
Duplex One-Step RT-qPCR Assays for Simultaneous Detection of Genomic and Subgenomic RNAs of SARS-CoV-2 Variants

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses
2.3. RNA Extractions
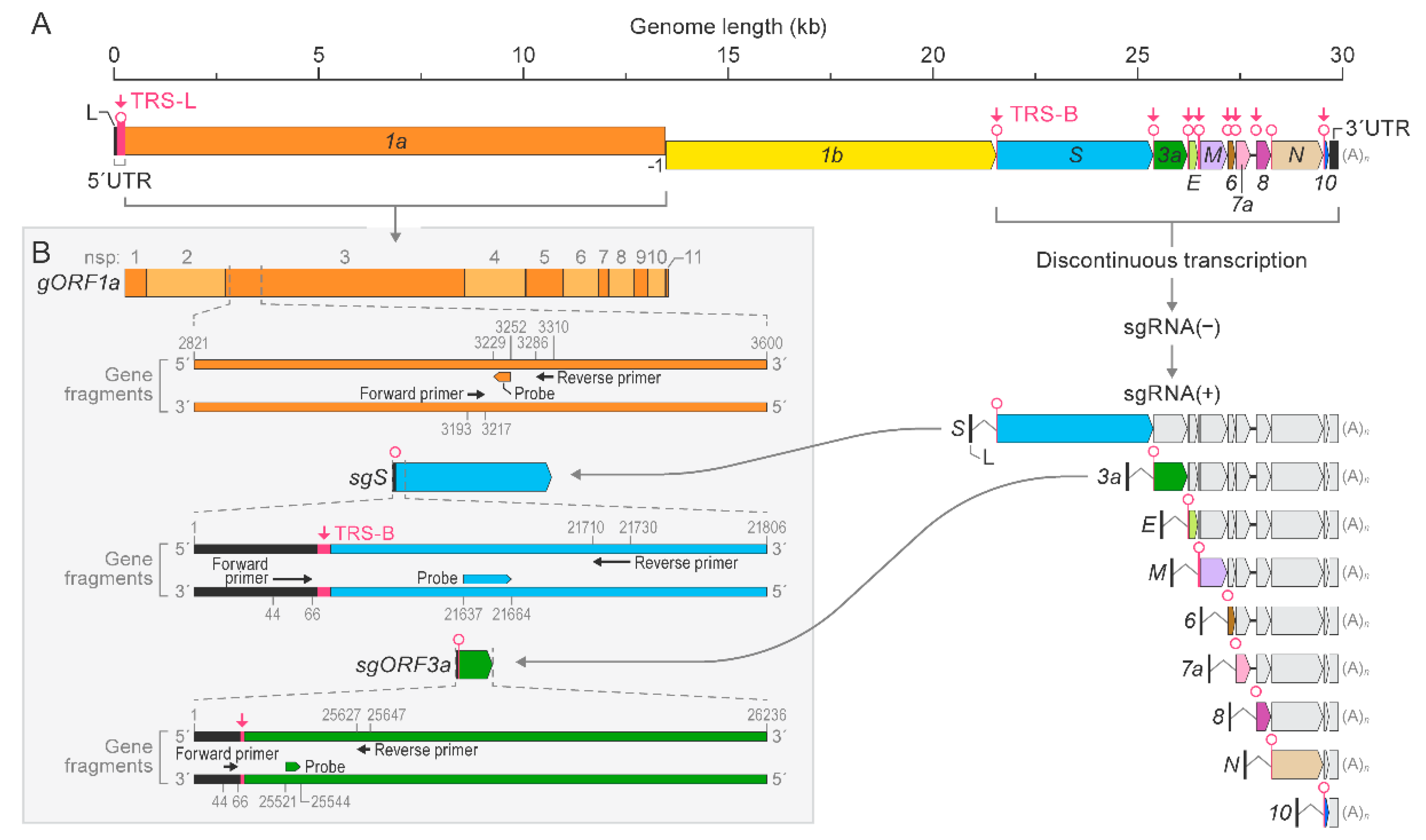
2.4. Gene Fragment Synthesis
2.5. RT-qPCR Assays
