
Inhibitors of Activin Receptor-like Kinase 5 Interfere with SARS-CoV-2 S-Protein Processing and Spike-Mediated Cell Fusion via Attenuation of Furin Expression

 , , , , and
, , , , and 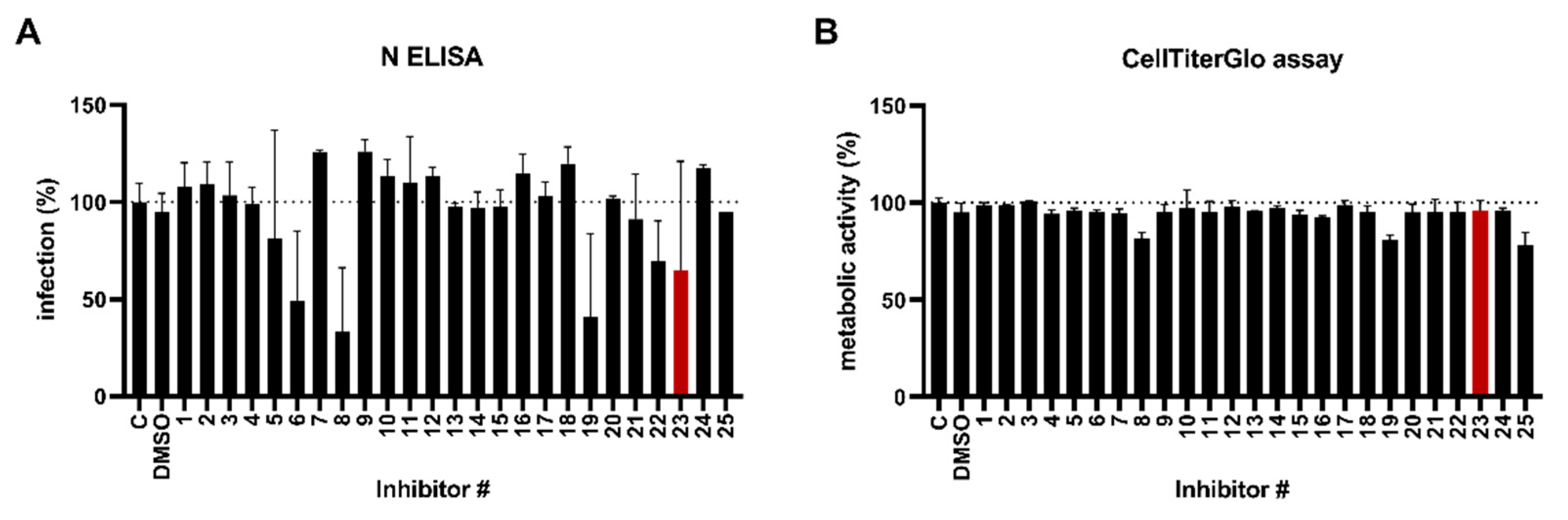{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Substances
2.3. Plasmids
- D614G: pCG1_SARS-2-Sdel18_D614G (D614G);
- Alpha: pCG1_SARS-2-Sdel18_DG614G + VOC-202012_01 (UK variant);
- Beta: pCG1_SARS-2-Sdel18_DG614G + SA_501-V2new (South Africa variant);
- Gamma: pCG1_SARS-2-Sdel18_DG614G + Brasil;
- Kappa: pCG1_SARS-2-Sdel18_D614G (IND) B.1.617.1 (kappa);
- Omicron BA.1: pCG1_SARS-2-Sdel18 (B.1.1.529/Omicron BA.1).
2.4. Nucleocapsid Protein In-Cell ELISA
2.5. CellTiter-Glo Luminescent Cell Viability Assay
2.6. Transfection
2.7. Immunoblotting
2.8. Cell–Cell Fusion Assay
3. Results
3.1. Identification of an ALK5 Kinase Inhibitor That Supresses SARS-CoV-2 Infection
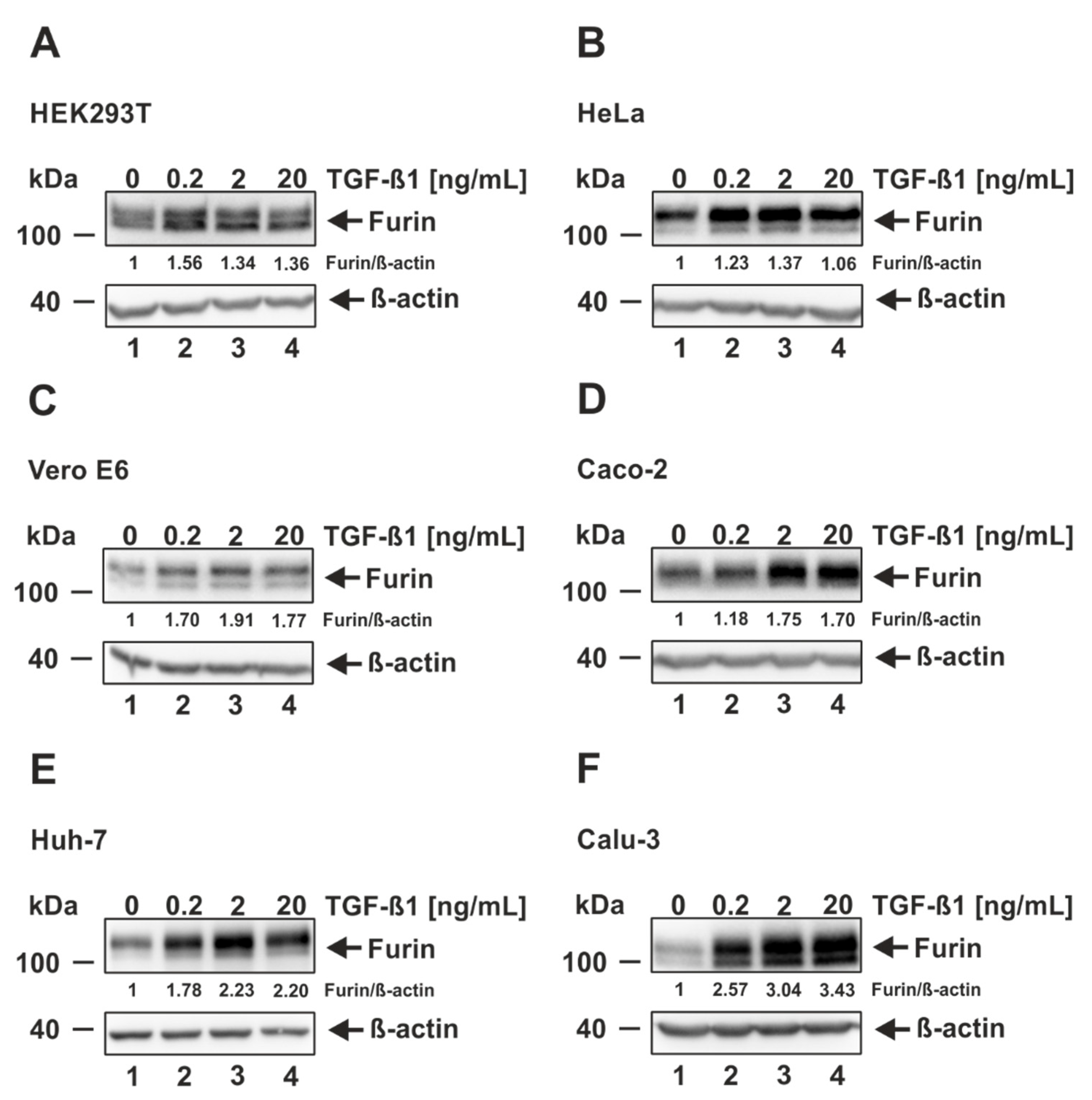
3.2. TGF-ß1 Induces Furin Expression in Cells Permissive and Non-Permissive for SARS-CoV-2 Infection
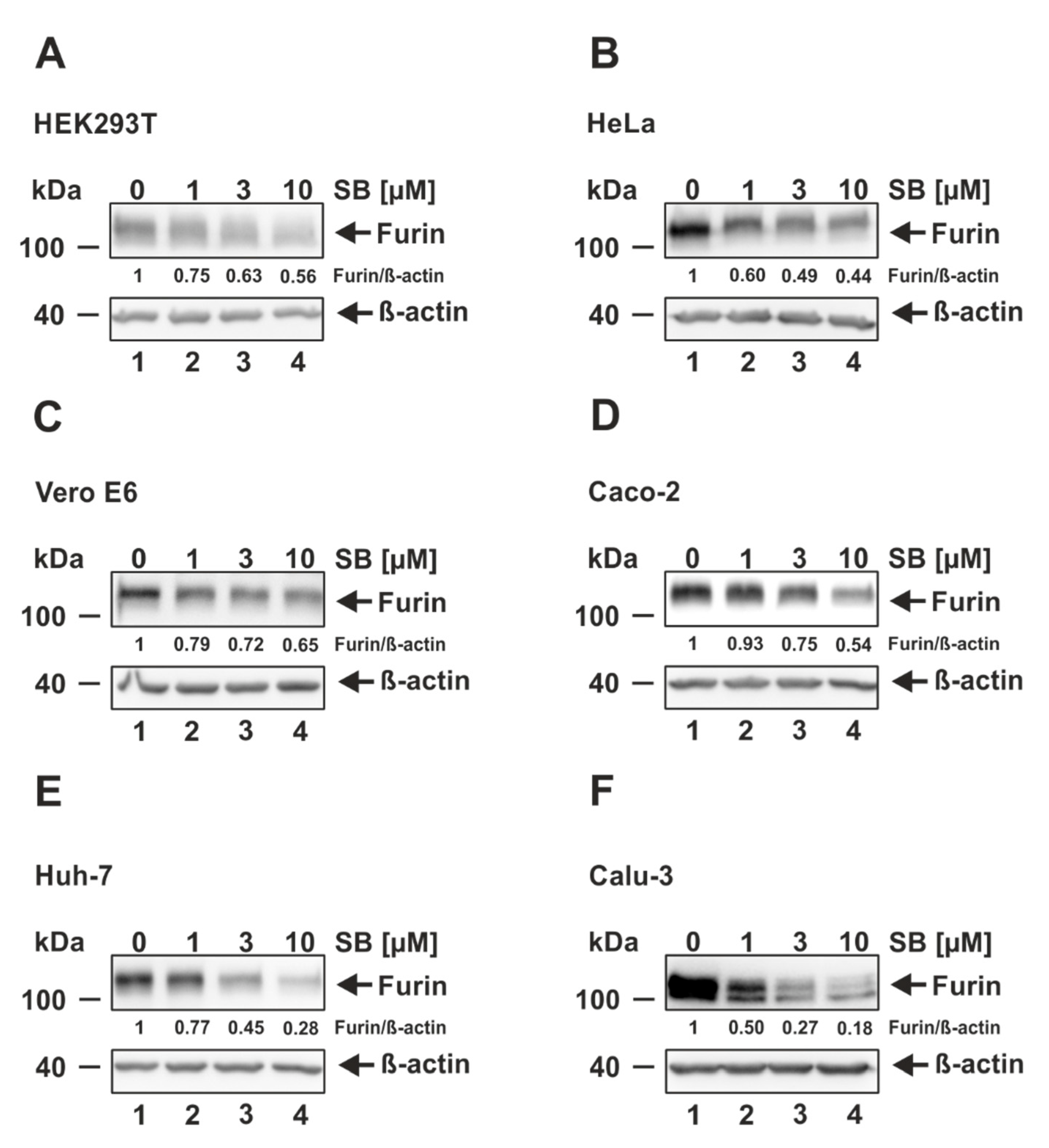
3.3. ALK5 Inhibition Decreases Furin Expression in Cells Permissive and Non-Permissive for SARS-CoV-2 Infection
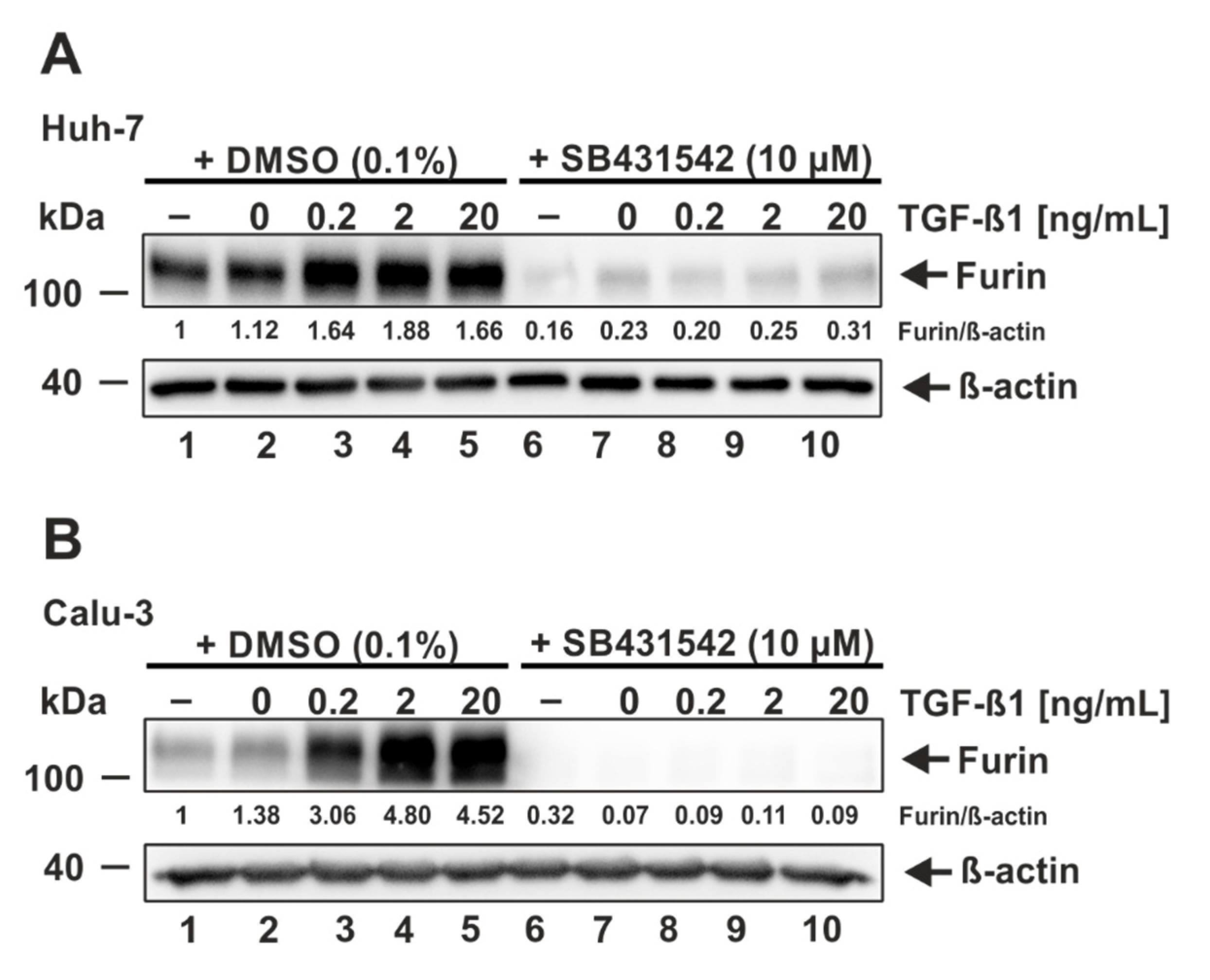
3.4. ALK5 Inhibition Prevents TGF-ß1-Induced Furin Expression
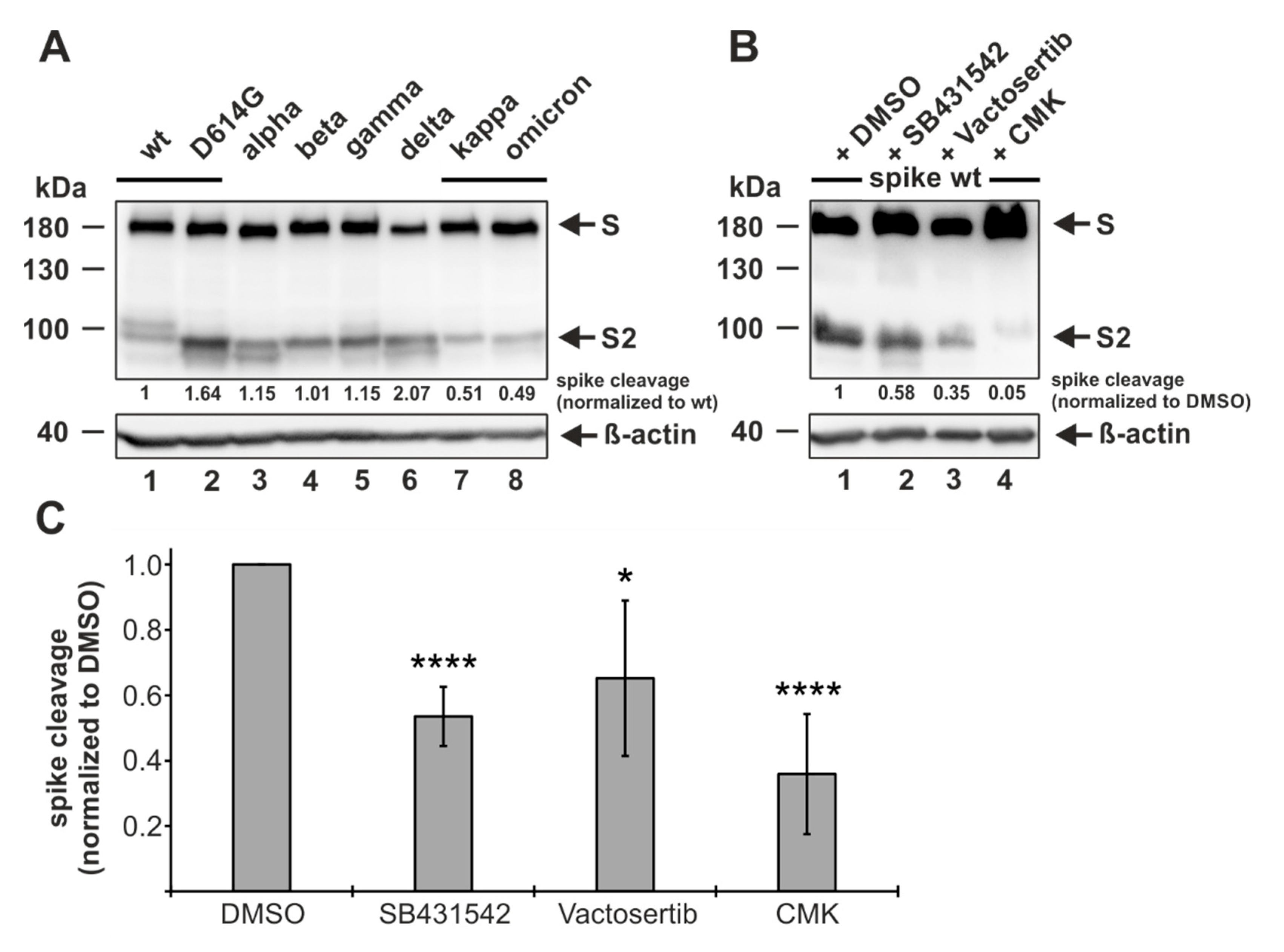
3.5. ALK5 Inhibition Reduces Proteolytic Processing of Spike
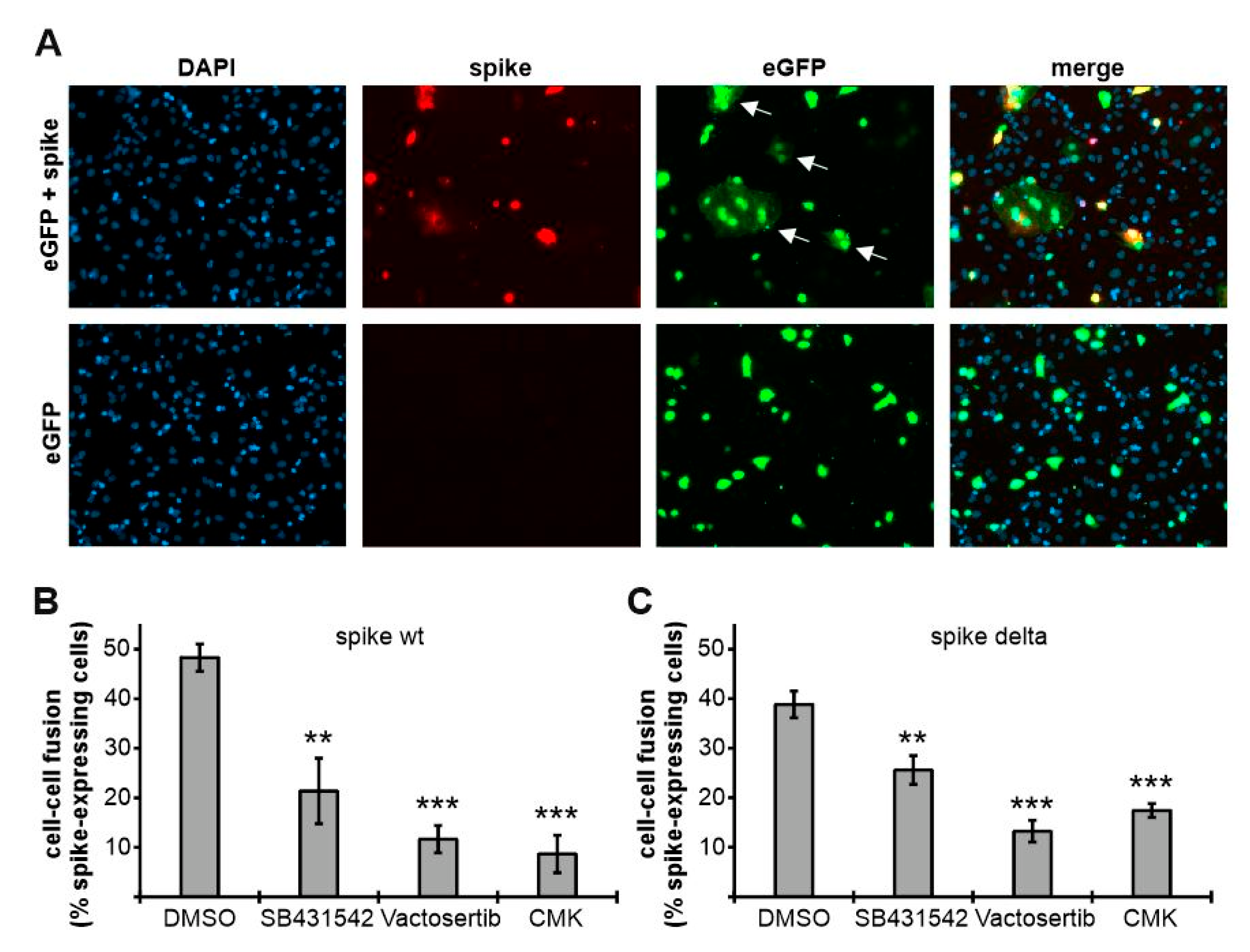
3.6. ALK5 Inhibition Attenuates Spike-Mediated Cell–Cell Fusion
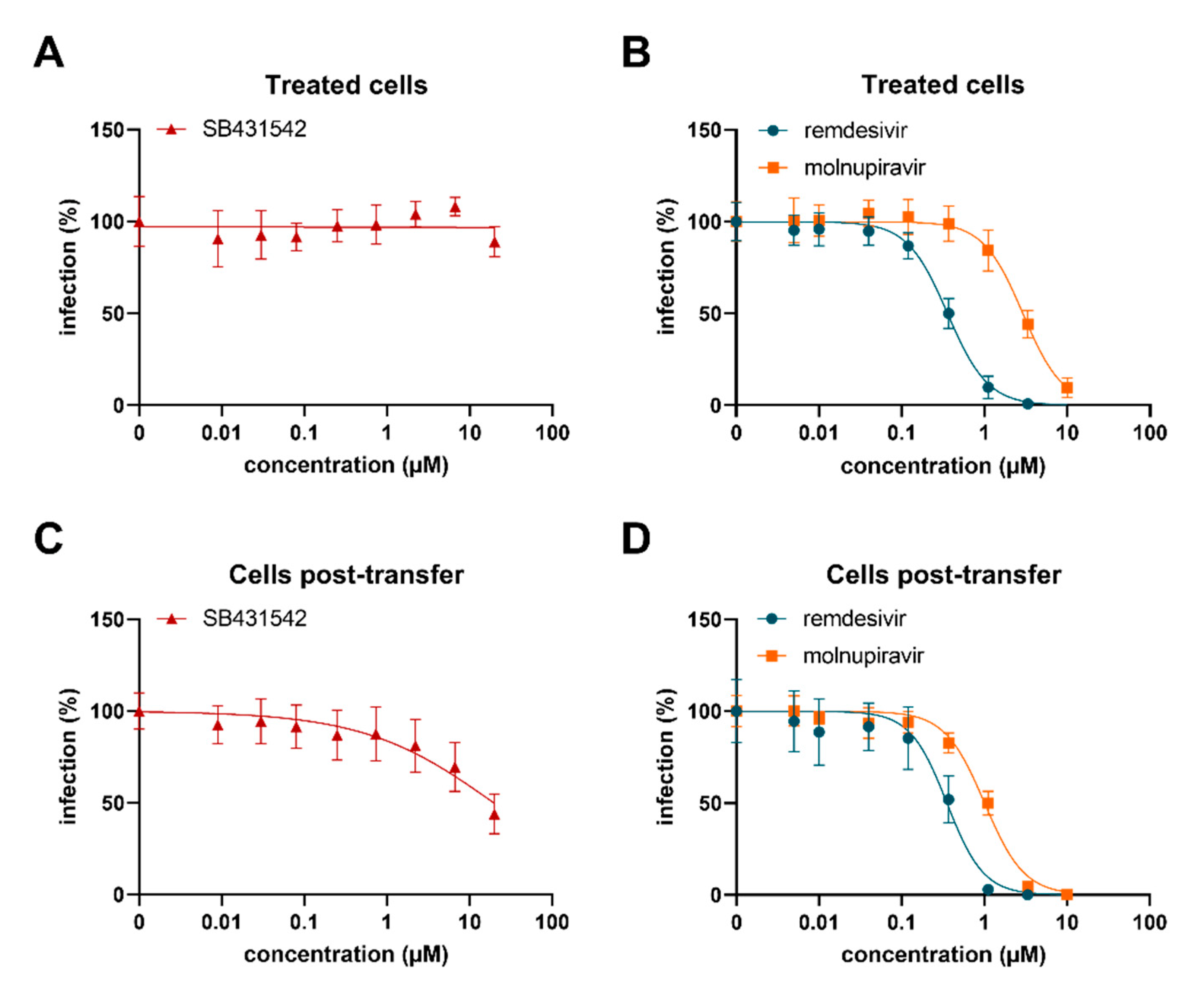
3.7. SB431542 Reduces Infectious SARS-CoV-2 Production
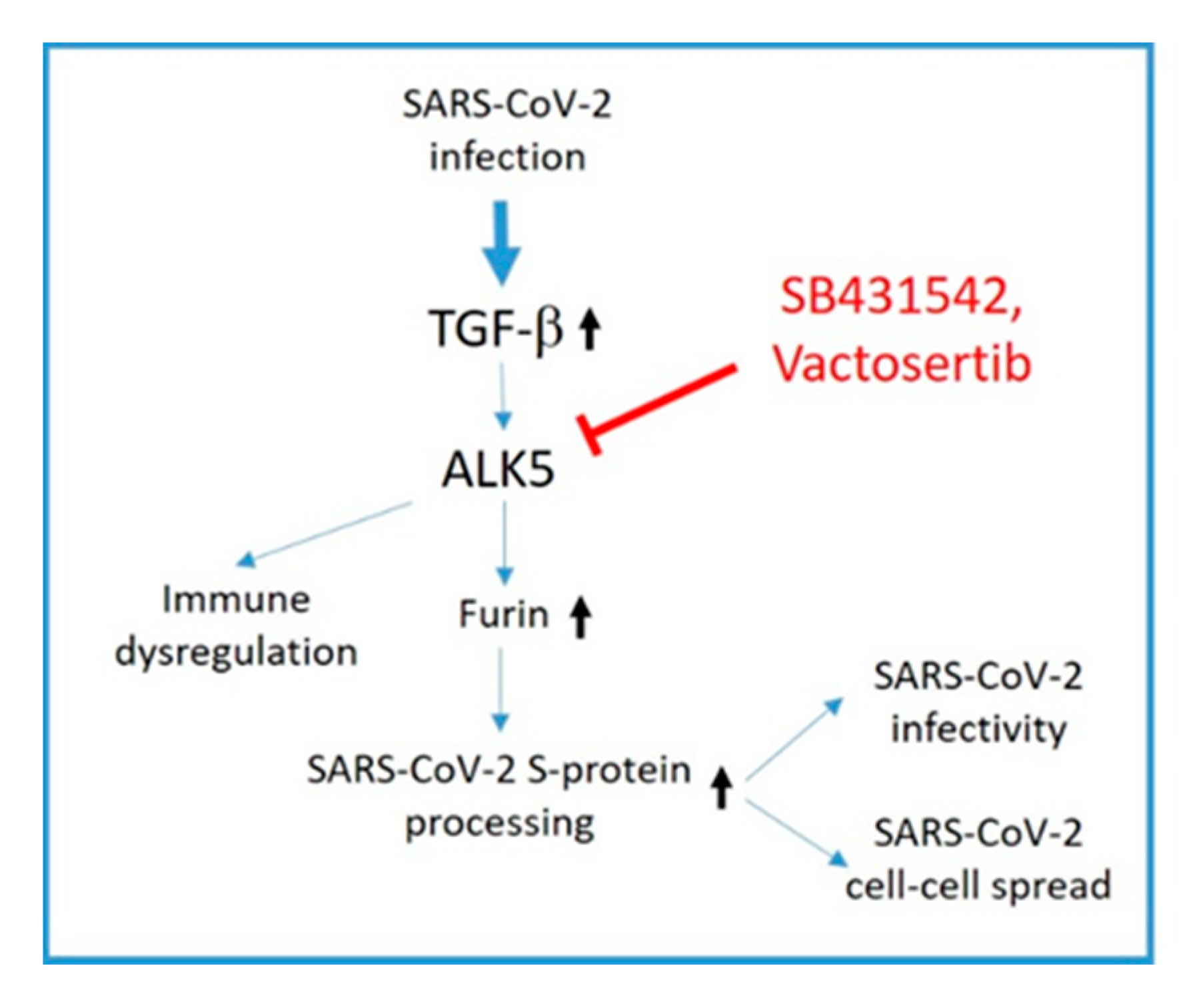
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, P.; Yang, X.L. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Zhao, S. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Sharma, P. Structural and functional insights into the spike protein mutations of emerging SARS-CoV-2 variants. Cell. Mol. Life Sci. CMLS 2021, 78, 7967–7989. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Potdar, V. SARS-CoV-2 Spike Mutations, L452R, T478K, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. Microorganisms 2021, 9, 1542. [Google Scholar] [CrossRef]
- Bestle, D.; Heindl, M.R. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci. Alliance 2020, 3, e202000786. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H. A Multibasic Cleavage Site in the Spike Protein of SARS-CoV-2 Is Essential for Infection of Human Lung Cells. Mol. Cell 2020, 78, 779–784. [Google Scholar] [CrossRef]
