (12,363–14,122) | 14,573 (13,506–19,158) | 15,719 (14,455–18,211) | 13,842 (12,854–22,062) | 16,248 (14,482–21,413) | 0.03 * | |
| GITR | 6324 (3639–7357) | 8786 (8697–9578) | 8766 (4694–9623) | 8949 (6872–13,358) | 8600 (4009–11,205) | 0.01 * | |
| GITR-L | 7141 (5566–9358) | 10,977 (10,013–11,386) | 10,868 (8042–11,318) | 9536 (7434–11,751) | 8277 (6766–10,993) | 0.02 * | |
| TIGIT | −1980 (−3328–(−466)) | 1233 (960–1897) | 251 (−2306–1453) | −703 (−1798–718) | −38 (−3501–2315) | 0.04 * | |
| CD3+ lymphocytes | CD3+ (% on total leukocytes) | 17.1 (14–21.8) | 9.2 (3.1–21.5) | 12.7 (5.2–17.2) | 5.7 (2.3–8.9) | 6.1 (2.7–13.1) | 0.01 * |
| CD108 | 1036 (791–1109) | 1337 (1106–1523) | 1335 (871–1514) | 1439 (1015–2060) | 1136 (925–1404) | 0.03 * | |
| CD137 | 5380 (4631–5938) | 6480 (6183–7581) | 6397 (4622–6827) | 6211 (5608–8997) | 5970 (4605–6656) | 0.048 * | |
| CD152 (%) | 1.35 (0.95–3.3) | 6.00 (2.9–6.6) | 3.44 (1.46–4.84) | 2.43 (0.75–3.95) | 4.44 (1.77–7.19) | 0.04 * | |
| CD162 (PSGL-1) | 5404 (4793–6388) | 6445 (6022–8215) | 6027 (4362–7906) | 10,556 (8427–12,860) | 6719 (5076–11,404) | <0.01 * | |
| CD223 | −1041 (−1319–(−835)) | −210 (−427–259) | −262 (−819–96) | −202 (−855–638) | −581 (−1108–218) | 0.01 * | |
| CD223 (%) | 1.23 (0.48–2.37) | 5.40 (1.36–8.8) | 4.19 (2.3–7.82) | 14.44 (6.77–37.4) | 6.41 (1.64–20.8) | <0.01 * | |
| CD226 (%) | 62.20 (48–78) | 41.20 (38.5–68.6) | 49.75 (25.1–67.6) | 54.7 (47.6–66.7) | 12.0 (6–64) | 0.02 * | |
| GITR | 5756 (3682–6965) | 7899 (7509–8308) | 7752 (4192–8869) | 7833 (6700–12,074) | 7687 (3707–9623) | 0.04 * | |
| TIGIT | −1805 (−3119–(−688)) | 340 (69–927) | 24 (−2422–876) | −585 (−1538–51) | −1307 (−3652–952) | 0.04 * | |
| Tim-3 | −36 (−157–52) | 93 (59–154) | 77 (−11–155) | 58 (−96–94) | 53 (−32–133) | 0.03 * | |
| CD4+ T cells | CD4+ (% on CD3+) | 41 (31–52) | 40 (32–46) | 40 (27–50) | 27 (24–2−34) | 29 (18–38) | 0.03 * |
| CD4em | 12.5 (6.8–52.2) | 11.7 (6.9–26.9) | 26 (13–34.5) | 8.8 (6.5–13.7) | 29.3 (20–53) | 0.04 * | |
| CD4td | 3.3 (1–14.8) | 3.6 (1.7–9.2) | 4.6 (1.5–17.8) | 7.5 (5.2–14.9) | 18.3 (9.4–22.4) | 0.01 * | |
| CD95 | 28,072 (25,207–32,591) | 38,858 (25,782–52,147) | 39,334 (26,476–48,056) | 25,126 (23,151–30,421) | 34,386 (32,863–43,106) | 0.04 * | |
| CD95 on CD4n | 10,044 (7857–12,140) | 14,434 (11,777–19,201) | 13,791 (8851–19,808) | 8366 (7302–10,149) | 13,444 (11,585–19,296) | 0.02 * | |
| CD108 | 1044 (777–1091) | 1341 (1138–1554) | 1359 (882–1539) | 1343 (1028–2195) | 1171 (915–1530) | 0.03 * | |
| CD108 (%) | 64.70 (56–69) | 53.80 (47.4–62.7) | 56.5 (51–64.2) | 65.3 (57.7–72.2) | 68.85 (53.7–76) | 0.04 * | |
| CD162 (PSGL-1) | 5444 (4809–6069) | 6299 (5553–7911) | 6371 (4414–8705) | 9790 (8489–13,163) | 6975 (5501–9656) | <0.01 * | |