2.6. Time-Course Experiments
2.7. Statistics
3. Results
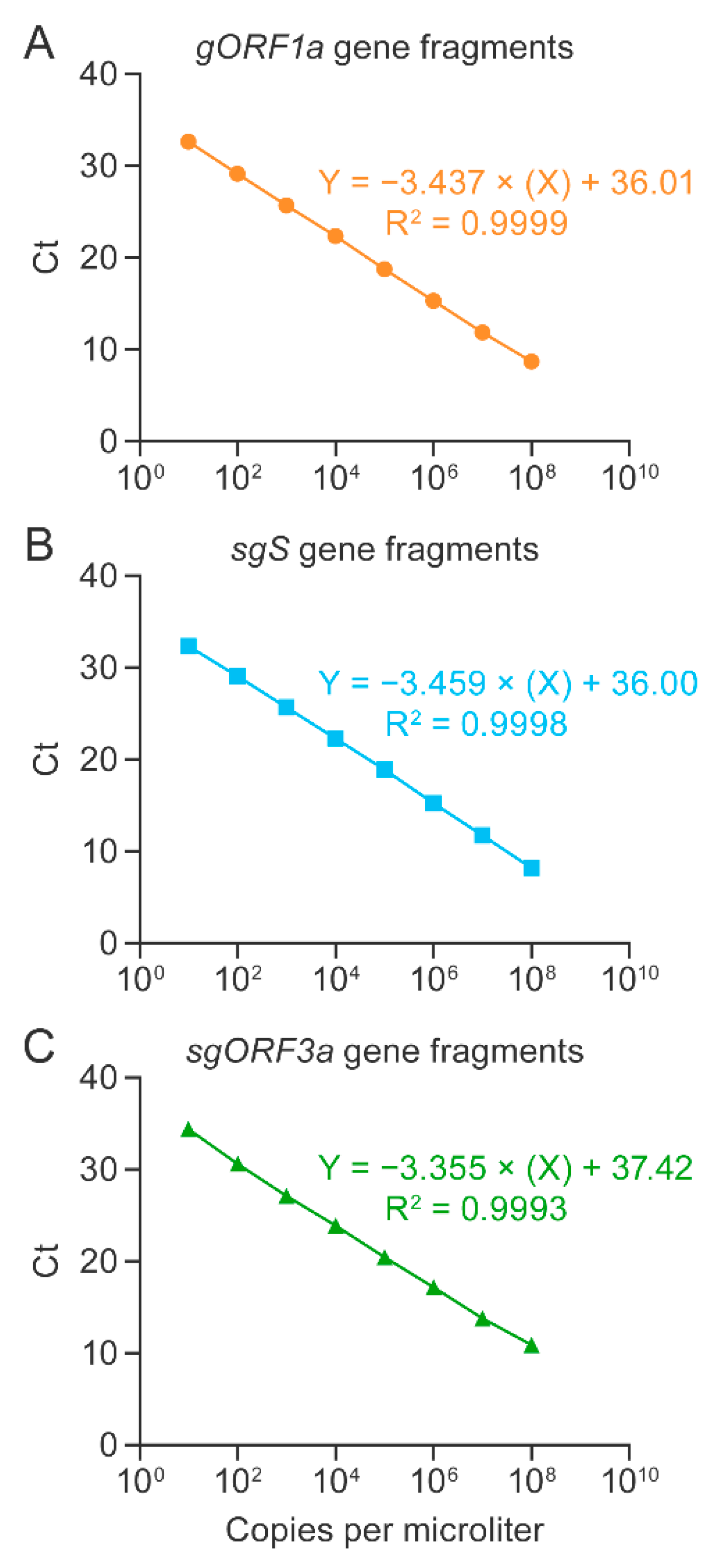
3.1. Singleplex RT-qPCR Assays Were Established and Optimized for the Detection of SARS-CoV-2 Genomic and Subgenomic Nucleic Acids Using Synthetic Gene Fragments
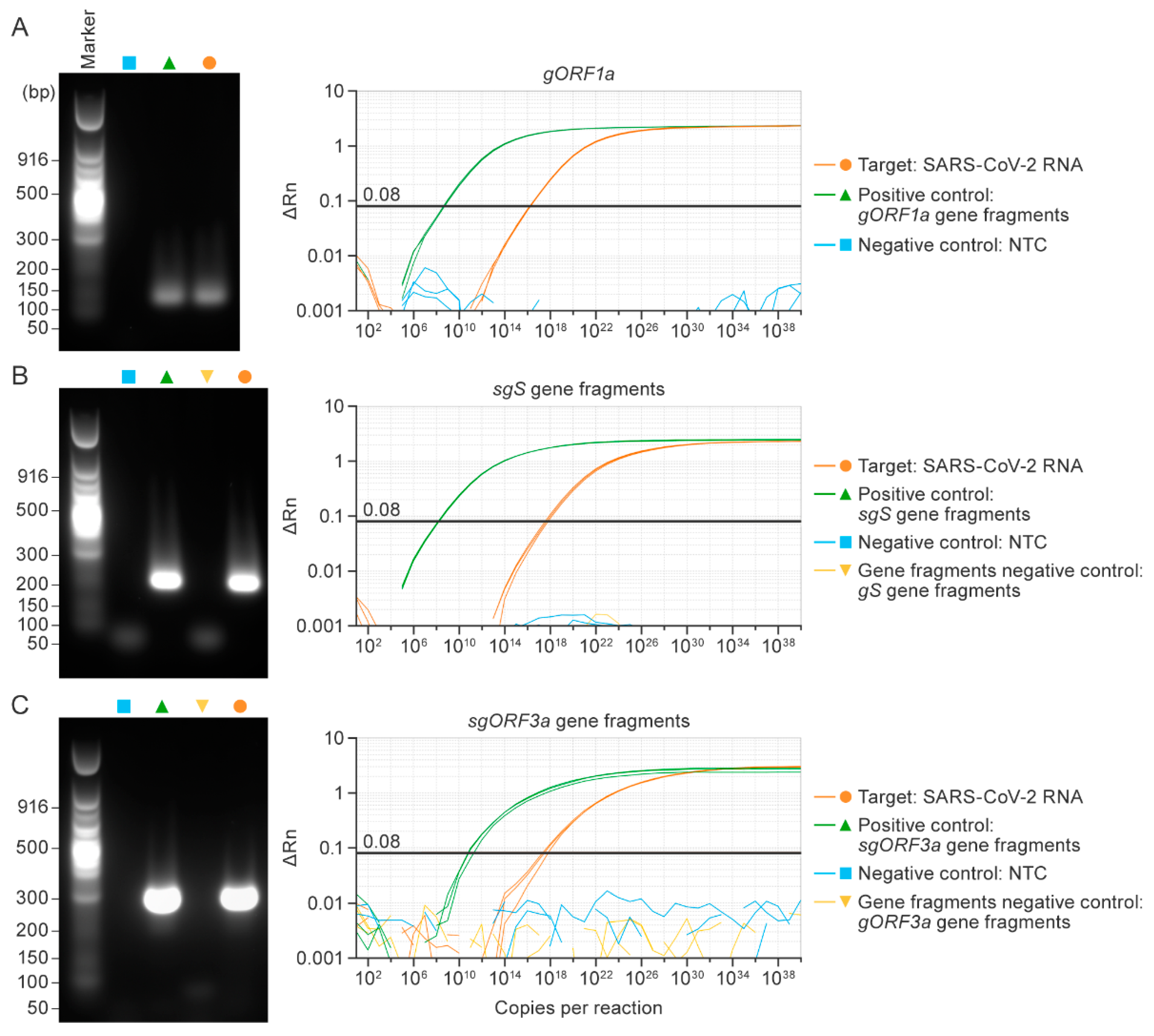
