The HMOX1 Pathway as a Promising Target for the Treatment and Prevention of SARS-CoV-2 of 2019 (COVID-19)



Abstract
:1. Introduction
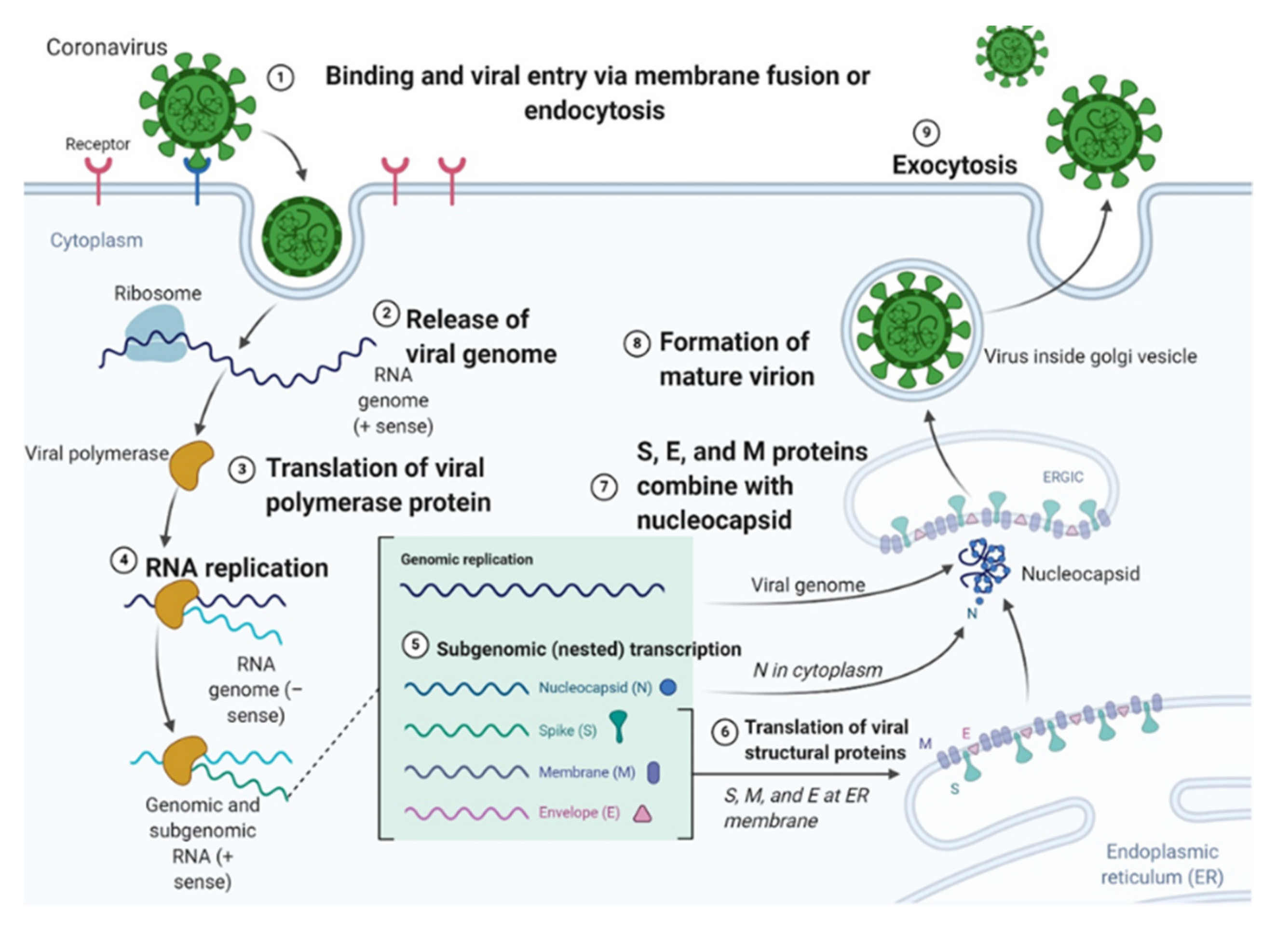
2. Life Cycle and Infectious Process of the SARS-CoV-2 Virus
3. Current Treatments for SARS-CoV-2 Patients
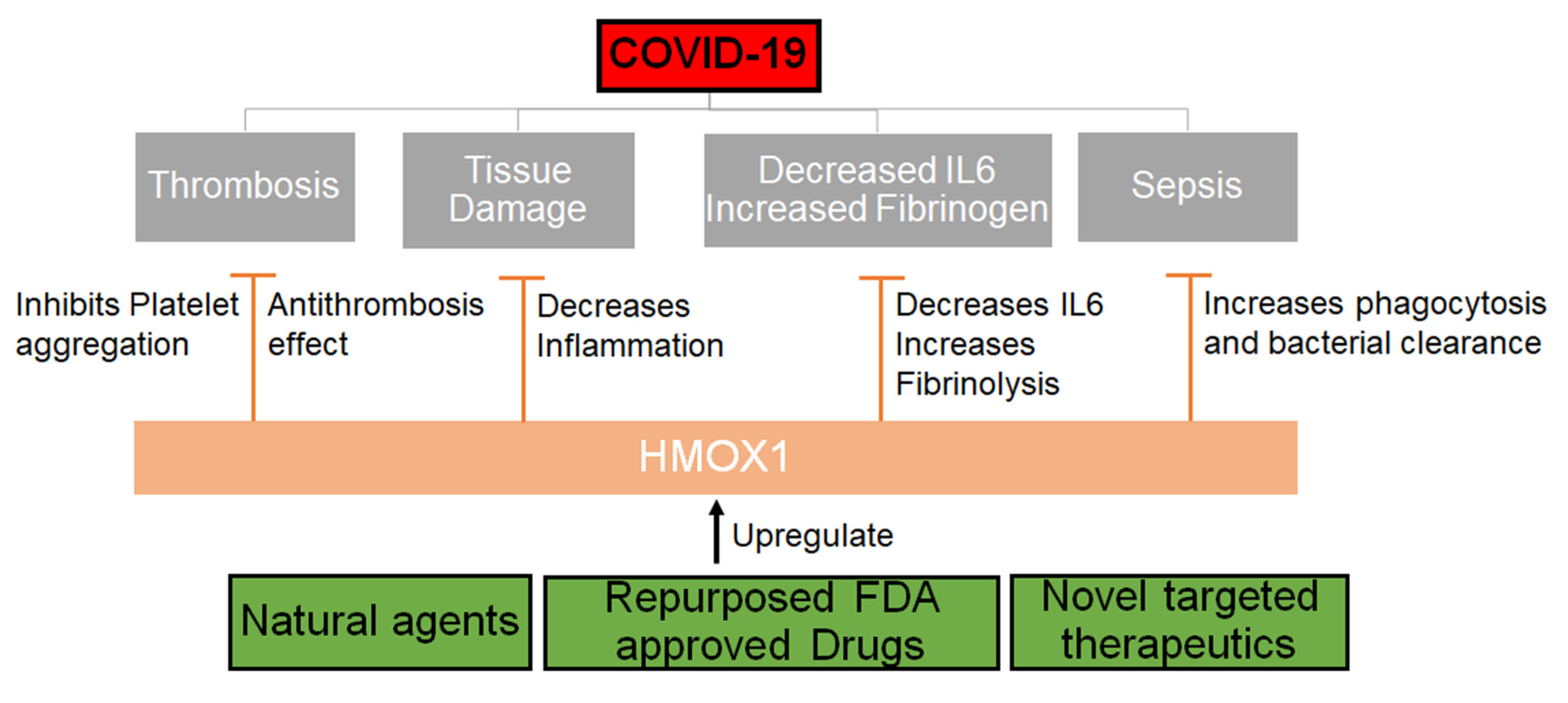
4. COVID-19 Related Thrombosis, Sepsis and Fibrinogenesis
5. Role of HMOX1 in Thrombosis, Fibrinolysis and Sepsis
6. Small Molecules for Host Directed Therapy of SARS-CoV-2
7. Natural Antiviral Agents that Upregulate the HMOX1 Pathway
8. Upregulation of Protein and mRNA Levels of HMOX1 Using Natural Compounds and Clinically Available Therapeutics
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Teuwen, L.A.; Geldhof, V.; Pasut, A.; Carmeliet, P. COVID-19: The vasculature unleashed. Nat. Rev. Immunol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons From the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72314 Cases From the Chinese Center for Disease Control and Prevention. JAMA 2020. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.; Hirsch, J.S.; Narasimhan, M.; Crawford, J.M.; McGinn, T.; Davidson, K.W.; Northwell, C.-R.C.; Barnaby, D.P.; Becker, L.B.; Chelico, J.D.; et al. Presenting Characteristics, Comorbidities, and Outcomes Among 5700 Patients Hospitalized With COVID-19 in the New York City Area. JAMA 2020. [Google Scholar] [CrossRef]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. Hlh Across Speciality Collaboration, U.K. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir-Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef]
- Magrone, T.; Magrone, M.; Jirillo, E. Focus on Receptors for Coronaviruses with Special Reference to Angiotensin-converting Enzyme 2 as a Potential Drug Target—A Perspective. Endocr. Metab. Immun. Disord. Drug Targets 2020, 20, 807–811. [Google Scholar] [CrossRef]
- Corbett, K.S.; Edwards, D.; Leist, S.R.; Abiona, O.M.; Boyoglu-Barnum, S.; Gillespie, R.A.; Himansu, S.; Schafer, A.; Ziwawo, C.T.; DiPiazza, A.T.; et al. SARS-CoV-2 mRNA Vaccine Development Enabled by Prototype Pathogen Preparedness. bioRxiv 2020. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Shirato, K.; Kawase, M.; Matsuyama, S. Wild-type human coronaviruses prefer cell-surface TMPRSS2 to endosomal cathepsins for cell entry. Virology 2018, 517, 9–15. [Google Scholar] [CrossRef] [PubMed]
- SARS-CoV-2 Life Cycle: Stages and Inhibition Targets. Available online: https://www.antibodies-online.com/resources/18/5410/sars-cov-2-life-cycle-stages-and-inhibition-targets/ (accessed on 31 March 2020).
- Deciphering the Biology of the 2019 Coronavirus. Available online: https://blog.labtag.com/deciphering-the-biology-of-the-2019-coronavirus/ (accessed on 6 February 2020).