| CD223 | −1026 (−1299–(−814)) | −122 (−330–391) | −208 (−853–131) | −233 (−813–828) | −563 (−1019–267) | 0.01 * | |
| CD223 (%) | 1.94 (1.32–5.1) | 10.50 (4.37–17.9) | 7.45 (2.67–16.5) | 16.4 (9.14–47.1) | 14.8 (4.07–34.6) | <0.01 * | |
| CD226 (%) | 66.20 (44.5–78.7) | 43.30 (38.9–69) | 49.5 (28.47–67.5) | 56.6 (44.17–63) | 26.3 (19.32–54.3) | 0.048 * | |
| GITR-L | 6152 (4177–7700) | 8802 (8358–9421) | 8696 (6227–9672) | 7276 (5723–9199) | 7040 (5452–8249) | 0.047 * | |
| TIGIT | −1869 (−3057–(−781)) | 432 (216–1224) | 176 (−2448–992) | −547 (−1451–312) | −1335 (−3623–975) | 0.02 * | |
| Tim-3 | −33 (−150–63) | 133 (92–181) | 105 (−2–170) | 62 (−90–101) | 51 (−33–156) | 0.01 * | |
| CD8+ T cells | CD95 | 28,294 (24,290–32,358) | 34,163 (30,558–42,046) | 35,421 (28,778–44,111) | 28,917 (25,500–31,972) | 38,506 (34,680–47,282) | 0.01 * |
| CD95 on CD8em | 62,501 (59,155–65,520) | 64,776 (64,905–91,663) | 67,625 (61,148–78,483) | 66,515 (61,442–72,600) | 73,030 (67,485–76,648) | 0.03 * | |
| CD95 on CD8n | 14,040 (11,774–17,343) | 15,616 (13,952–16,373) | 19,502 (15,790–27,431) | 14,319 (10,584–16,957) | 22,946 (17,821–30,006) | 0.01 * | |
| CD95 on CD8td | 43,095 (40,524–45,710) | 44,195 (40,622–46,163) | 43,763 (42,306–51,144) | 44,161 (41,126–49,327) | 48,633 (46,499–51,618) | 0.04 * | |
| CD137 | 5572 (4914–5907) | 6179 (5981–7071) | 6103 (4956–6422) | 6653 (5460–9098) | 6077 (5001–6970) | 0.04 * | |
| CD162 (PSGL-1) | 6126 (4876–7704) | 8503 (6556–9800) | 6690 (4427–8301) | 11,804 (9267–14,075) | 7002 (4975–13,726) | <0.01 * | |
| CD8+ T cells | CD223 | −1201 (−1316–(−942)) | −373 (−643–83) | −310 (−988–(−47)) | −304 (−954–424) | −711 (−1226–107) | 0.01 * |
| CD223 (%) | 5.63 (2.88–7.8) | 10.8 (3.8–24.8) | 8.16 (4.41–22.3) | 22.35 (13.2–41.4) | 14.5 (6.6–34.2) | 0.01 * | |
| CD226 (%) | 65.1 (48.75–69.1) | 55.40 (43.6–77.4) | 68.35 (52.45–77.3) | 57.8 (44.3–68.6) | 33.65 (10–58.1) | 0.03 * | |
| GITR | 5831 (3668–6673) | 7434 (6981–7621) | 7200 (4322–8030) | 8038 (6458–12,341) | 7256 (3877–9892) | 0.01 * | |
| Tim-3 | −43 (−174–33) | 89 (77–160) | 99 (20–150) | 40 (−112–70) | 49 (−44–135) | 0.01 * | |
| γδ+ T cells | γδ+ (on total leukocytes) | 0.35 (0.18–0.75) | 0.08 (0.06–0.12) | 0.22 (0.06–0.51) | 0.14 (0.05–0.32) | 0.09 (0.06–0.21) | 0.04 * |
| B cells | CD19+ (% on total leukocytes) | 2.61 (1.37–4.38) | 1.52 (0.92–3.21) | 1.90 (0.88–2.5) | 0.72 (0.48–1.52) | 0.69 (0.15–1.71) | <0.01 * |
| trB cell (% on total leukocytes) | 1.04 (0.27–1.39) | 0.28 (0.18–0.38) | 0.40 (0.16–0.68) | 0.34 (0.15–0.69) | 0.20 (0.05–0.72) | 0.02 * | |
| Plasma cell (% on CD19+) | 9.15 (2.62–14.4) | 2.31 (1.9–4.16) | 4.02 (1.22–10.5) | 21.5 (7–24.85) | 10.1 (3.12–16.5) | 0.04 * |