- Takeda, M. Proteolytic activation of SARS-CoV-2 spike protein. Microbiol. Immunol. 2022, 66, 15–23. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M. Mechanisms of SARS-CoV-2 entry into cells. Nat. Reviews. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. Methods Mol. Biol. 2015, 1282, 1–23. [Google Scholar]
- Buchrieser, J.; Dufloo, J. Syncytia formation by SARS-CoV-2-infected cells. EMBO J. 2020, 39, e106267. [Google Scholar] [CrossRef]
- Zeng, C.; Evans, J.P. SARS-CoV-2 spreads through cell-to-cell transmission. Proc. Natl. Acad. Sci. USA 2022, 119, e2111400119. [Google Scholar] [CrossRef]
- Bussani, R.; Schneider, E. Persistence of viral RNA, pneumocyte syncytia and thrombosis are hallmarks of advanced COVID-19 pathology. EBioMedicine 2020, 61, 103104. [Google Scholar] [CrossRef]
- Tian, S.; Hu, W. Pulmonary Pathology of Early-Phase 2019 Novel Coronavirus (COVID-19) Pneumonia in Two Patients with Lung Cancer. J. Thorac. Oncol. Off. Publ. Int. Assoc. Study Lung Cancer 2020, 15, 700–704. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, L. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Rajah, M.M.; Hubert, M. SARS-CoV-2 Alpha, Beta, and Delta variants display enhanced Spike-mediated syncytia formation. EMBO J. 2021, 40, e108944. [Google Scholar] [CrossRef]
- Cheng, Y.W.; Chao, T.L. Furin Inhibitors Block SARS-CoV-2 Spike Protein Cleavage to Suppress Virus Production and Cytopathic Effects. Cell Rep. 2020, 33, 108254. [Google Scholar] [CrossRef]
- Xia, S.; Lan, Q. The role of furin cleavage site in SARS-CoV-2 spike protein-mediated membrane fusion in the presence or absence of trypsin. Signal Transduct. Target. Ther. 2020, 5, 92. [Google Scholar] [CrossRef]
- Johnson, B.A.; Xie, X. Loss of furin cleavage site attenuates SARS-CoV-2 pathogenesis. Nature 2021, 591, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.Y.; Wang, P. Attenuated SARS-CoV-2 variants with deletions at the S1/S2 junction. Emerg. Microbes Infect. 2020, 9, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Goldhill, D.H. The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat. Microbiol. 2021, 6, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, M.; Toba, S. SARS-CoV-2 Bearing a Mutation at the S1/S2 Cleavage Site Exhibits Attenuated Virulence and Confers Protective Immunity. mBio 2021, 12, e0141521. [Google Scholar] [CrossRef]