- Cao, W.; Li, T. COVID-19: Towards understanding of pathogenesis. Cell Res. 2020, 30, 367–369. [Google Scholar] [CrossRef] [PubMed]
- Cao, X. COVID-19: Immunopathology and its implications for therapy. Nat. Rev. Immunol. 2020, 20, 269–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atal, S.; Fatima, Z. IL-6 Inhibitors in the Treatment of Serious COVID-19: A Promising Therapy? Pharm. Med. 2020, 34, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine storm and leukocyte changes in mild versus severe SARS-CoV-2 infection: Review of 3939 COVID-19 patients in China and emerging pathogenesis and therapy concepts. J. Leukoc. Biol. 2020. [Google Scholar] [CrossRef]
- Saha, A.; Sharma, A.R.; Bhattacharya, M.; Sharma, G.; Lee, S.S.; Chakraborty, C. Tocilizumab: A Therapeutic Option for the Treatment of Cytokine Storm Syndrome in COVID-19. Arch. Med. Res. 2020. [Google Scholar] [CrossRef]
- Antinori, S.; Bonazzetti, C.; Gubertini, G.; Capetti, A.; Pagani, C.; Morena, V.; Rimoldi, S.; Galimberti, L.; Sarzi-Puttini, P.; Ridolfo, A.L. Tocilizumab for cytokine storm syndrome in COVID-19 pneumonia: An increased risk for candidemia? Autoimmun. Rev. 2020, 19, 102564. [Google Scholar] [CrossRef]
- Maeda, T.; Obata, R.; Rizk, D.D.; Kuno, T. The Association of Interleukin-6 value, Interleukin inhibitors and Outcomes of Patients with COVID-19 in New York City. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Amirian, E.S.; Levy, J.K. Current knowledge about the antivirals remdesivir (GS-5734) and GS-441524 as therapeutic options for coronaviruses. One Health 2020, 9. [Google Scholar] [CrossRef]
- Winkler, U.; Jensen, M.; Manzke, O.; Schulz, H.; Diehl, V.; Engert, A. Cytokine-release syndrome in patients with B-cell chronic lymphocytic leukemia and high lymphocyte counts after treatment with an anti-CD20 monoclonal antibody (rituximab, IDEC-C2B8). Blood 1999, 94, 2217–2224. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.C.; Koh, L.P.; Tan, P. Fatal cytokine release syndrome with chimeric anti-CD20 monoclonal antibody rituximab in a 71-year-old patient with chronic lymphocytic leukemia. J. Clin. Oncol. 1999, 17, 1962–1963. [Google Scholar] [CrossRef] [PubMed]
- Wise, J. Covid-19 and thrombosis: What do we know about the risks and treatment? BMJ Brit. Med. J. 2020, 369. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; O’Meara, M.J.; Guo, J.Z.; Swaney, D.L.; Tummino, T.A.; Huttenhain, R.; et al. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef]
- Origassa, C.S.; Camara, N.O. Cytoprotective role of heme oxygenase-1 and heme degradation derived end products in liver injury. World J. Hepatol. 2013, 5, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Siu, K.L.; Yuen, K.S.; Castano-Rodriguez, C.; Ye, Z.W.; Yeung, M.L.; Fung, S.Y.; Yuan, S.; Chan, C.P.; Yuen, K.Y.; Enjuanes, L.; et al. Severe acute respiratory syndrome coronavirus ORF3a protein activates the NLRP3 inflammasome by promoting TRAF3-dependent ubiquitination of ASC. FASEB J. 2019, 33, 8865–8877. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microb. 2020, 27, 992–1000.e3. [Google Scholar] [CrossRef]
- Naito, Y.; Takagi, T.; Higashimura, Y. Heme oxygenase-1 and anti-inflammatory M2 macrophages. Arch. Biochem. Biophys. 2014, 564, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Sisay, M. Available Evidence and Ongoing Clinical Trials of Remdesivir: Could It Be a Promising Therapeutic Option for COVID-19? Front. Pharmacol. 2020, 11, 791. [Google Scholar] [CrossRef]
- Jorgensen, S.C.J.; Kebriaei, R.; Dresser, L.D. Remdesivir: Review of Pharmacology, Pre-clinical Data, and Emerging Clinical Experience for COVID-19. Pharmacotherapy 2020. [Google Scholar] [CrossRef]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the Treatment of Covid-19—Preliminary Report. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Gopel, S.; Bethge, W.; Martus, P.; Kreth, F.; Iftner, T.; Joos, S.; Dobele, S.; Mordmuller, B.; Kremsner, P.; Ettrich, T.; et al. Test and treat COVID 65 plus—Hydroxychloroquine versus placebo in early ambulatory diagnosis and treatment of older patients with COVID19: A structured summary of a study protocol for a randomised controlled trial. Trials 2020, 21, 635. [Google Scholar] [CrossRef] [PubMed]
- Vouri, S.M.; Thai, T.N.; Winterstein, A.G. An evaluation of co-use of chloroquine or hydroxychloroquine plus azithromycin on cardiac outcomes: A pharmacoepidemiological study to inform use during the COVID19 pandemic. Res. Soc. Adm. Pharm. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Chahinian, H.; Yahi, N. Synergistic antiviral effect of hydroxychloroquine and azithromycin in combination against SARS-CoV-2: What molecular dynamics studies of virus-host interactions reveal. Int. J. Antimicrob. Agents 2020. [Google Scholar] [CrossRef] [PubMed]
- Parra-Lara, L.G.; Martinez-Arboleda, J.J.; Rosso, F. Azithromycin and SARS-CoV-2 infection: Where we are now and where we are going. J. Glob. Antimicrob. Resist. 2020. [Google Scholar] [CrossRef]
- Juurlink, D.N. Safety considerations with chloroquine, hydroxychloroquine and azithromycin in the management of SARS-CoV-2 infection. CMAJ 2020, 192, E450–E453. [Google Scholar] [CrossRef] [Green Version]
