
Epidemiological and Immunological Features of Obesity and SARS-CoV-2

 , , , , , , add
Show full author list
, , , , , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Setting and Study Population
2.2. Covariates
2.3. Laboratory Analyses
2.4. Data Classification and Analyses
3. Results
3.1. BMI and Serostatus
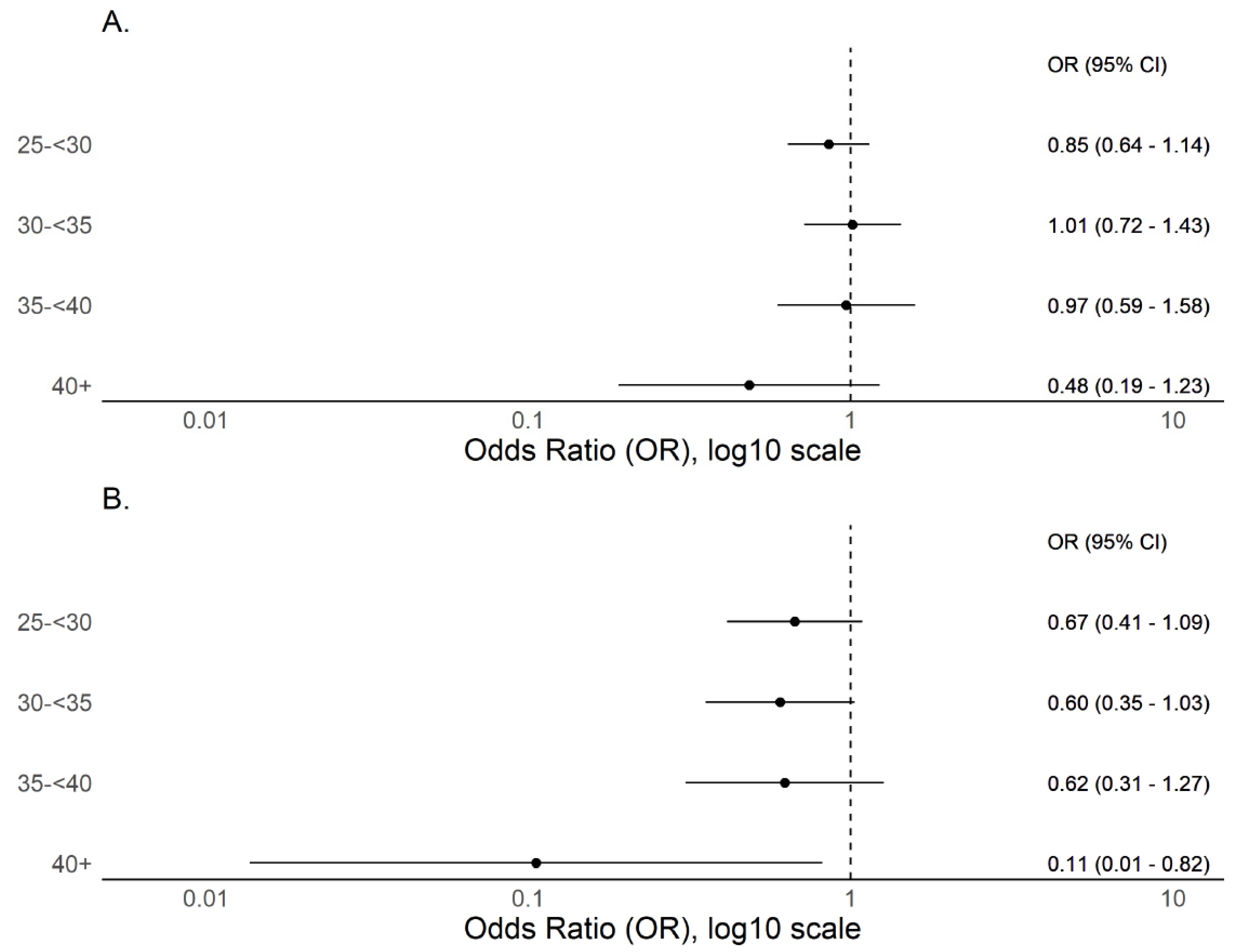
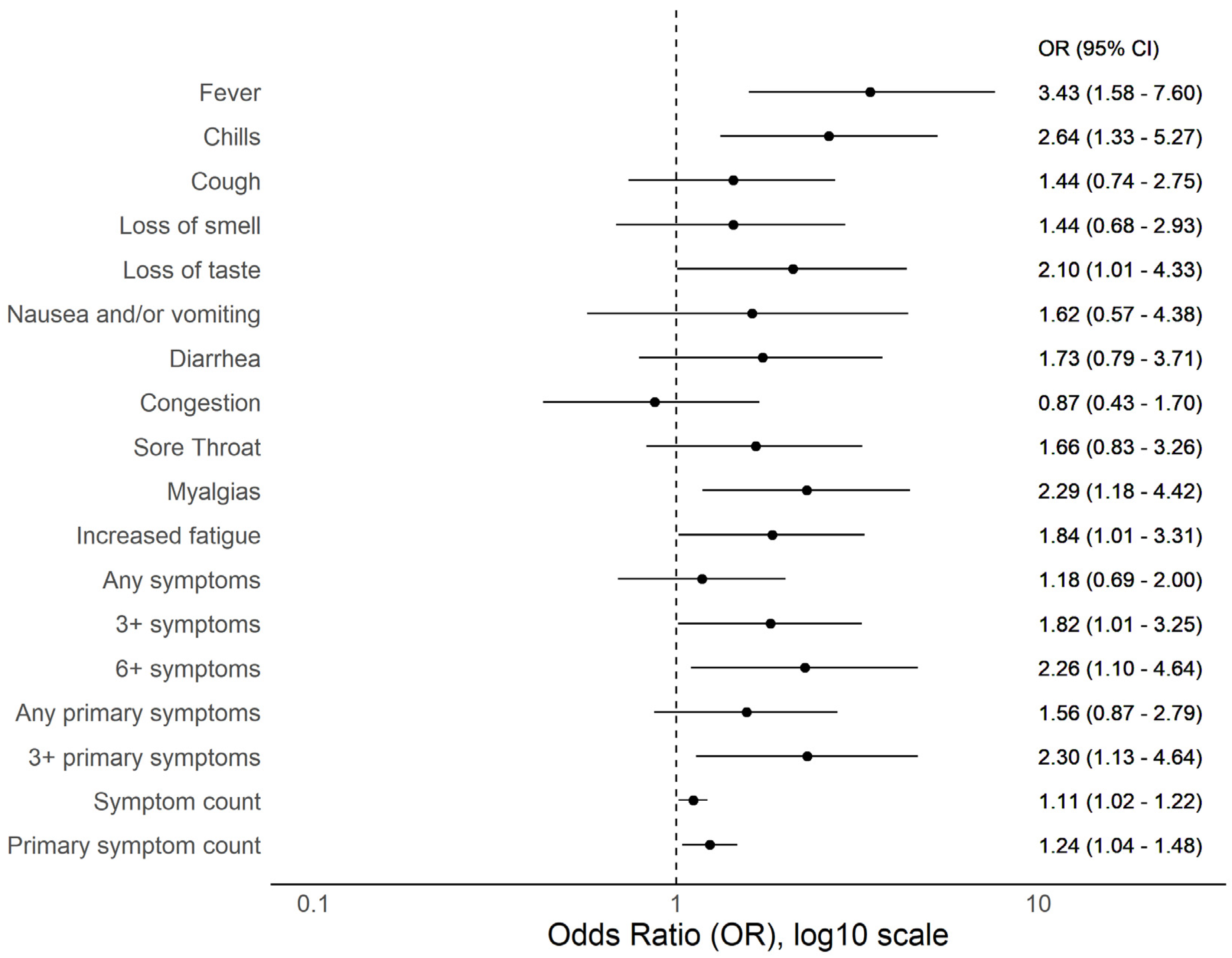
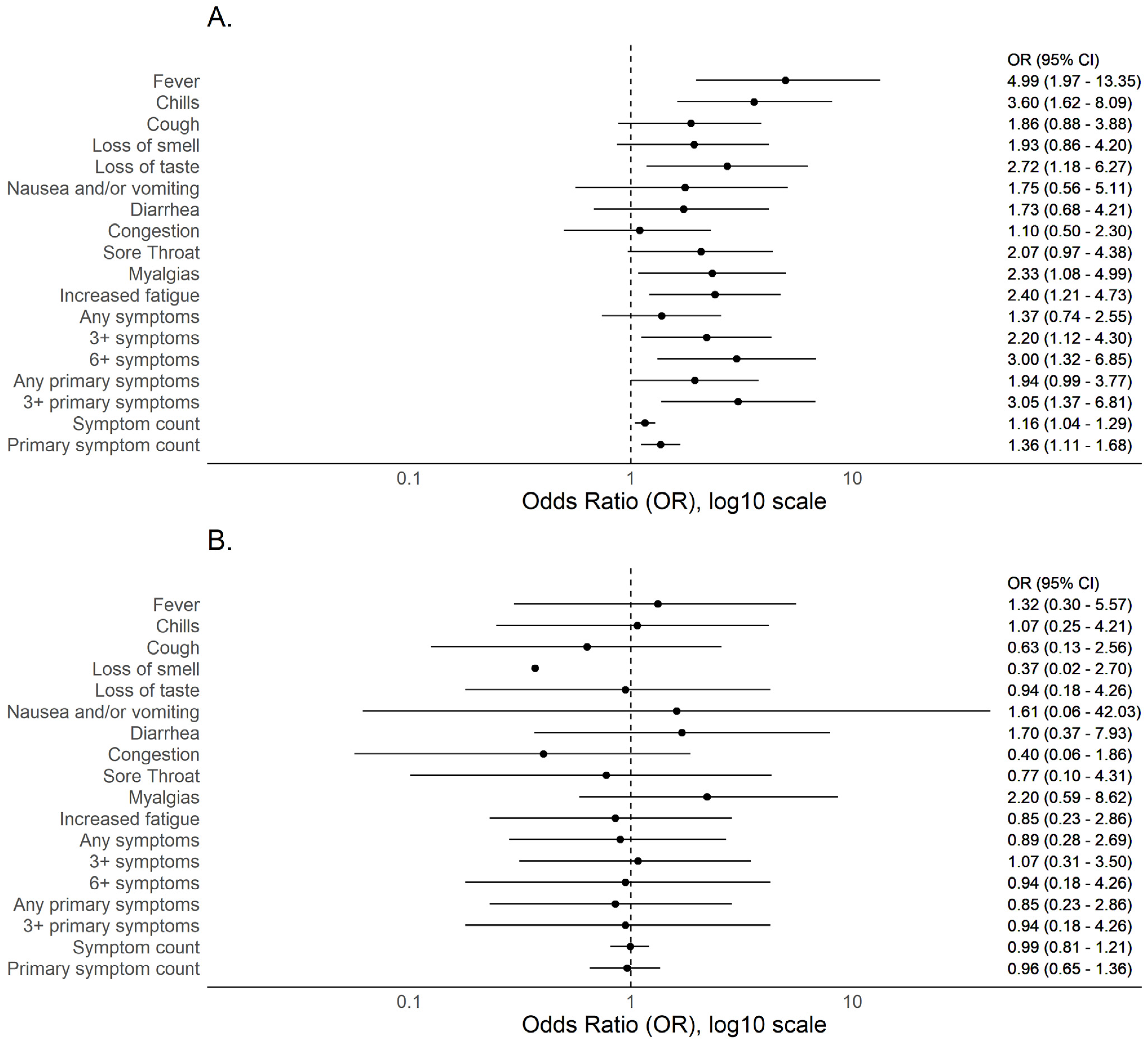
3.2. BMI and COVID-19 Compatible Symptoms
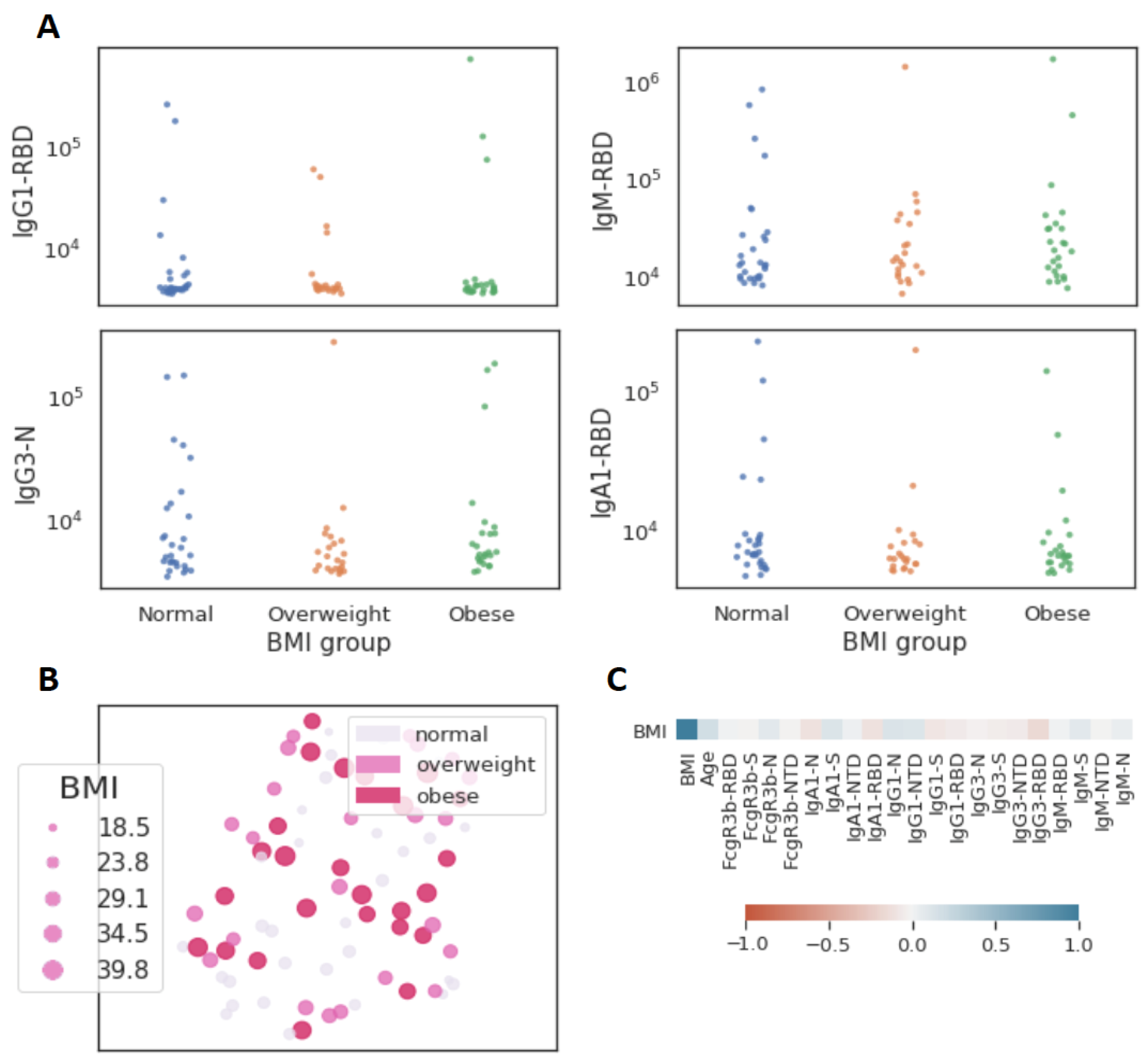
3.3. Obesity and Functional Immune Response