- Xing, Y.; Li, X. Natural Polymorphisms Are Present in the Furin Cleavage Site of the SARS-CoV-2 Spike Glycoprotein. Front. Genet. 2020, 11, 783. [Google Scholar] [CrossRef]
- Chen, A.T.; Altschuler, K. COVID-19 CG enables SARS-CoV-2 mutation and lineage tracking by locations and dates of interest. eLife 2021, 10, e63409. [Google Scholar] [CrossRef]
- Chan, Y.A.; Zhan, S.H. The emergence of the spike furin cleavage site in SARS-CoV-2. Mol. Biol. Evol. 2021, 39, msab327. [Google Scholar] [CrossRef]
- Wu, C.; Zheng, M. Furin: A Potential Therapeutic Target for COVID-19. iScience 2020, 23, 101642. [Google Scholar] [CrossRef]
- Rahbar Saadat, Y.; Hosseiniyan Khatibi, S.M. Host Serine Proteases: A Potential Targeted Therapy for COVID-19 and Influenza. Front. Mol. Biosci. 2021, 8, 725528. [Google Scholar] [CrossRef]
- Blanchette, F.; Day, R. TGFbeta1 regulates gene expression of its own converting enzyme furin. J. Clin. Investig. 1997, 99, 1974–1983. [Google Scholar] [CrossRef] [Green Version]
- Blanchette, F.; Rivard, N. Cross-talk between the p42/p44 MAP kinase and Smad pathways in transforming growth factor beta 1-induced furin gene transactivation. J. Biol. Chem. 2001, 276, 33986–33994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, F.; Rudd, P. Involvement of Smads in TGFbeta1-induced furin (fur) transcription. J. Cell. Physiol. 2001, 188, 264–273. [Google Scholar] [CrossRef]
- O’Sullivan, M.J.; Mitchel, J.A. In well-differentiated primary human bronchial epithelial cells, TGF-β1 and TGF-β2 induce expression of furin. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 320, L246–L253. [Google Scholar] [CrossRef]
- Weiss, A.; Attisano, L. The TGFbeta superfamily signaling pathway. Wiley Interdiscip. Reviews. Dev. Biol. 2013, 2, 47–63. [Google Scholar] [CrossRef]
- Franzén, P.; ten Dijke, P. Cloning of a TGF beta type I receptor that forms a heteromeric complex with the TGF beta type II receptor. Cell 1993, 75, 681–692. [Google Scholar] [CrossRef]
- Baumann, J.; Huang, S.F. Furin inhibition prevents hypoxic and TGFβ-mediated blood-brain barrier disruption. Exp. Cell Res. 2019, 383, 111503. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.E.; Lee, W.C. Tgf-beta type I receptor (Alk5) kinase inhibitors in oncology. Curr. Pharm. Biotechnol. 2011, 12, 2190–2202. [Google Scholar] [CrossRef]