- Pothen, L.; Yildiz, H.; De Greef, J.; Penaloza, A.; Beauloye, C.; Belkhir, L.; Yombi, J.C. Safety use of hydroxychloroquine and its combination with azithromycin in the context of Sars-CoV-2 outbreak: Clinical experience in a Belgian tertiary center. Travel Med. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Seftel, M.D. Additional safety consideration for azithromycin in the management of SARS-CoV-2 infection. CMAJ 2020, 192, E483. [Google Scholar] [CrossRef]
- Casadevall, A.; Joyner, M.J.; Pirofski, L.A. SARS-CoV-2 viral load and antibody responses: The case for convalescent plasma therapy. J. Clin. Investig. 2020. [Google Scholar] [CrossRef]
- Salazar, E.; Kuchipudi, S.V.; Christensen, P.A.; Eagar, T.N.; Yi, X.; Zhao, P.; Jin, Z.; Long, S.W.; Olsen, R.J.; Chen, J.; et al. Relationship between Anti-Spike Protein Antibody Titers and SARS-CoV-2 In Vitro Virus Neutralization in Convalescent Plasma. bioRxiv 2020. [Google Scholar] [CrossRef]
- Mira, E.; Yarce, O.A.; Ortega, C.; Fernandez, S.; Pascual, N.M.; Gomez, C.; Alvarez, M.A.; Molina, I.J.; Lama, R.; Santamaria, M. Rapid recovery of a SARS-CoV-2-infected X-linked agammaglobulinemia patient after infusion of COVID-19 convalescent plasma. J. Allergy Clin. Immunol. Pract. 2020. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, A. Plasma from donors convalescent from SARS-CoV-2 infection-A matter of priorities. Transfus. Clin. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, C.M.; Cummings, M.J.; Rajagopalan, K.N.; Borden, S.; Bitan, Z.C.; Wolf, A.; Kantor, A.; Briese, T.; Meyer, B.J.; Jacobson, S.D.; et al. Evaluating the efficacy and safety of human anti-SARS-CoV-2 convalescent plasma in severely ill adults with COVID-19: A structured summary of a study protocol for a randomized controlled trial. Trials 2020, 21, 499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Pang, R.; Xue, X.; Bao, J.; Ye, S.; Dai, Y.; Zheng, Y.; Fu, Q.; Hu, Z.; Yi, Y. Anti-SARS-CoV-2 virus antibody levels in convalescent plasma of six donors who have recovered from COVID-19. Aging 2020, 12, 6536–6542. [Google Scholar] [CrossRef]
- Morillas, J.A.; Canosa, F.M.; Srinivas, P.; Asadi, T.; Calabrese, C.; Rajendram, P.; Budev, M.; Poggio, E.D.; Menon, K.V.N.; Gastman, B.; et al. Tocilizumab therapy in 5 solid and composite tissue transplant recipients with early ARDS due to SARS-CoV-2. Am. J. Transpl. 2020. [Google Scholar] [CrossRef]
- Shabani, M.; Shokouhi, S.; Moradi, O.; Saffaei, A.; Sahraei, Z. Tocilizumab administration in patients with SARS-CoV-2 infection: Subcutaneous injection vs intravenous infusion. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Magro, G. SARS-CoV-2 and COVID-19: Is interleukin-6 (IL-6) the ‘culprit lesion‘ of ARDS onset? What is there besides Tocilizumab? SGP130Fc. Cytokine X 2020. [Google Scholar] [CrossRef]
- Quartuccio, L.; Sonaglia, A.; McGonagle, D.; Fabris, M.; Peghin, M.; Pecori, D.; Monte, A.; Bove, T.; Curcio, F.; Bassi, F.; et al. Profiling COVID-19 pneumonia progressing into the cytokine storm syndrome: Results from a single Italian Centre study on tocilizumab versus standard of care. J. Clin. Virol. 2020. [Google Scholar] [CrossRef]
- Antony, S.J.; Davis, M.A.; Davis, M.G.; Almaghlouth, N.K.; Guevara, R.; Omar, F.; Del Rey, F.; Hassan, A.; Arian, M.U.; Antony, N.; et al. Early use of tocilizumab in the prevention of adult respiratory failure in SARS-CoV-2 infections and the utilization of interleukin-6 levels in the management. J. Med. Virol. 2020. [Google Scholar] [CrossRef]
- Costanzo, M.; De Giglio, M.A.R.; Roviello, G.N. SARS-CoV-2: Recent Reports on Antiviral Therapies Based on Lopinavir/Ritonavir, Darunavir/Umifenovir, Hydroxychloroquine, Remdesivir, Favipiravir and Other Drugs for the Treatment of the New Coronavirus. Curr. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Smolders, E.J.; Te Brake, L.H.; Burger, D.M. SARS-CoV-2 and HIV protease inhibitors: Why lopinavir/ritonavir will not work for COVID-19 infection. Antivir. Ther. 2020. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Lee, Y.L.; Chen, C.P.; Lin, Y.C.; Liu, C.E.; Liao, C.H.; Cheng, S.H. Lopinavir/ritonavir did not shorten the duration of SARS CoV-2 shedding in patients with mild pneumonia in Taiwan. J. Microbiol. Immunol. Infect. 2020, 53, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Hraiech, S.; Bourenne, J.; Kuteifan, K.; Helms, J.; Carvelli, J.; Gainnier, M.; Meziani, F.; Papazian, L. Lack of viral clearance by the combination of hydroxychloroquine and azithromycin or lopinavir and ritonavir in SARS-CoV-2-related acute respiratory distress syndrome. Ann. Intensive Care 2020, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Liu, X.Y.; Zhu, Y.N.; Huang, L.; Dan, B.T.; Zhang, G.J.; Gao, Y.H. Factors associated with prolonged viral shedding and impact of lopinavir/ritonavir treatment in hospitalised non-critically ill patients with SARS-CoV-2 infection. Eur. Respir. J. 2020, 56. [Google Scholar] [CrossRef] [PubMed]
- Muralidharan, N.; Sakthivel, R.; Velmurugan, D.; Gromiha, M.M. Computational studies of drug repurposing and synergism of lopinavir, oseltamivir and ritonavir binding with SARS-CoV-2 protease against COVID-19. J. Biomol. Struct. Dyn. 2020, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lecronier, M.; Beurton, A.; Burrel, S.; Haudebourg, L.; Deleris, R.; Le Marec, J.; Virolle, S.; Nemlaghi, S.; Bureau, C.; Mora, P.; et al. Comparison of hydroxychloroquine, lopinavir/ritonavir, and standard of care in critically ill patients with SARS-CoV-2 pneumonia: An opportunistic retrospective analysis. Crit. Care 2020, 24, 418. [Google Scholar] [CrossRef] [PubMed]