| Type | No Secondary Infection (n = 54) | Secondary Infection (n = 33) | p-Value | |
|---|---|---|---|---|
| Monocytes | HLA-DR | 74,440 (45,851–111,503) | 38,090 (13,075–89,843) | 0.048 * |
| PD-L1 | 14,218 (11,665–18,379) | 17,496 (15,090–21,905) | 0.03 * | |
| Neutrophils | CD206 | 4875 (4104–6600) | 6244 (5094–8127) | 0.03 * |
| CD226 | 8805 (7398–10,773) | 10,708 (9325–11,952) | 0.01 * | |
| Dendritic cells (% of total leukocytes) | Myeloid (CD11c+CD123+) | 0.175 (0.092–0.315) | 0.073 (0.018–0.165) | <0.01 * |
| Plasmacytoid (CD11c-CD123+) | 0.044 (0.02–0.114) | 0.011 (0.005–0.064) | 0.01 * | |
| CD1c+ | 0.180 (0.076–0.418) | 0.077 (0.024–0.18) | <0.01 * | |
| CD141+ | 0.048 (0.017–0.084) | 0.010 (0.003–0.031) | <0.01 * | |
| NK cells | CD3-CD56+ (% on total leukocytes) | 2.905 (1.39–4.67) | 0.770 (0.475–1.895) | <0.01 * |
| CD56dim (% of total NKs) | 2.670 (1.29–4.51) | 0.720 (0.460–1.82) | <0.01 * | |
| CD56bright (% of total NKs) | 0.105 (0.07–0.178) | 0.045 (0.014–0.098) | <0.01 * | |
| CD226 | 10,064 (8033–12,322) | 8733 (6803–10,029) | 0.01 * | |
| NKT cells | CD56+CD3+ (% on total leukocytes) | 1.0 (0.5–2.3) | 0.5 (0.2–1.0) | 0.03 * |
| CD3+ lymphocytes | CD3+ (% on total leukocytes) | 14.6 (6.4–19.8) | 6.1 (2.8–10.3) | <0.01 * |
| CD223 (%) | 2.65 (0.84–8.01) | 7.27 (2.38–20.2) | 0.01 * | |
| CD226 | 9491 (6584–12,228) | 7216 (5546–8445) | <0.01 * | |
| CD226 (%) | 57.1 (41.4–72.7) | 35.6 (11.05–67.1) | 0.02 * | |
| CD4+ T cells | CD4+ (% on total leukocytes) | 38.3 (30.6–50.8) | 27.4 (20.8–39.5) | <0.01 * |
| CD4+ (% on CD3+) | 63.3 (55.9–69.3) | 49.2 (25.5–66.3) | 0.01 * | |
| CD223 (%) | 5.67 (1.94–18.1) | 12.6 (5.75–20.65) | 0.03 * | |
| CD226 | 9347 (6878–12,193) | 7306 (5933–8640) | <0.01 * | |
| CD226 (%) | 58.3 (38.9–77.2) | 38.7 (18.05–60.55) | <0.01 * | |
| CD276 (%) | 14.7 (6.2–26.3) | 5.08 (1.43–22.8) | 0.02 * | |
| CD8+ T cells | CD223 (%) | 6.84 (3.8–17.9) | 15.40 (7.18–27.7) | 0.03 * |
| CD226 | 10,016 (7291–12,953) | 7824 (6081–9337) | 0.03 * | |
| CD226 (%) | 62.9 (46.3–73.7) | 43.6 (18.65–64.05) | 0.01 * | |
| CD276 (%) | 12.6 (7.08–22.6) | 5.0 (2.57–17.65) | 0.02 * | |
| CD95 on CD8em | 64,506 (59,473–73,422) | 72,949 (66,760–76,753) | 0.01 * | |
| γδ+ T cells | Naïve (%) | 8.2 (3.1–28.2) | 30.5 (9.9–53.6) | 0.01 * |
| Central memory (%) | 16.6 (10.1–38.9) | 33.1 (21.4–44.8) | 0.01 * | |
| B cells | CD19+ (% on total leukocytes) | 1.9 (1.1–3.7) | 0.8 (0.3–1.6) | <0.01 * |
| nB cell (% on CD19+) | 38.6 (22.2–51.2) | 20.5 (3.3–38.8) | 0.02 * | |
| smB cell (% on CD19+) | 13.7 (6.6–32) | 26.0 (14–48.8) | 0.03 * | |
| Plasma cell (% on CD19+) | 3.2 (0.7–6.8) | 6.6 (1.1–12.6) | 0.04 * |
| Type | Pre-Secondary Infection | Post-Secondary Infection | p-Value | |
|---|---|---|---|---|