- Weisberg, E.; Parent, A. Repurposing of Kinase Inhibitors for Treatment of COVID-19. Pharm. Res. 2020, 37, 167. [Google Scholar] [CrossRef]
- Pillaiyar, T.; Laufer, S. Kinases as Potential Therapeutic Targets for Anti-coronaviral Therapy. J. Med. Chem. 2021, 65, 955–982. [Google Scholar] [CrossRef]
- Shen, W.X.; Luo, R.C. Features of Cytokine Storm Identified by Distinguishing Clinical Manifestations in COVID-19. Front. Public Health 2021, 9, 671788. [Google Scholar] [CrossRef]
- Ferreira-Gomes, M.; Kruglov, A. SARS-CoV-2 in severe COVID-19 induces a TGF-β-dominated chronic immune response that does not target itself. Nat. Commun. 2021, 12, 1961. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Xu, X. SARS-CoV-2 induces transcriptional signatures in human lung epithelial cells that promote lung fibrosis. Respir. Res. 2020, 21, 182. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, Y. Transcriptomic characteristics of bronchoalveolar lavage fluid and peripheral blood mononuclear cells in COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Amanat, F.; Stadlbauer, D. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat. Med. 2020, 26, 1033–1036. [Google Scholar] [CrossRef]
- Hoffmann, M.; Arora, P. SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393. [Google Scholar] [CrossRef]
- Hoffmann, M.; Hofmann-Winkler, H. SARS-CoV-2 variant B.1.617 is resistant to bamlanivimab and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 36, 109415. [Google Scholar] [CrossRef]
- Hoffmann, M.; Krüger, N. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456. [Google Scholar] [CrossRef]
- Groß, R.; Zanoni, M. Heterologous ChAdOx1 nCoV-19 and BNT162b2 prime-boost vaccination elicits potent neutralizing antibody responses and T cell reactivity against prevalent SARS-CoV-2 variants. EBioMedicine 2022, 75, 103761. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, T. Ultrapotent antibodies against diverse and highly transmissible SARS-CoV-2 variants. Science 2021, 373, eabh1766. [Google Scholar] [CrossRef]
- Conzelmann, C.; Gilg, A. An enzyme-based immunodetection assay to quantify SARS-CoV-2 infection. Antivir. Res. 2020, 181, 104882. [Google Scholar] [CrossRef]