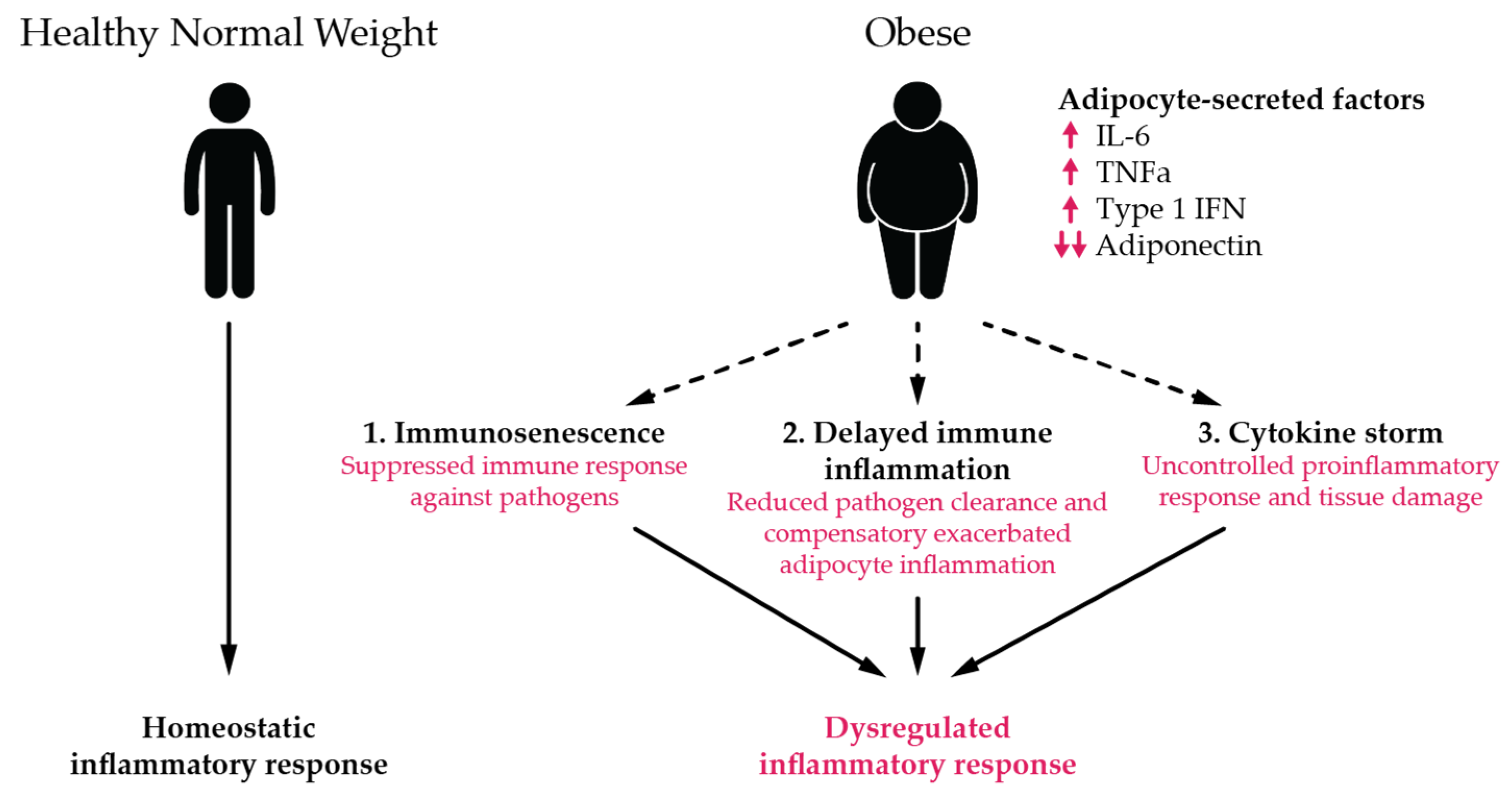
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Hamer, M.; Gale, C.R.; Kivimäki, M.; Batty, G.D. Overweight, obesity, and risk of hospitalization for COVID-19: A community-based cohort study of adults in the United Kingdom. Proc. Natl Acad Sci. USA 2020, 117, 21011–21013. [Google Scholar] [CrossRef] [PubMed]
- Seidu, S.; Gillies, C.; Zaccardi, F.; Kunutsor, S.K.; Hartmann-Boyce, J.; Yates, T. The impact of obesity on severe disease and mortality in people with SARS-CoV-2: A systematic review and meta-analysis. Endocrinol. Diabetes Metab. 2020, 4, e00176. [Google Scholar] [CrossRef]
- WHO. Obesity and Overweight: Fact Sheet; WHO Media Centre: Geneva, Switzerland, 2020; Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 2 October 2020).