- Denholm, J.T.; Davis, J.; Paterson, D.; Roberts, J.; Morpeth, S.; Snelling, T.; Zentner, D.; Rees, M.; O’Sullivan, M.; Price, D.; et al. The Australasian COVID-19 Trial (ASCOT) to assess clinical outcomes in hospitalised patients with SARS-CoV-2 infection (COVID-19) treated with lopinavir/ritonavir and/or hydroxychloroquine compared to standard of care: A structured summary of a study protocol for a randomised controlled trial. Trials 2020, 21, 646. [Google Scholar] [CrossRef]
- Du, Y.X.; Chen, X.P. Response to “Dose Rationale for Favipiravir Use in Patients Infected With SARS-CoV-2”. Clin. Pharmacol. Ther. 2020, 108, 190. [Google Scholar] [CrossRef]
- Shannon, A.; Selisko, B.; Le, N.; Huchting, J.; Touret, F.; Piorkowski, G.; Fattorini, V.; Ferron, F.; Decroly, E.; Meier, C.; et al. Favipiravir strikes the SARS-CoV-2 at its Achilles heel, the RNA polymerase. bioRxiv 2020. [Google Scholar] [CrossRef]
- Eloy, P.; Solas, C.; Touret, F.; Mentre, F.; Malvy, D.; de Lamballerie, X.; Guedj, J. Dose Rationale for Favipiravir Use in Patients Infected With SARS-CoV-2. Clin. Pharmacol. Ther. 2020, 108, 188. [Google Scholar] [CrossRef]
- Gendelman, O.; Amital, H.; Bragazzi, N.L.; Watad, A.; Chodick, G. Continuous hydroxychloroquine or colchicine therapy does not prevent infection with SARS-CoV-2: Insights from a large healthcare database analysis. Autoimmun. Rev. 2020, 19, 102566. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.M.; Levy, J.H. COVID-19 and its implications for thrombosis and anticoagulation. Blood 2020, 135, 2033–2040. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, M.; Thachil, J.; Iba, T.; Levy, J.H. Coagulation abnormalities and thrombosis in patients with COVID-19. Lancet. Haematol. 2020, 7, e438–e440. [Google Scholar] [CrossRef]
- Nahum, J.; Morichau-Beauchant, T.; Daviaud, F.; Echegut, P.; Fichet, J.M.; Maillet, J.M.; Thierry, S. Venous Thrombosis Among Critically Ill Patients With Coronavirus Disease 2019 (COVID-19). JAMA Netw. Open 2020, 3. [Google Scholar] [CrossRef]
- Lee, T.S.; Chau, L.Y. Heme oxygenase-1 mediates the anti-inflammatory effect of interleukin-10 in mice. Nat. Med. 2002, 8, 240–246. [Google Scholar] [CrossRef]
- Du, Y.; Zhang, H.; Xu, Y.; Ding, Y.; Chen, X.R.; Mei, Z.F.; Ding, H.Y.; Jie, Z.J. Association among genetic polymorphisms of GSTP1, HO-1, and SOD-3 and chronic obstructive pulmonary disease susceptibility. Int. J. Chronic. Obstr. 2019, 14, 2081–2088. [Google Scholar] [CrossRef] [Green Version]
- Ayer, A.; Zarjou, A.; Agarwal, A.; Stocker, R. Heme Oxygenases in Cardiovascular Health and Disease. Physiol. Rev. 2016, 96, 1449–1508. [Google Scholar] [CrossRef]
- Schmidt, W.N.; Mathahs, M.M.; Zhu, Z. Heme and HO-1 Inhibition of HCV, HBV, and HIV. Front. Pharmacol. 2012, 3, 129. [Google Scholar] [CrossRef] [Green Version]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiol. Mol. Biol. R. 2012, 76, 16–32. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Su, W.; Yu, Q.; Zhang, M.; Di, C.; Lin, X.; Wu, M.; Xia, Z. Heme oxygenase-1 protects airway epithelium against apoptosis by targeting the proinflammatory NLRP3-RXR axis in asthma. J. Biol. Chem. 2018, 293, 18454–18465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, R.; Reed, J.C. ER stress-induced cell death mechanisms. BBA Mol. Cell Res. 2013, 1833, 3460–3470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, I.Y.; Moriyama, M.; Chang, M.F.; Ichinohe, T. Severe Acute Respiratory Syndrome Coronavirus Viroporin 3a Activates the NLRP3 Inflammasome. Front. Microbiol. 2019, 10, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, D.M.; Sorum, B.; Hoel, C.M.; Sridharan, S.; Remis, J.P.; Toso, D.B.; Brohawn, S.G. Cryo-EM structure of the SARS-CoV-2 3a ion channel in lipid nanodiscs. bioRxiv 2020. [Google Scholar] [CrossRef]
- Lu, W.; Zheng, B.J.; Xu, K.; Schwarz, W.; Du, L.; Wong, C.K.; Chen, J.; Duan, S.; Deubel, V.; Sun, B. Severe acute respiratory syndrome-associated coronavirus 3a protein forms an ion channel and modulates virus release. Proc. Natl. Acad. Sci. USA 2006, 103, 12540–12545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Shu, T.; Wu, D.; Mu, J.; Wang, C.; Huang, M.; Han, Y.; Zhang, X.Y.; Zhou, W.; Qiu, Y.; et al. The ORF3a protein of SARS-CoV-2 induces apoptosis in cells. Cell Mol. Immunol. 2020. [Google Scholar] [CrossRef]
- Yue, Y.; Nabar, N.R.; Shi, C.S.; Kamenyeva, O.; Xiao, X.; Hwang, I.Y.; Wang, M.; Kehrl, J.H. SARS-Coronavirus Open Reading Frame-3a drives multimodal necrotic cell death. Cell Death Dis. 2018, 9, 904. [Google Scholar] [CrossRef]
- Peng, L.; Mundada, L.; Stomel, J.M.; Liu, J.J.; Sun, J.; Yet, S.F.; Fay, W.P. Induction of heme oxygenase-1 expression inhibits platelet-dependent thrombosis. Antioxid. Redox. Signal. 2004, 6, 729–735. [Google Scholar] [CrossRef]
- True, A.L.; Olive, M.; Boehm, M.; San, H.; Westrick, R.J.; Raghavachari, N.; Xu, X.; Lynn, E.G.; Sack, M.N.; Munson, P.J.; et al. Heme oxygenase-1 deficiency accelerates formation of arterial thrombosis through oxidative damage to the endothelium, which is rescued by inhaled carbon monoxide. Circ. Res. 2007, 101, 893–901. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, S.; Weltermann, A.; Fritsche, R.; Marsik, C.; Wagner, O.; Kyrle, P.A.; Eichinger, S. Genetic variation in heme oxygenase 1 (HMOX1) and the risk of recurrent venous thromboembolism. J. Vasc. Surg. 2008, 47, 566–570. [Google Scholar] [CrossRef] [Green Version]