- Klann, K.; Bojkova, D. Growth Factor Receptor Signaling Inhibition Prevents SARS-CoV-2 Replication. Mol. Cell 2020, 80, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Rudd, C.E. GSK-3 Inhibition as a Therapeutic Approach Against SARs CoV2: Dual Benefit of Inhibiting Viral Replication While Potentiating the Immune Response. Front. Immunol. 2020, 11, 1638. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Verma, A. Targeting the coronavirus nucleocapsid protein through GSK-3 inhibition. Proc. Natl. Acad. Sci. USA 2021, 118, e2113401118. [Google Scholar] [CrossRef] [PubMed]
- Appelberg, S.; Gupta, S. Dysregulation in Akt/mTOR/HIF-1 signaling identified by proteo-transcriptomics of SARS-CoV-2 infected cells. Emerg. Microbes Infect. 2020, 9, 1748–1760. [Google Scholar] [CrossRef]
- Garcia, G., Jr.; Sharma, A. Antiviral drug screen identifies DNA-damage response inhibitor as potent blocker of SARS-CoV-2 replication. Cell Rep. 2021, 35, 108940. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, Q. 5-Iodotubercidin inhibits SARS-CoV-2 RNA synthesis. Antivir. Res. 2022, 198, 105254. [Google Scholar] [CrossRef]
- Chen, F.; Shi, Q. A systems-level study reveals host-targeted repurposable drugs against SARS-CoV-2 infection. Mol. Syst. Biol. 2021, 17, e10239. [Google Scholar] [CrossRef]
- Adhami, M.; Sadeghi, B. Repurposing novel therapeutic candidate drugs for coronavirus disease-19 based on protein-protein interaction network analysis. BMC Biotechnol. 2021, 21, 22. [Google Scholar] [CrossRef]
- Inman, G.J.; Nicolás, F.J. SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2002, 62, 65–74. [Google Scholar] [CrossRef]
- Papa, G.; Mallery, D.L. Furin cleavage of SARS-CoV-2 Spike promotes but is not essential for infection and cell-cell fusion. PLoS Pathog. 2021, 17, e1009246. [Google Scholar] [CrossRef]
- Hörnich, B.F.; Großkopf, A.K. SARS-CoV-2 and SARS-CoV Spike-Mediated Cell-Cell Fusion Differ in Their Requirements for Receptor Expression and Proteolytic Activation. J. Virol. 2021, 95, e00002-21. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J. Delta spike P681R mutation enhances SARS-CoV-2 fitness over Alpha variant. Biorxiv Prepr. Serv. Biol. 2021, 39, 110829. [Google Scholar] [CrossRef]
- Neerukonda, S.N.; Vassell, R. SARS-CoV-2 Delta Variant Displays Moderate Resistance to Neutralizing Antibodies and Spike Protein Properties of Higher Soluble ACE2 Sensitivity, Enhanced Cleavage and Fusogenic Activity. Viruses 2021, 13, 2485. [Google Scholar] [CrossRef]
- Meng, B.; Abdullahi, A. Altered TMPRSS2 usage by SARS-CoV-2 Omicron impacts tropism and fusogenicity. Nature 2022, 603, 706–714. [Google Scholar] [CrossRef]