- Popkin, B.M.; Du, S.; Green, W.D.; Beck, M.A.; Algaith, T.; Herbst, C.H.; Alsukait, R.; Alluhidan, M.; Alazemi, N.; Shekar, M. Individuals with obesity and COVID-19: A global perspective on the epidemiology and biological relationships. Obes. Rev. 2020, 21, e13128. [Google Scholar] [CrossRef] [PubMed]
- Tartof, S.Y.; Qian, L.; Hong, M.V.; Wei, M.R.; Nadjafi, R.F.; Fischer, H.; Li, M.Z.; Shaw, D.S.F.; Caparosa, M.S.L.; Nau, C.L.; et al. Obesity and Mortality Among Patients Diagnosed With COVID-19: Results From an Integrated Health Care Organization. Ann. Intern. Med. 2020, 173, 773–781. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kompoti, M. Obesity and infection. Lancet Infect. Dis. 2006, 6, 438–446. [Google Scholar] [CrossRef]
- Kwong, J.C.; Campitelli, M.A.; Rosella, L.C. Obesity and Respiratory Hospitalizations During Influenza Seasons in Ontario, Canada: A Cohort Study. Clin. Infect. Dis. 2011, 53, 413–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC). Intensive-care patients with severe novel influenza A (H1N1) virus infection—Michigan, June 2009. MMWR Morb. Mortal. Wkly. Rep. 2009, 58, 749–752. [Google Scholar]
- Jung, C.-Y.; Park, H.; Kim, D.W.; Lim, H.; Chang, J.H.; Choi, Y.J.; Kim, S.W.; Chang, T.I. Association between Body Mass Index and Risk of COVID-19: A Nationwide Case-Control Study in South Korea. Clin. Infect. Dis. 2020, 73, e1855–e1862. [Google Scholar] [CrossRef]
- Ghoneim, S.; Butt, M.U.; Hamid, O.; Shah, A.; Asaad, I. The incidence of COVID-19 in patients with metabolic syndrome and non-alcoholic steatohepatitis: A population-based study. Metab. Open 2020, 8, 100057. [Google Scholar] [CrossRef]
- García, L.F. Immune Response, Inflammation, and the Clinical Spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef]
- Karlsson, E.A.; Sheridan, P.A.; Beck, M.A. Diet-Induced Obesity Impairs the T Cell Memory Response to Influenza Virus Infection. J. Immunol. 2010, 184, 3127–3133. [Google Scholar] [CrossRef] [Green Version]
- Neidich, S.D.; Green, W.D.; Rebeles, J.; Karlsson, E.; Schultz-Cherry, S.; Noah, T.L.; Chakladar, S.; Hudgens, M.G.; Weir, S.S.; Beck, M.A. Increased risk of influenza among vaccinated adults who are obese. Int. J. Obes. 2017, 41, 1324–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wadman, M. Why COVID-19 Is More Deadly in People with Obesity—Even if They’re Young. Science. 2020. Available online: https://www.sciencemag.org/news/2020/09/why-covid-19-more-deadly-people-obesity-even-if-theyre-young (accessed on 9 October 2020).
- Townsend, M.J.; Kyle, T.K.; Stanford, F.C. COVID-19 Vaccination and Obesity: Optimism and Challenges. Obesity 2021, 29, 634–635. [Google Scholar] [CrossRef]
- SARS-CoV-2-specific ELISA development. J. Immunol. Methods 2020, 484–485, 112832. [CrossRef]
- Nilles, E.J.; Karlson, E.W.; Norman, M.; Gilboa, T.; Fischinger, S.; Atyeo, C.; Zhou, G.; Bennett, C.L.; Tolan, N.V.; Oganezova, K.; et al. Evaluation of three commercial and two non-commercial immunoassays for the detection of prior infection to SARS-CoV-2. J. Appl. Lab. Med. 2021, 6, 1561–1570. [Google Scholar] [CrossRef]