3.2. Singleplex RT-qPCR Assays Specifically Detected SARS-CoV-2 Genomic or Subgenomic Nucleic Acids
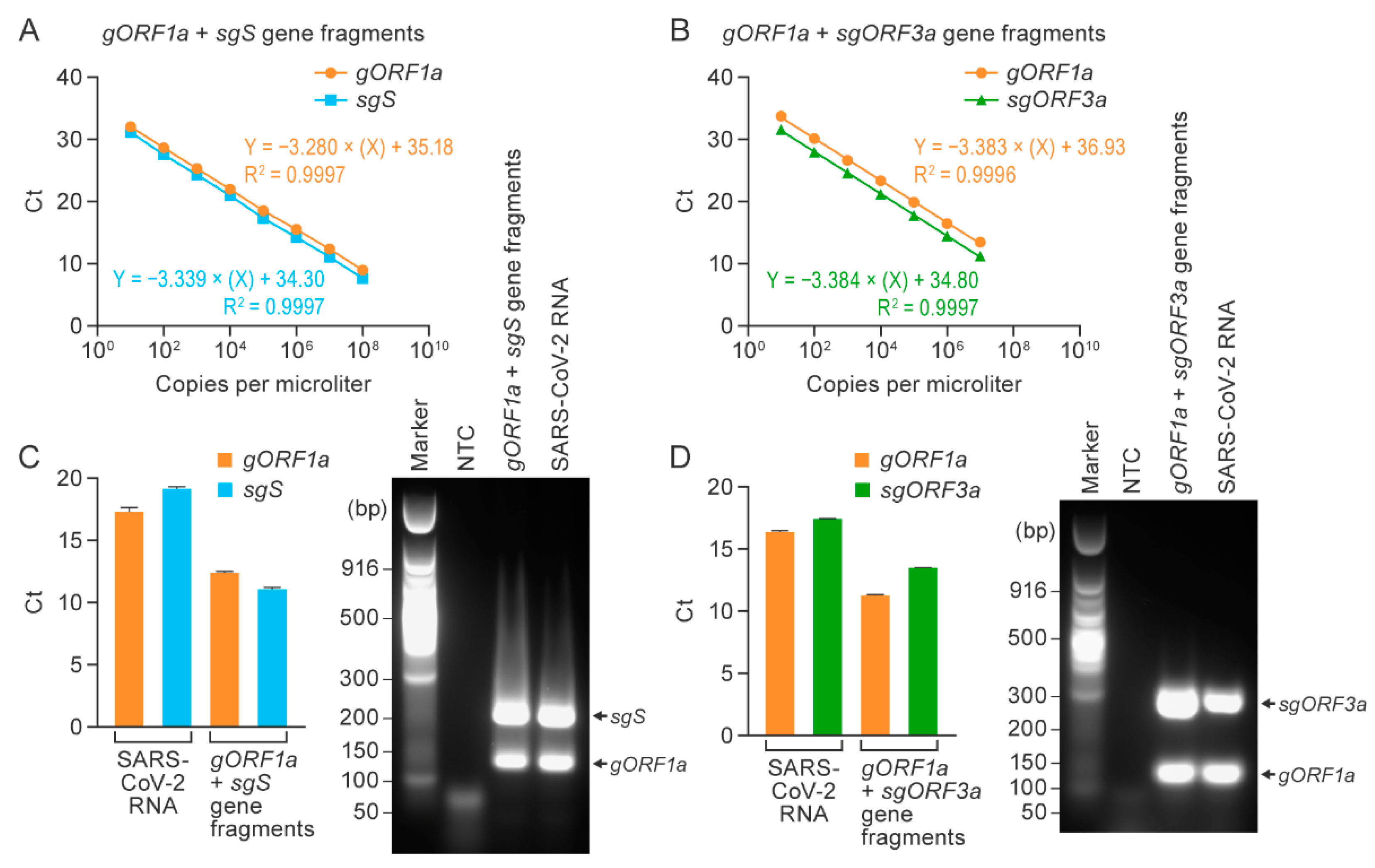
3.3. Duplex RT-qPCR Assays Specifically Detected SARS-CoV-2 Genomic and Subgenomic Nucleic Acids
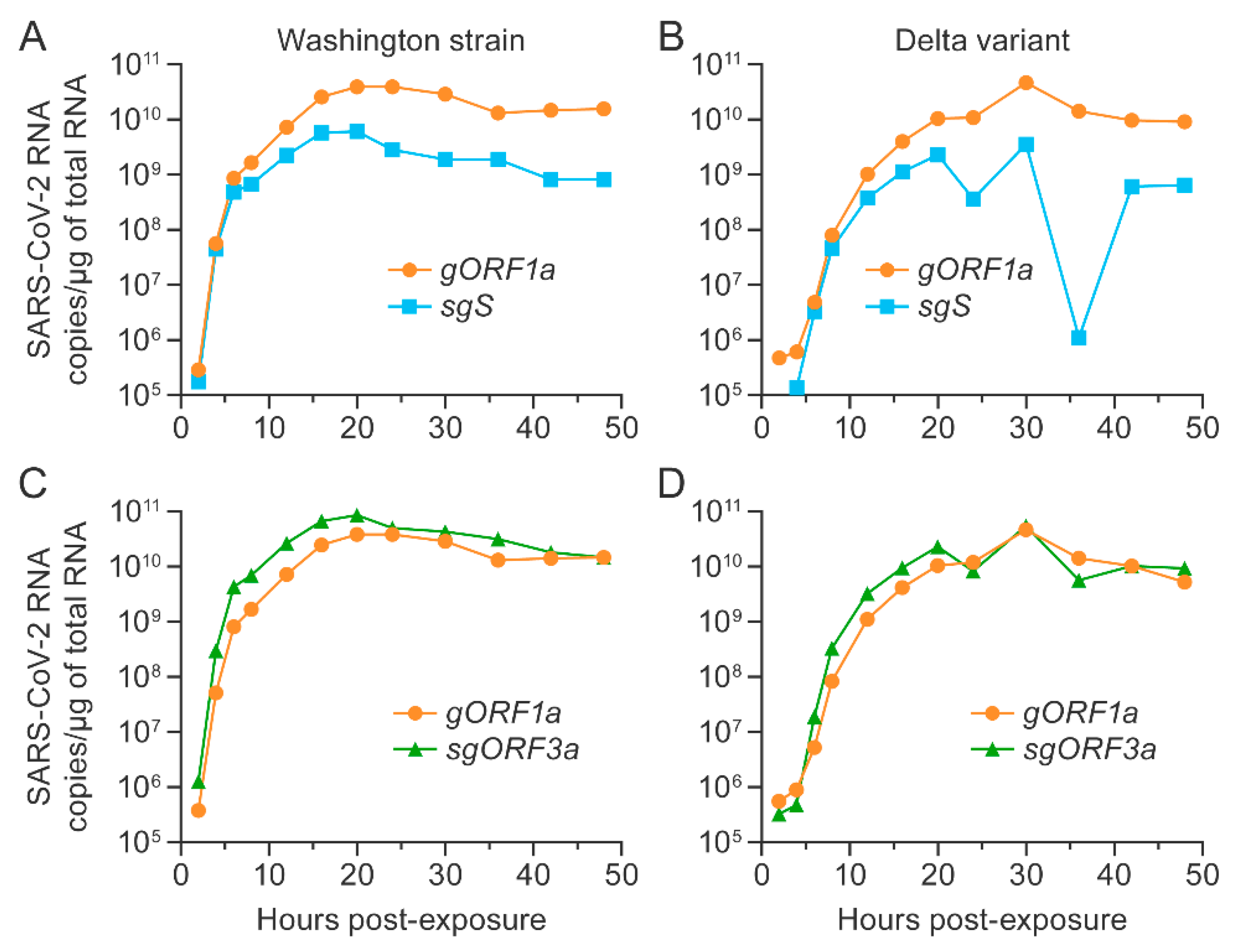