- Xia, S.; Liu, M. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.D.; Parveen, A. SARS-CoV-2: Emergence of New Variants and Effectiveness of Vaccines. Front. Cell. Infect. Microbiol. 2021, 11, 777212. [Google Scholar] [CrossRef]
- Ng, T.I.; Correia, I. Antiviral Drug Discovery for the Treatment of COVID-19 Infections. Viruses 2022, 14, 961. [Google Scholar] [CrossRef]
- Callahan, J.F.; Burgess, J.L. Identification of novel inhibitors of the transforming growth factor beta1 (TGF-beta1) type 1 receptor (ALK5). J. Med. Chem. 2002, 45, 999–1001. [Google Scholar] [CrossRef]
- Heldin, C.H.; Moustakas, A. Signaling Receptors for TGF-β Family Members. Cold Spring Harb. Perspect. Biol. 2016, 8, a022053. [Google Scholar] [CrossRef] [Green Version]
- Tzavlaki, K.; Moustakas, A. TGF-β Signaling. Biomolecules 2020, 10, 487. [Google Scholar] [CrossRef] [Green Version]
- Hojyo, S.; Uchida, M. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef]
- Oronsky, B.; Larson, C. A Review of Persistent Post-COVID Syndrome (PPCS). Clin. Rev. Allergy Immunol. 2021, 20, 1–9. [Google Scholar] [CrossRef]
- Chen, W. A potential treatment of COVID-19 with TGF-β blockade. Int. J. Biol. Sci. 2020, 16, 1954–1955. [Google Scholar] [CrossRef]
- Hao, S.; Ning, K. Long-Term Modeling of SARS-CoV-2 Infection of In Vitro Cultured Polarized Human Airway Epithelium. mBio 2020, 11, e02852-20. [Google Scholar] [CrossRef]
- Laprise, M.H.; Grondin, F. Furin gene (fur) regulation in differentiating human megakaryoblastic Dami cells: Involvement of the proximal GATA recognition motif in the P1 promoter and impact on the maturation of furin substrates. Blood 2002, 100, 3578–3587. [Google Scholar] [CrossRef] [Green Version]
- McMahon, S.; Grondin, F. Hypoxia-enhanced expression of the proprotein convertase furin is mediated by hypoxia-inducible factor-1: Impact on the bioactivation of proproteins. J. Biol. Chem. 2005, 280, 6561–6569. [Google Scholar] [CrossRef] [Green Version]
- Ayoubi, T.A.; Creemers, J.W. Expression of the dibasic proprotein processing enzyme furin is directed by multiple promoters. J. Biol. Chem. 1994, 269, 9298–9303. [Google Scholar] [CrossRef]
- Ghazavi, A.; Ganji, A. Cytokine profile and disease severity in patients with COVID-19. Cytokine 2021, 137, 155323. [Google Scholar] [CrossRef] [PubMed]