- Atyeo, C.; Fischinger, S.; Zohar, T.; Slein, M.D.; Burke, J.; Loos, C.; McCulloch, D.J.; Newman, K.L.; Wolf, C.; Yu, J.; et al. Distinct Early Serological Signatures Track with SARS-CoV-2 Survival. Immunity 2020, 53, 524–532.e4. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.J.; Sheridan, P.A.; Karlsson, E.; Schultz-Cherry, S.; Shi, Q.; Beck, M.A. Diet-Induced Obese Mice Exhibit Altered Heterologous Immunity during a Secondary 2009 Pandemic H1N1 Infection. J. Immunol. 2013, 191, 2474–2485. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, P.A.; Paich, H.A.; Handy, J.; Karlsson, E.; Hudgens, M.G.; Sammon, A.B.; Holland, L.A.; Weir, S.; Noah, T.L.; Beck, M.A. Obesity is associated with impaired immune response to influenza vaccination in humans. Int. J. Obes. 2012, 36, 1072–1077. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, P.; Curtis, N. Factors That Influence the Immune Response to Vaccination. Clin. Microbiol. Rev. 2019, 32, e00084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becht, E.; McInnes, L.; Healy, J.; Dutertre, C.-A.; Kwok, I.W.H.; Ng, L.G.; Ginhoux, F.; Newell, E.W. Dimensionality reduction for visualizing single-cell data using UMAP. Nat. Biotechnol. 2019, 37, 38–44. [Google Scholar] [CrossRef]
- De Lusignan, S.; Dorward, J.; Correa, A.; Jones, N.; Akinyemi, O.; Amirthalingam, G.; Andrews, N.; Byford, R.; Dabrera, G.; Elliot, A.; et al. Risk factors for SARS-CoV-2 among patients in the Oxford Royal College of General Practitioners Research and Surveillance Centre primary care network: A cross-sectional study. Lancet Infect. Dis. 2020, 20, 1034–1042. [Google Scholar] [CrossRef]
- Gibbons, C.L.; Mangen, M.-J.J.; Plass, D.; Havelaar, A.H.; Brooke, R.J.; Kramarz, P.; Peterson, K.L.; Stuurman, A.L.; Cassini, A.; Fèvre, E.M.; et al. Measuring underreporting and under-ascertainment in infectious disease datasets: A comparison of methods. BMC Public Health 2014, 14, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipsitch, M.; Donnelly, C.; Fraser, C.; Blake, I.; Cori, A.; Dorigatti, I.; Ferguson, N.; Garske, T.; Mills, H.L.; Riley, S.; et al. Potential Biases in Estimating Absolute and Relative Case-Fatality Risks during Outbreaks. PLoS Negl. Trop. Dis. 2015, 9, e0003846. [Google Scholar] [CrossRef] [Green Version]
- Kass, D.A.; Duggal, P.; Cingolani, O. Obesity could shift severe COVID-19 disease to younger ages. Lancet 2020, 395, 1544–1545. [Google Scholar] [CrossRef]
- Moderbacher, C.R.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Painter, S.D.; Ovsyannikova, I.G.; Poland, G.A. The weight of obesity on the human immune response to vaccination. Vaccine 2015, 33, 4422–4429. [Google Scholar] [CrossRef] [Green Version]
- Alarcon, P.C.; Damen, M.S.; Madan, R.; Deepe, G.S.; Spearman, P.; Way, S.S.; Divanovic, S. Adipocyte inflammation and pathogenesis of viral pneumonias: An overlooked contribution. Mucosal Immunol. 2021, 14, 1224–1234. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Covariate 1 | All Participants (n = 4469) | Seropositive Participants (n = 322) | OR (95% CI) | p-Value 5 | |
|---|---|---|---|---|---|
| N | N | % | |||
| Age group | |||||
| 18–29 y | 1668 | 133 | 8.0% | ref | |
| 30–39 y | 1761 | 104 | 5.9% | 0.72 (0.56 to 0.94) | 0.0174 * |
| 40–49 y | 584 | 50 | 8.6% | 1.08 (0.77 to 1.52) | 0.6545 |
| 50–59 y | 315 | 26 | 8.3% | 1.04 (0.67 to 1.61) | 0.8666 |
| 60+ y | 85 | 2 | 2.4% | 0.28 (0.07 to 1.14) | 0.076 |
| BMI | |||||
| <18.5 | 34 | 3 | 8.8% | 1.44 (0.43 to 4.80) | 0.5500 |
| 18.5–<25 | 1686 | 106 | 6.3% | ref | |
| 25–<30 | 1523 | 101 | 6.6% | 1.06 (0.80 to 1.40) | 0.6916 |
| 30–<35 | 676 | 61 | 9.0% | 1.48 (1.06 to 2.05) | 0.0196 * |