3.4. SARS-CoV-2 Washington Strain and Delta Variant Genomic and Subgenomic RNAs Were Longitudinally Detected in Vero E6 Cells Expressing TMPRSS2
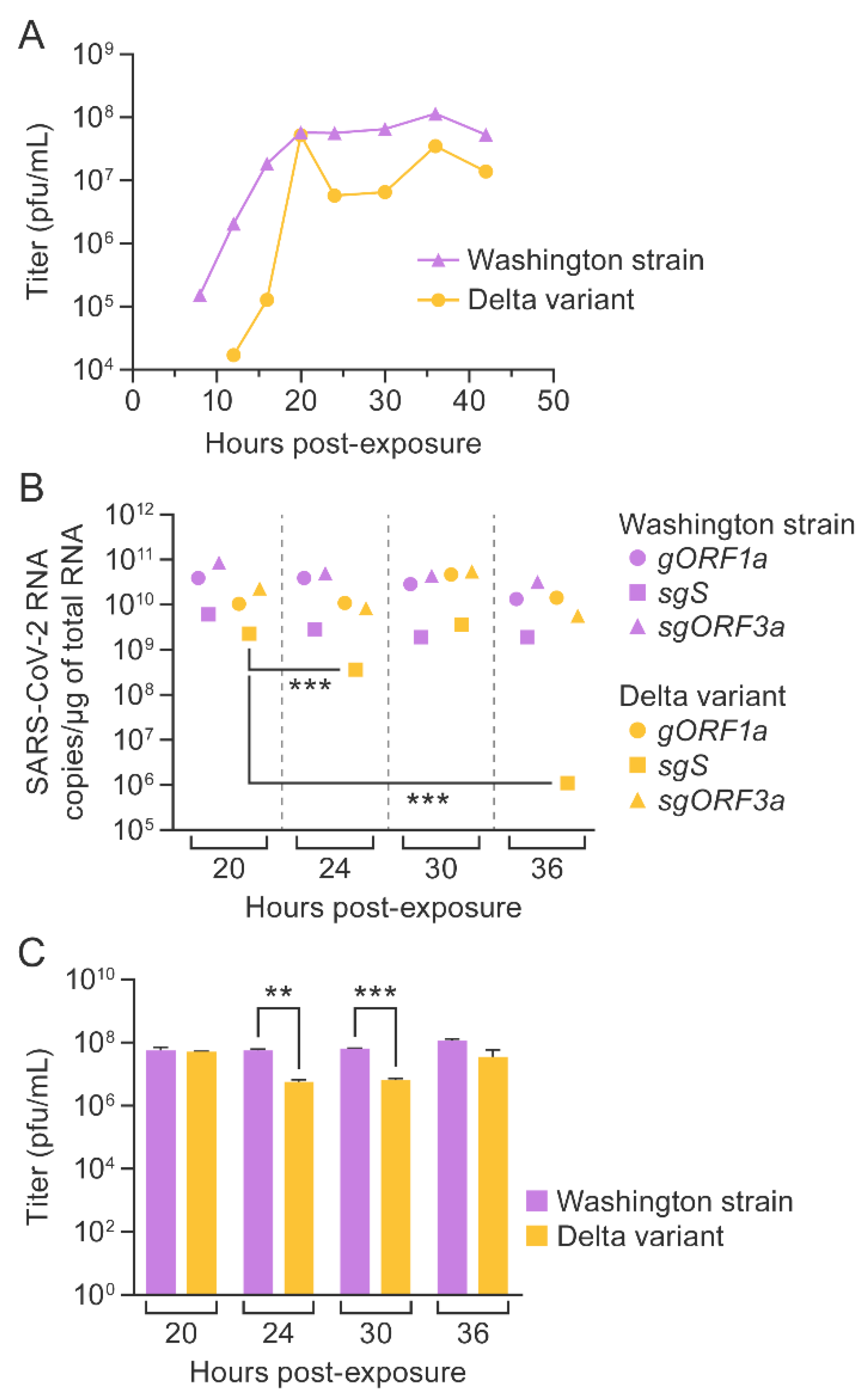
3.5. SARS-CoV-2 Washington Strain and Delta Variant Titers Correlated with Genomic and Subgenomic RNA Copy Numbers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boshier, F.A.T.; Pang, J.; Penner, J.; Parker, M.; Alders, N.; Bamford, A.; Grandjean, L.; Grunewald, S.; Hatcher, J.; Best, T.; et al. Evolution of viral variants in remdesivir-treated and untreated SARS-CoV-2-infected pediatrics patients. J. Med. Virol. 2022, 94, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Johns Hopkins University & Medicine Coronavirus Resource Center. COVID-19 Dashboard. 2022. Available online: https://coronavirus.jhu.edu/map.html (accessed on 4 January 2022).