- Bach, F.H. Heme oxygenase-1 as a protective gene. Wien. Klin. Wochenschr. 2002, 114, 1–3. [Google Scholar] [PubMed]
- Lindenblatt, N.; Bordel, R.; Schareck, W.; Menger, M.D.; Vollmar, B. Vascular heme oxygenase-1 induction suppresses microvascular thrombus formation in vivo. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gende, O.A. Carbon monoxide inhibits capacitative calcium entry in human platelets. Thromb. Res. 2004, 114, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Toda, K.; Karimova, A.; Yan, S.F.; Naka, Y.; Yet, S.F.; Pinsky, D.J. Paradoxical rescue from ischemic lung injury by inhaled carbon monoxide driven by derepression of fibrinolysis. Nat. Med. 2001, 7, 598–604. [Google Scholar] [CrossRef]
- Durante, W. Carbon monoxide and bile pigments: Surprising mediators of vascular function. Vasc. Med. 2002, 7, 195–202. [Google Scholar] [CrossRef]
- Soares, M.P.; Usheva, A.; Brouard, S.; Berberat, P.O.; Gunther, L.; Tobiasch, E.; Bach, F.H. Modulation of endothelial cell apoptosis by heme oxygenase-1-derived carbon monoxide. Antioxid. Redox. Signal 2002, 4, 321–329. [Google Scholar] [CrossRef]
- Chung, S.W.; Liu, X.; Macias, A.A.; Baron, R.M.; Perrella, M.A. Heme oxygenase-1-derived carbon monoxide enhances the host defense response to microbial sepsis in mice. J. Clin. Investig. 2008, 118, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Otterbein, L.E.; May, A.; Chin, B.Y. Carbon monoxide increases macrophage bacterial clearance through Toll-like receptor (TLR)4 expression. Cell Mol. Biol. 2005, 51, 433–440. [Google Scholar]
- Overhaus, M.; Moore, B.A.; Barbato, J.E.; Behrendt, F.F.; Doering, J.G.; Bauer, A.J. Biliverdin protects against polymicrobial sepsis by modulating inflammatory mediators. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G695–G703. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.P.; Jiang, L.; Kang, K.; Fei, D.S.; Meng, X.L.; Nan, C.C.; Pan, S.H.; Zhao, M.R.; Zhao, M.Y. Hemin inhibits NLRP3 inflammasome activation in sepsis-induced acute lung injury, involving heme oxygenase-1. Int. Immunopharmacol. 2014, 20, 24–32. [Google Scholar] [CrossRef]
- Uddin, M.J.; Joe, Y.; Kim, S.K.; Oh Jeong, S.; Ryter, S.W.; Pae, H.O.; Chung, H.T. IRG1 induced by heme oxygenase-1/carbon monoxide inhibits LPS-mediated sepsis and pro-inflammatory cytokine production. Cell Mol. Immunol. 2016, 13, 170–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carchman, E.H.; Rao, J.; Loughran, P.A.; Rosengart, M.R.; Zuckerbraun, B.S. Heme oxygenase-1-mediated autophagy protects against hepatocyte cell death and hepatic injury from infection/sepsis in mice. Hepatology 2011, 53, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Boi, M.; Gaudio, E.; Bonetti, P.; Kwee, I.; Bernasconi, E.; Tarantelli, C.; Rinaldi, A.; Testoni, M.; Cascione, L.; Ponzoni, M.; et al. The BET Bromodomain Inhibitor OTX015 Affects Pathogenetic Pathways in Preclinical B-cell Tumor Models and Synergizes with Targeted Drugs. Clin. Cancer Res. 2015, 21, 1628–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Song, S.; Han, H.; Xu, H.; Fan, G.; Qian, C.; Qiu, Y.; Zhou, W.; Zhuang, W.; Li, B. The BET Bromodomain Inhibitor OTX015 Synergizes with Targeted Agents in Multiple Myeloma. Mol. Pharm. 2018, 15, 5387–5396. [Google Scholar] [CrossRef]
- Donati, B.; Lorenzini, E.; Ciarrocchi, A. BRD4 and Cancer: Going beyond transcriptional regulation. Molecular Cancer 2018, 17. [Google Scholar] [CrossRef]
- Zhou, B.; Hu, J.; Xu, F.; Chen, Z.; Bai, L.; Fernandez-Salas, E.; Lin, M.; Liu, L.; Yang, C.Y.; Zhao, Y.; et al. Discovery of a Small-Molecule Degrader of Bromodomain and Extra-Terminal (BET) Proteins with Picomolar Cellular Potencies and Capable of Achieving Tumor Regression. J. Med. Chem. 2018, 61, 462–481. [Google Scholar] [CrossRef]
- Di Costanzo, A.; Del Gaudio, N.; Migliaccio, A.; Altucci, L. Epigenetic drugs against cancer: An evolving landscape. Arch. Toxicol. 2014, 88, 1651–1668. [Google Scholar] [CrossRef]
- Lu, J.; Qian, Y.; Altieri, M.; Dong, H.; Wang, J.; Raina, K.; Hines, J.; Winkler, J.D.; Crew, A.P.; Coleman, K.; et al. Hijacking the E3 Ubiquitin Ligase Cereblon to Efficiently Target BRD4. Chem. Biol. 2015, 22, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Winter, G.E.; Buckley, D.L.; Paulk, J.; Roberts, J.M.; Souza, A.; Dhe-Paganon, S.; Bradner, J.E. Phthalimide conjugation as a strategy for in vivo target protein degradation. Science 2015, 348, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, R.; Licandro, S.A.; Astorgues-Xerri, L.; Lettera, E.; Panini, N.; Romano, M.; Erba, E.; Ubezio, P.; Bello, E.; Libener, R.; et al. Promising in vivo efficacy of the BET bromodomain inhibitor OTX015/MK-8628 in malignant pleural mesothelioma xenografts. Int. J. Cancer 2017, 140, 197–207. [Google Scholar] [CrossRef]
- Gandhi, V.; Plunkett, W.; Cortes, J.E. Omacetaxine: A protein translation inhibitor for treatment of chronic myelogenous leukemia. Clin. Cancer Res. 2014, 20, 1735–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobos, E.J.; Entrena, J.M.; Nieto, F.R.; Cendan, C.M.; Del Pozo, E. Pharmacology and therapeutic potential of sigma(1) receptor ligands. Curr. Neuropharmacol. 2008, 6, 344–366. [Google Scholar] [CrossRef] [PubMed]