- Witkowski, M.; Tizian, C. Untimely TGFβ responses in COVID-19 limit antiviral functions of NK cells. Nature 2021, 600, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Shuai, H.; Chan, J.F. Attenuated replication and pathogenicity of SARS-CoV-2 B.1.1.529 Omicron. Nature 2022, 603, 693–699. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lu, L. SARS-CoV-2 Omicron variant shows less efficient replication and fusion activity when compared with Delta variant in TMPRSS2-expressed cells. Emerg. Microbes Infect. 2022, 11, 277–283. [Google Scholar] [CrossRef]
- Arora, P.; Sidarovich, A. B.1.617.2 enters and fuses lung cells with increased efficiency and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 37, 109825. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, T. Membrane fusion and immune evasion by the spike protein of SARS-CoV-2 Delta variant. Science 2021, 374, 1353–1360. [Google Scholar] [CrossRef]
- Saito, A.; Irie, T. Enhanced fusogenicity and pathogenicity of SARS-CoV-2 Delta P681R mutation. Nature 2022, 602, 300–306. [Google Scholar] [CrossRef]
- Quarleri, J.; Galvan, V. Omicron variant of the SARS-CoV-2: A quest to define the consequences of its high mutational load. GeroScience 2022, 44, 53–56. [Google Scholar] [CrossRef]
- Rajah, M.M.; Bernier, A. The Mechanism and Consequences of SARS-CoV-2 Spike-Mediated Fusion and Syncytia Formation. J. Mol. Biol. 2021, 434, 167280. [Google Scholar] [CrossRef]
- Jaimes, J.A.; Millet, J.K. Proteolytic Cleavage of the SARS-CoV-2 Spike Protein and the Role of the Novel S1/S2 Site. iScience 2020, 23, 101212. [Google Scholar] [CrossRef]
- Tang, T.; Jaimes, J.A. Proteolytic Activation of SARS-CoV-2 Spike at the S1/S2 Boundary: Potential Role of Proteases beyond Furin. ACS Infect. Dis. 2021, 7, 264–272. [Google Scholar] [CrossRef]
- Dubois, C.M.; Blanchette, F. Evidence that furin is an authentic transforming growth factor-beta1-converting enzyme. Am. J. Pathol. 2001, 158, 305–316. [Google Scholar] [CrossRef]
- Lehmann, M.; André, F. Deficient processing and activity of type I insulin-like growth factor receptor in the furin-deficient LoVo-C5 cells. Endocrinology 1998, 139, 3763–3771. [Google Scholar] [CrossRef]
- Khatib, A.M.; Siegfried, G. Inhibition of proprotein convertases is associated with loss of growth and tumorigenicity of HT-29 human colon carcinoma cells: Importance of insulin-like growth factor-1 (IGF-1) receptor processing in IGF-1-mediated functions. J. Biol. Chem. 2001, 276, 30686–30693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winn, B.J. Is there a role for insulin-like growth factor inhibition in the treatment of COVID-19-related adult respiratory distress syndrome? Med. Hypotheses 2020, 144, 110167. [Google Scholar] [CrossRef] [PubMed]