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Gulyaeva, A.A.; Gorbalenya, A.E. A nidovirus perspective on SARS-CoV-2. Biochem. Biophys. Res. Commun. 2021, 538, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early transmission dynamics in Wuhan, China, of novel coronavirus-infected pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Yuan, S.; Kok, K.-H.; To, K.K.-W.; Chu, H.; Yang, J.; Xing, F.; Liu, J.; Yip, C.C.-Y.; Poon, R.W.-S.; et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: A study of a family cluster. Lancet 2020, 395, 514–523. [Google Scholar] [CrossRef] [Green Version]
- Konings, F.; Perkins, M.D.; Kuhn, J.H.; Pallen, M.J.; Alm, E.J.; Archer, B.N.; Barakat, A.; Bedford, T.; Bhiman, J.N.; Caly, L.; et al. SARS-CoV-2 variants of interest and concern naming scheme conducive for global discourse. Nat. Microbiol. 2021, 6, 821–823. [Google Scholar] [CrossRef]
- Rotondo, J.C.; Martini, F.; Maritati, M.; Mazziotta, C.; Di Mauro, G.; Lanzillotti, C.; Barp, N.; Gallerani, A.; Tognon, M.; Contini, C. SARS-CoV-2 Infection: New Molecular, Phylogenetic, and Pathogenetic Insights. Efficacy of Current Vaccines and the Potential Risk of Variants. Viruses 2021, 13, 1687. [Google Scholar] [CrossRef]
- Rahimi, A.; Mirzazadeh, A.; Tavakolpour, S. Genetics and genomics of SARS-CoV-2: A review of the literature with the special focus on genetic diversity and SARS-CoV-2 genome detection. Genomics 2021, 113, 1221–1232. [Google Scholar] [CrossRef]
- Kim, D.; Lee, J.-Y.; Yang, J.-S.; Kim, J.W.; Kim, V.N.; Chang, H. The architecture of SARS-CoV-2 transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef]
- V’Kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, S.; Zhou, L.; Wang, X.; Yang, H.; Li, W. Coronavirus Disease 2019 (COVID-19): Emerging detection technologies and auxiliary analysis. J. Clin. Lab. Anal. 2022, 36, e24152. [Google Scholar] [CrossRef] [PubMed]
- Zollo, M.; Ferrucci, V.; Izzo, B.; Quarantelli, F.; Domenico, C.D.; Comegna, M.; Paolillo, C.; Amato, F.; Siciliano, R.; Castaldo, G.; et al. SARS-CoV-2 subgenomic N (sgN) transcripts in oro-nasopharyngeal swabs correlate with the highest viral load, as evaluated by five different molecular methods. Diagnostics 2021, 11, 288. [Google Scholar] [CrossRef] [PubMed]
- Penarrubia, L.; Ruiz, M.; Porco, R.; Rao, S.N.; Juanola-Falgarona, M.; Manissero, D.; Lopez-Fontanals, M.; Pareja, J. Multiple assays in a real-time RT-PCR SARS-CoV-2 panel can mitigate the risk of loss of sensitivity by new genomic variants during the COVID-19 outbreak. Int. J. Infect. Dis. 2020, 97, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Shimizu, T.; Noguchi, T.; Nakano, S.; Yamamoto, M.; Nagao, M. Comparison of 12 molecular detection assays for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). J. Mol. Diagn. 2021, 23, 164–170. [Google Scholar] [CrossRef]
- Dharavath, B.; Yadav, N.; Desai, S.; Sunder, R.; Mishra, R.; Ketkar, M.; Bhanshe, P.; Gupta, A.; Redhu, A.K.; Patkar, N.; et al. A one-step, one-tube real-time RT-PCR based assay with an automated analysis for detection of SARS-CoV-2. Heliyon 2020, 6, e04405. [Google Scholar] [CrossRef]
- Petrillo, S.; Carrà, G.; Bottino, P.; Zanotto, E.; De Santis, M.C.; Margaria, J.P.; Giorgio, A.; Mandili, G.; Martini, M.; Cavallo, R.; et al. A novel multiplex qRT-PCR assay to detect SARS-CoV-2 infection: High sensitivity and increased testing capacity. Microorganisms 2020, 8, 1064. [Google Scholar] [CrossRef]