- Maurice, T.; Su, T.P. The pharmacology of sigma-1 receptors. Pharmacol. Ther. 2009, 124, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vavers, E.; Zvejniece, L.; Maurice, T.; Dambrova, M. Allosteric Modulators of Sigma-1 Receptor: A Review. Front. Pharmacol. 2019, 10, 223. [Google Scholar] [CrossRef]
- Patel, M.J.; Tripathy, S.; Mukhopadhyay, K.D.; Wangjam, T.; Cabang, A.B.; Morris, J.; Wargovich, M.J. A supercritical CO2 extract of neem leaf (A. indica) and its bioactive liminoid, nimbolide, suppresses colon cancer in preclinical models by modulating pro-inflammatory pathways. Mol. Carcinog. 2018, 57, 1156–1165. [Google Scholar] [CrossRef]
- Mahapatra, S.; Young, C.Y.; Kohli, M.; Karnes, R.J.; Klee, E.W.; Holmes, M.W.; Tindall, D.J.; Donkena, K.V. Antiangiogenic Effects and Therapeutic Targets of Azadirachta indica Leaf Extract in Endothelial Cells. Evid. Based Complement. Altern. Med. 2012, 2012, 303019. [Google Scholar] [CrossRef] [Green Version]
- Morris, J.; Gonzales, C.B.; De La Chapa, J.J.; Cabang, A.B.; Fountzilas, C.; Patel, M.; Orozco, S.; Wargovich, M.J. The Highly Pure Neem Leaf Extract, SCNE, Inhibits Tumorigenesis in Oral Squamous Cell Carcinoma via Disruption of Pro-tumor Inflammatory Cytokines and Cell Signaling. Front. Oncol. 2019, 9, 890. [Google Scholar] [CrossRef] [Green Version]
- Gautam, M.K.; Gangwar, M.; Singh, S.K.; Goel, R.K. Effects of Azardirachta indica on Vascular Endothelial Growth Factor and Cytokines in Diabetic Deep Wound. Planta Med. 2015, 81, 713–721. [Google Scholar] [CrossRef]
- Natarajan, V.; Pushkala, S.; Karuppiah, V.P.; Prasad, P.V. Anti dermatophytic activity of Azardirachta indica (neem) by invitro study. Indian J. Pathol. Microbiol. 2002, 45, 311–313. [Google Scholar]
- Alzohairy, M.A. Therapeutics Role of Azadirachta indica (Neem) and Their Active Constituents in Diseases Prevention and Treatment. Evid. Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [Green Version]
- Spradlin, J.N.; Hu, X.; Ward, C.C.; Brittain, S.M.; Jones, M.D.; Ou, L.; To, M.; Proudfoot, A.; Ornelas, E.; Woldegiorgis, M.; et al. Harnessing the anti-cancer natural product nimbolide for targeted protein degradation. Nat. Chem. Biol. 2019, 15, 747–755. [Google Scholar] [CrossRef]
- Tiwari, V.; Darmani, N.A.; Yue, B.Y.; Shukla, D. In vitro antiviral activity of neem (Azardirachta indica L.) bark extract against herpes simplex virus type-1 infection. Phytother. Res. 2010, 24, 1132–1140. [Google Scholar] [CrossRef] [Green Version]
- Parida, M.M.; Upadhyay, C.; Pandya, G.; Jana, A.M. Inhibitory potential of neem (Azadirachta indica Juss) leaves on dengue virus type-2 replication. J. Ethnopharmacol. 2002, 79, 273–278. [Google Scholar] [CrossRef]
- Faccin-Galhardi, L.C.; Yamamoto, K.A.; Ray, S.; Ray, B.; Linhares, R.E.C.; Nozawa, C. The in vitro antiviral property of Azadirachta indica polysaccharides for poliovirus. J. Ethnopharmacol. 2012, 142, 86–90. [Google Scholar] [CrossRef]
- Badam, L.; Joshi, S.P.; Bedekar, S.S. ‘In vitro’ antiviral activity of neem (Azadirachta indica. A. Juss) leaf extract against group B coxsackieviruses. J. Commun. Dis. 1999, 31, 79–90. [Google Scholar] [PubMed]
- Saha, S.; Galhardi, L.C.; Yamamoto, K.A.; Linhares, R.E.; Bandyopadhyay, S.S.; Sinha, S.; Nozawa, C.; Ray, B. Water-extracted polysaccharides from Azadirachta indica leaves: Structural features, chemical modification and anti-bovine herpesvirus type 1 (BoHV-1) activity. Int. J. Biol. Macromol. 2010, 47, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Song, X.; Yin, Z.Q.; Cheng, A.C.; Jia, R.Y.; Deng, Y.X.; Ye, K.C.; Shi, C.F.; Lv, C.; Zhang, W. Antiviral activity and mode of action of extracts from neem seed kernel against duck plague virus in vitro1. Poult. Sci. 2012, 91, 2802–2807. [Google Scholar] [CrossRef] [PubMed]
- Anyaehie, U.B. Medicinal properties of fractionated acetone/water neem [Azadirachta indica] leaf extract from Nigeria: A review. Niger. J. Physiol. Sci. 2009, 24, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, S.; Karnes, R.J.; Holmes, M.W.; Young, C.Y.; Cheville, J.C.; Kohli, M.; Klee, E.W.; Tindall, D.J.; Donkena, K.V. Novel molecular targets of Azadirachta indica associated with inhibition of tumor growth in prostate cancer. AAPS J. 2011, 13, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkotoky, S.; Banerjee, M. A computational prediction of SARS-CoV-2 structural protein inhibitors from Azadirachta indica (Neem). J. Biomol. Struct. Dyn. 2020, 1–17. [Google Scholar] [CrossRef]
- Aouidate, A.; Ghaleb, A.; Chtita, S.; Aarjane, M.; Ousaa, A.; Maghat, H.; Sbai, A.; Choukrad, M.; Bouachrine, M.; Lakhlifi, T. Identification of a novel dual-target scaffold for 3CLpro and RdRp proteins of SARS-CoV-2 using 3D-similarity search, molecular docking, molecular dynamics and ADMET evaluation. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Oja, A.E.; Saris, A.; Ghandour, C.A.; Kragten, N.A.; Hogema, B.M.; Nossent, E.J.; Vrielink, H. Divergent SARS-CoV-2-specific T and B cell responses in severe but not mild COVID-19. bioRxiv 2020, 159202. Available online: https://doi.org/10.1101/2020.06.18.159202 (accessed on 18 June 2020).