| Chemokines and Cytokines | CXCL10 | 154.9 (98.5–409) | 124.2 (66–182) | 0.01 * |
| IFNγ | 30.4 (5.9–44.4) | 1.4 (0–38.8) | 0.01 * | |
| Il-2 | 3.7 (0–8) | 0.7 (0–5) | 0.046 * | |
| IL-6 | 78.1 (26.4–230) | 44 (5.2–91) | 0.01 * | |
| IL-10 | 8 (0–14.5) | 1.8 (0–10.2) | 0.03 * | |
| TNFα | 0 (0–5.3) | 0 (0–0) | <0.01 * | |
| Immune checkpoint | sPD-1 | 4.6 (3.4–14.1) | 3.6 (2–7.2) | <0.01 * |
| Galectin-9 | 110,810 (39,726–162,758) | 44,249 (33,744–68,212) | <0.01 * | |
| Thrombosis | sP-Selectin | 14,173 (6786–39,637) | 8819 (1326–24,858) | 0.03 * |
| sPSGL-1 | 206.4 (0–11,689) | 0 (0–266) | 0.01 * | |
| Monocytes | CD14+ (% of total leukocytes) | 5.0 (1.8–8.6) | 6.8 (4.4–9.1) | 0.04 * |
| m-MDSCs (% of total leukocytes) | 1.18 (0.71–2.3) | 0.87 (0.52–1.6) | 0.02 * | |
| CD223 | −575.9 (−1338–1487) | −1568 (−2472–1186) | 0.04 * | |
| GITR | 18,059 (9591–23,306) | 10,086 (6968–21,470) | 0.03 * | |
| Intermediate MΦ (% of total leukocytes) | 0.2 (0.1–0.3) | 0.3 (0.2–0.5) | 0.01 * | |
| Non-classic MΦ (% of total leukocytes) | 0.2 (0.1–0.5) | 0.7 (0.4–1) | <0.01 * | |
| CD62L (on non-classic MΦ) | 3759 (270–7275) | 912 (−6.6–2196) | 0.01 * | |
| NK cells | B7H5 | 2715 (2120–3783) | 2020 (1620–3566) | 0.04 * |
| CD108 | 1661 (1179–2107) | 863 (494–1995) | <0.01 * | |
| CD137 | 7491 (4999–9334) | 5274 (4089–8188) | 0.02 * | |
| GITR | 8974 (4634–13,211) | 4495 (3643–10,609) | <0.01 * | |
| GITR-L | 10,434 (8021–11,657) | 7648 (6541–11,361) | 0.01 * | |
| CD4+ T cells | CD4cm | 34 (13.2–43.3) | 45 (30–57.7) | <0.01 * |
| CD4td | 15.8 (6.1–30.2) | 3.2 (1.7–6.8) | <0.01 * | |
| B7H5 | 2284 (1941–3077) | 1721 (1396–2693) | 0.047 * | |
| CD108 | 1459 (1030–1659) | 896 (653–1857) | 0.01 * | |
| CD223 | 99.8 (−565–1429) | −342 (−895–1128) | 0.03 * | |
| CD226 (%) | 46.7 (31–61.6) | 63.2 (48–78) | <0.01 * | |
| CD226 | 7943 (6665–10,849) | 10,158 (7808–13,161) | 0.02 * | |
| GITR | 8249 (4171–13,928) | 4357 (3587–9626) | 0.02 * | |
| Tim-3 (%) | 43.2 (37.4–50.6) | 50.85 (39–61.3) | 0.02 * | |
| CD8+ T cells | CD108 | 1252 (924–1776) | 956 (711–1604) | 0.02 * |
| CD137 | 6311 (4754–8189) | 4843 (3938–7123) | 0.03 * | |
| CD152 (%) | 21.2 (9.2–33.4) | 36.2 (16.6–48.8) | 0.02 * | |
| GITR | 7572 (4274–11,453) | 4130 (3229.8–8635) | <0.01 * | |
| γδ+ T cells | γδ+ (% on total leukocytes) | 0.135 (0.04–0.365) | 0.22 (0.11–0.49) | 0.04 * |
| Central memory (%) | 28.55 (14.3–46.1) | 42 (29–51.6) | 0.01 * | |
| B cells | nB cell (% on CD19+) | 28.15 (17.6–51) | 18.8 (6.5–33) | 0.02 * |
| trB cell (% on CD19+) | 7.78 (0.86–12.7) | 12.8 (4.2–20.5) | 0.02 * | |
| CD27−IgD− (% on CD19+) | 24.25 (15–39.8) | 35.8 (18.6–53) | 0.04 * |
| Type | Pre-Treatment | Post-Treatment | p-Value | |
| Chemokines | CXCL10 | 232 (106–693) | 138 (73–234) | 0.03 |