| 35–<40 | 246 | 23 | 9.3% | 1.54 (0.96 to 2.47) | 0.0742 |
| ≥40 | 105 | 5 | 4.8% | 0.75 (0.30 to 1.87) | 0.5308 |
| Ethnicity | |||||
| Not Hispanic/Not Latinx | 2492 | 113 | 4.5% | ref | |
| Hispanic/Latinx | 1274 | 155 | 12.2% | 2.91 (2.26 to 3.75) | <0.0001 **** |
| Race | |||||
| White | 2862 | 185 | 6.5% | ref | |
| American Indian/Alaska Native | 32 | 3 | 9.4% | 1.50 (0.45 to 4.96) | 0.5092 |
| Asian | 442 | 18 | 4.1% | 0.61 (0.37 to 1.01) | 0.0535 |
| Black | 72 | 2 | 2.8% | 0.41 (0.10 to 1.70) | 0.2207 |
| Native Hawaiian/Pacific Islander | 29 | 2 | 6.9% | 1.07 (0.25 to 4.54) | 0.9249 |
| More than one race | 292 | 13 | 4.5% | 0.67 (0.38 to 1.20) | 0.1796 |
| Sex 2 | |||||
| Female | 600 | 40 | 6.7% | ref | |
| Male | 3730 | 267 | 7.2% | 1.08 (0.77 to 1.52) | 0.6634 |
| Children ≤ 18 y in household | |||||
| No | 3014 | 204 | 6.8% | ref | |
| Yes | 1342 | 106 | 7.9% | 1.18 (0.93 to 1.51) | 0.1808 |
| No. in household | |||||
| 1 | 640 | 41 | 6.4% | ref | |
| 2–4 | 3027 | 214 | 7.1% | 1.11 (0.79 to 1.57) | 0.5490 |
| >4 | 659 | 51 | 7.7% | 1.23 (0.80 to 1.88) | 0.3499 |
| Primary work location | |||||
| Cape Canaveral, Florida | 268 | 17 | 6.3% | ref | |
| Hawthorne, California | 2859 | 111 | 3.9% | 0.60 (0.35 to 1.01) | 0.0544 |
| McGregor, Texas | 257 | 21 | 8.2% | 1.31 (0.68 to 2.55) | 0.4202 |
| Seattle, Washington | 253 | 5 | 2.0% | 0.30 (0.11 to 0.82) | 0.0190 * |
| South Texas, Texas | 712 | 160 | 22.5% | 4.28 (2.54 to 7.21) | <0.0001 **** |
| Other | 69 | 1 | 1.4% | 0.23 (0.03 to 1.79) | 0.1623 |
| Comorbidities 3,4 | |||||
| Asthma | 368 | 20 | 5.4% | 0.72 (0.45 to 1.15) | 0.1721 |
| Hypertension | 356 | 26 | 7.3% | 1.02 (0.67 to 1.54) | 0.9405 |
| Diabetes mellitus | 101 | 11 | 10.9% | 1.59 (0.84 to 3.01) | 0.1509 |
| Coronary heart disease | 17 | 1 | 5.9% | 0.80 (0.11 to 6.08) | 0.8329 |
| Stroke | 9 | 2 | 22.2% | 3.70 (0.76 to 17.87) | 0.1039 |
| Emphysema/COPD | 9 | 1 | 11.1% | 1.61 (0.20 to 12.93) | 0.6532 |
| Cancer—not receiving treatment | 39 | 2 | 5.1% | 0.69 (0.17 to 2.89) | 0.6163 |
| Other lung disease | 26 | 2 | 7.7% | 1.07 (0.25 to 4.56) | 0.9233 |
| Other immunocompromised | 61 | 4 | 6.6% | 0.92 (0.33 to 2.55) | 0.8710 |
| Other chronic medical condition | 176 | 9 | 5.1% | 0.72 (0.36 to 1.43) | 0.3471 |
| Smoking history | |||||
| Never | 3769 | 263 | 7.0% | ref | |
| Prior | 367 | 24 | 6.5% | 0.93 (0.61 to 1.44) | 0.7514 |
| Current | 229 | 23 | 10.0% | 1.49 (0.95 to 2.33) | 0.0826 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nilles, E.J.; Siddiqui, S.M.; Fischinger, S.; Bartsch, Y.C.; de St. Aubin, M.; Zhou, G.; Gluck, M.J.; Berger, S.; Rhee, J.; Petersen, E.; et al. Epidemiological and Immunological Features of Obesity and SARS-CoV-2. Viruses 2021, 13, 2235. https://doi.org/10.3390/v13112235
Nilles EJ, Siddiqui SM, Fischinger S, Bartsch YC, de St. Aubin M, Zhou G, Gluck MJ, Berger S, Rhee J, Petersen E, et al. Epidemiological and Immunological Features of Obesity and SARS-CoV-2. Viruses. 2021; 13(11):2235. https://doi.org/10.3390/v13112235
Chicago/Turabian StyleNilles, Eric J., Sameed M. Siddiqui, Stephanie Fischinger, Yannic C. Bartsch, Michael de St. Aubin, Guohai Zhou, Matthew J. Gluck, Samuel Berger, Justin Rhee, Eric Petersen, and et al. 2021. "Epidemiological and Immunological Features of Obesity and SARS-CoV-2" Viruses 13, no. 11: 2235. https://doi.org/10.3390/v13112235