{kind=link}
{kind=link}
| Potential Drug | Mode of Action | Current Status | Reference |
|---|---|---|---|
| Remdesivir | Remdesivir (a variant of Adenosine) which hinders the coronavirus RNA polymerase—a key enzyme that coronavirus requires to replicate its genetic material and proliferate in a human body. | On 1 May 2020, the FDA issued an emergency use authorization (EUA) for remdesivir | [32,33] |
| Hydroxychloroquine and chloroquine | Hydroxychloroquine and chloroquine work via changing the chemical environment of human cell membranes, consequently not allowing the virus to enter and multiply inside the cells. | FDA has issued a caution that hydroxychloroquine (HCQ) or chloroquine should not be used outside of a hospital setting/clinical trials for COVID-19 due to serious heart-related side effects. | [34,35] |
| Azithromycin | Azithromycin is an antibiotic used to treat bacterial infections like bronchitis and pneumonia. | Azithromycin is FDA approved but should not be used in combination with hydroxychloroquine together due to serious side effect concerns. | [36,37,38,39,40] |
| Convalescent plasma and antibody-based therapies | Convalescent plasma is taken from people who have developed antibodies for COVID-19. This could potentially help fight the coronavirus infection in new patients. Synthetically produced antibodies can inhibit viral infection and pathogenesis. | The FDA issued an Emergency Investigational New Drug approval for the use of convalescent plasma to treat people with COVID-19 on 24 March 2020. | [41,42,43,44,45,46] (antibody refs) |
| Actemra (tocilizumab) | Actemra blocks interleukin-6 (IL-6), a cytokine involved in human immune response. | The efficacy and safety phase 2 trial of Tocilizumab for the treatment of COVID-19 is under way. | [19,47,48,49,50,51] |
| Kaletra (lopinavir/ritonavir) | KALETRA is a combination of lopinavir and ritonavir. Lopinavir is a potent inhibitor of the Human Immunodeficiency Virus (HIV) protease. Ritonavir obstructs the CYP3A-mediated metabolism of lopinavir, resulting in increased plasma levels of lopinavir. | A cluster randomized controlled trial (RCT) of oral Kaletra (lopinavir/ritonavir) as Post-Exposure Prophylaxis (PEP) is underway for COVID-19. | [52,53,54,55,56,57,58,59] |
| Tamiflu (oseltamivir) | Oseltamivir works by inhibiting the viral neuraminidase enzyme activity. The enzyme is found on the surface of the virus (H1N1). | Several ongoing clinical trials are looking at Tamiflu in combination with other medications for COVID-19. | [52,57] |
| Avigan (favipiravir) | Favipiravir/Avigan induces a rapid mutation rate of the virus RNA polymerase complex, which results in a large proportion of inactive viruses amongst the virus population. | Favipiravir/Avigan is an approved drug in Japan and China against flu. Clinical trial is underway in US to start in Boston. | [52,60,61,62] |
| Colcrys (colchicine) | Colchicine could work if the immune system becomes too activated and a cytokine storm occurs. | [63] |
| Neem Plant Part | Virus Type | Reference |
|---|---|---|
| Aqueous extract preparation from the barks of neem (NBE) | Herpes Simplex Virus 1 (HSV-1) | [114] |
| Crude aqueous extract of neem leaves and pure neem compound (Azadirachtin) | Dengue virus type 2 | [115] |
| Neem’s polysaccharides extracted from leaves | Poliovirus | [116] |
| Neem leaf extract | Coxsackie virus B-4 | [117] |
| Water-extracted polysaccharides from neem leaves | Anti-bovine herpesvirus Type 1 | [118] |
| Neem seed kernel extracts | Duck plague virus | [119] |
| Fractionated neem-leaf extract | Human Immunodeficiency Virus 1 (HIV-1) | [120] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batra, N.; De Souza, C.; Batra, J.; Raetz, A.G.; Yu, A.-M. The HMOX1 Pathway as a Promising Target for the Treatment and Prevention of SARS-CoV-2 of 2019 (COVID-19). Int. J. Mol. Sci. 2020, 21, 6412. https://doi.org/10.3390/ijms21176412
Batra N, De Souza C, Batra J, Raetz AG, Yu A-M. The HMOX1 Pathway as a Promising Target for the Treatment and Prevention of SARS-CoV-2 of 2019 (COVID-19). International Journal of Molecular Sciences. 2020; 21(17):6412. https://doi.org/10.3390/ijms21176412
Chicago/Turabian StyleBatra, Neelu, Cristabelle De Souza, Jyoti Batra, Alan G. Raetz, and Ai-Ming Yu. 2020. "The HMOX1 Pathway as a Promising Target for the Treatment and Prevention of SARS-CoV-2 of 2019 (COVID-19)" International Journal of Molecular Sciences 21, no. 17: 6412. https://doi.org/10.3390/ijms21176412