| Monocytes | CD33 (Siglec-3) | 15,857 (7479–18,207) | 19,236 (14,302–24,540) | 0.02 |
| CD152 (CTLA-4) | 639 (314–1259) | −452 (−2574–712) | <0.01 | |
| CD170 (Siglec-5) | −2215 (−3086–(−714)) | 446 (−1662–7808) | 0.01 | |
| CD223 | −1495 (−3479–(−746)) | −576 (−2338–2259) | 0.01 | |
| CD276 (B7H3) | 3225 (2268–4559) | 625 (−2643–4110) | 0.02 | |
| HLA-DR | 103,485 (64,040–111,089) | 55,363 (24,152–93,803) | 0.047 | |
| HLA-DR (on classical MΦ) | 48,135 (33,733–68,059) | 31,843 (17,650–47,653) | 0.02 | |
| PD-L1 (on non-classic MΦ) | 16,881 (4664–27,670) | 33,213 (6880–40,379) | 0.04 | |
| Dendritic Cells | CD1c+ (% of total DC) | 17.5 (11.4–23.1) | 6.8 (2.53–14.53) | 0.02 |
| NK cells | B7H5 | 2223 (1780–2530) | 2769 (1983–4710) | 0.01 |
| CD162 (PSGL-1) | 6178 (4079–9165) | 8716 (5885–11,483) | 0.04 | |
| CD270 (HVEM) | 40,659 (33,705–42,478) | 42,504 (39,611–56,296) | 0.02 | |
| CD4+ T cells | B7H5 | 1857 (1629–2030) | 2273 (1746–4452) | 0.01 |
| CD108 | 1085 (789–1289) | 1431 (866–2706) | 0.03 | |
| CD226 | 7179 (6213–9084) | 9551 (7460–12,402) | 0.03 | |
| CD270 (HVEM) | 33,929 (29,624–34,958) | 37,145 (34,469–49,570) | <0.01 | |
| CD8+ T cells | B7H5 | 1932 (1746–2181) | 2397 (1924–3829) | 0.01 |
| CD108 | 970 (848–1109) | 1250 (915–2286) | 0.02 | |
| CD223 (%) | 5.11 (3–18.1) | 17 (5.06–69.3) | 0.04 | |
| CD270 (HVEM) | 35,512 (33,183–36,840) | 38,963 (35,854–47,186) | <0.01 | |
| B cells | usB cell (% on CD19+) | 4.16 (2.35–6.35) | 7.7 (3.76–13.7) | 0.047 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lozano-Rodríguez, R.; Terrón-Arcos, V.; López, R.; Martín-Gutiérrez, J.; Martín-Quirós, A.; Maroun-Eid, C.; del Val, E.M.; Cañada-Illana, C.; Pascual Iglesias, A.; Quiroga, J.V.; et al. Differential Immune Checkpoint and Ig-like V-Type Receptor Profiles in COVID-19: Associations with Severity and Treatment. J. Clin. Med. 2022, 11, 3287. https://doi.org/10.3390/jcm11123287
Lozano-Rodríguez R, Terrón-Arcos V, López R, Martín-Gutiérrez J, Martín-Quirós A, Maroun-Eid C, del Val EM, Cañada-Illana C, Pascual Iglesias A, Quiroga JV, et al. Differential Immune Checkpoint and Ig-like V-Type Receptor Profiles in COVID-19: Associations with Severity and Treatment. Journal of Clinical Medicine. 2022; 11(12):3287. https://doi.org/10.3390/jcm11123287
Chicago/Turabian StyleLozano-Rodríguez, Roberto, Verónica Terrón-Arcos, Raúl López, Juan Martín-Gutiérrez, Alejandro Martín-Quirós, Charbel Maroun-Eid, Elena Muñoz del Val, Carlos Cañada-Illana, Alejandro Pascual Iglesias, Jaime Valentín Quiroga, and et al. 2022. "Differential Immune Checkpoint and Ig-like V-Type Receptor Profiles in COVID-19: Associations with Severity and Treatment" Journal of Clinical Medicine 11, no. 12: 3287. https://doi.org/10.3390/jcm11123287