- Rodón, J.; Carducci, M. Pharmacokinetic, pharmacodynamic and biomarker evaluation of transforming growth factor-β receptor I kinase inhibitor, galunisertib, in phase 1 study in patients with advanced cancer. Investig. New Drugs 2015, 33, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, T.A.; Vieito, M. First-In-Human Phase I Study of a Next-Generation, Oral, TGFβ Receptor 1 Inhibitor, LY3200882, in Patients with Advanced Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2021, 27, 6666–6676. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Nokihara, H. Phase 1 study of galunisertib, a TGF-beta receptor I kinase inhibitor, in Japanese patients with advanced solid tumors. Cancer Chemother. Pharmacol. 2015, 76, 1143–1152. [Google Scholar] [CrossRef]
- Jung, S.Y.; Hwang, S. Pharmacokinetic characteristics of vactosertib, a new activin receptor-like kinase 5 inhibitor, in patients with advanced solid tumors in a first-in-human phase 1 study. Investig. New Drugs 2020, 38, 812–820. [Google Scholar] [CrossRef]
- Anderton, M.J.; Mellor, H.R. Induction of heart valve lesions by small-molecule ALK5 inhibitors. Toxicol. Pathol. 2011, 39, 916–924. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezger, M.C.; Conzelmann, C.; Weil, T.; von Maltitz, P.; Albers, D.P.J.; Münch, J.; Stamminger, T.; Schilling, E.-M. Inhibitors of Activin Receptor-like Kinase 5 Interfere with SARS-CoV-2 S-Protein Processing and Spike-Mediated Cell Fusion via Attenuation of Furin Expression. Viruses 2022, 14, 1308. https://doi.org/10.3390/v14061308
Mezger MC, Conzelmann C, Weil T, von Maltitz P, Albers DPJ, Münch J, Stamminger T, Schilling E-M. Inhibitors of Activin Receptor-like Kinase 5 Interfere with SARS-CoV-2 S-Protein Processing and Spike-Mediated Cell Fusion via Attenuation of Furin Expression. Viruses. 2022; 14(6):1308. https://doi.org/10.3390/v14061308
Chicago/Turabian StyleMezger, Maja C., Carina Conzelmann, Tatjana Weil, Pascal von Maltitz, Dan P. J. Albers, Jan Münch, Thomas Stamminger, and Eva-Maria Schilling. 2022. "Inhibitors of Activin Receptor-like Kinase 5 Interfere with SARS-CoV-2 S-Protein Processing and Spike-Mediated Cell Fusion via Attenuation of Furin Expression" Viruses 14, no. 6: 1308. https://doi.org/10.3390/v14061308