- da Silva Queiroz, J.A.; Rampazzo, R.C.P.; da Silva Filhoa, E.B.; Oliveira, G.S.; da Costa Oliveira, S.; Souza, L.F.B.; dos Santos Pereira, S.; de Souza Rodrigues, M.M.; Maia, A.C.S.; da Silva, C.C.; et al. Development of a quantitative one-step multiplex RT-qPCR assay for the detection of SARS-CoV-2 in a biological matrix. Int J. Infect. Dis. 2021, 104, 373–378. [Google Scholar] [CrossRef]
- Kudo, E.; Israelow, B.; Vogels, C.B.F.; Lu, P.; Wyllie, A.L.; Tokuyama, M.; Venkataraman, A.; Brackney, D.E.; Ott, I.M.; Petrone, M.E.; et al. Detection of SARS-CoV-2 RNA by multiplex RT-qPCR. PLoS Biol. 2020, 18, e3000867. [Google Scholar] [CrossRef]
- Laverack, M.; Tallmadge, R.L.; Venugopalan, R.; Cronk, B.; Zhang, X.; Rauh, R.; Saunders, A.; Nelson, W.M.; Plocharczyk, E.; Diel, D.G. Clinical evaluation of a multiplex real-time RT-PCR assay for detection of SARS-CoV-2 in individual and pooled upper respiratory tract samples. Arch. Virol. 2021, 166, 2551–2561. [Google Scholar] [CrossRef]
- Ishige, T.; Murata, S.; Taniguchi, T.; Miyabe, A.; Kitamura, K.; Kawasaki, K.; Nishimura, M.; Igari, H.; Matsushita, K. Highly sensitive detection of SARS-CoV-2 RNA by multiplex rRT-PCR for molecular diagnosis of COVID-19 by clinical laboratories. Clin. Chim. Acta 2020, 507, 139–142. [Google Scholar] [CrossRef]
- Waggoner, J.J.; Stittleburg, V.; Pond, R.; Saklawi, Y.; Sahoo, M.K.; Babiker, A.; Hussaini, L.; Kraft, C.S.; Pinsky, B.A.; Anderson, E.J.; et al. Triplex real-time RT-PCR for severe acute respiratory syndrome coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1633–1635. [Google Scholar] [CrossRef]
- Reijns, M.A.M.; Thompson, L.; Acosta, J.C.; Black, H.A.; Sanchez-Luque, F.J.; Diamond, A.; Parry, D.A.; Daniels, A.; O’Shea, M.; Uggenti, C.; et al. A sensitive and affordable multiplex RT-qPCR assay for SARS-CoV-2 detection. PLoS Biol. 2020, 18, e3001030. [Google Scholar] [CrossRef]
- Gadkar, V.J.; Goldfarb, D.M.; Young, V.; Watson, N.; Al-Rawahi, G.N.; Srigley, J.A.; Tilley, P. Development and validation of a new triplex real-time quantitative reverse Transcriptase-PCR assay for the clinical detection of SARS-CoV-2. Mol. Cell. Probes 2021, 58, 101744. [Google Scholar] [CrossRef] [PubMed]
- Bland, J.; Kavanaugh, A.; Hong, L.K.; Perez, O.; Kadkol, S.S. A multiplex one-step RT-qPCR protocol to detect SARS-CoV-2 in NP/OP swabs and saliva. Curr. Protoc. 2021, 1, e145. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Lu, J.; Wang, N.; He, W.-T.; Zhang, L.; Zhao, W.; Su, S. Development of a TaqMan-probe-based multiplex real-time PCR for the simultaneous detection of emerging and reemerging swine coronaviruses. Virulence 2020, 11, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Mancini, F.; Barbanti, F.; Scaturro, M.; Fontana, S.; Di Martino, A.; Marsili, G.; Puzelli, S.; Calzoletti, L.; Facchini, M.; Di Mario, G.; et al. Multiplex real-time reverse-transcription polymerase chain reaction assays for diagnostic testing of severe acute respiratory syndrome coronavirus 2 and seasonal influenza viruses: A challenge of the phase 3 pandemic setting. J. Infect. Dis. 2021, 223, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Pabbaraju, K.; Wong, A.A.; Ma, R.; Zelyas, N.; Tipples, G.A. Development and validation of a multiplex reverse transcriptase-PCR assay for simultaneous testing of influenza A, influenza B and SARS-CoV-2. J. Virol. Methods 2021, 293, 114151. [Google Scholar] [CrossRef]
- Shu, B.; Kirby, M.K.; Davis, W.G.; Warnes, C.; Liddell, J.; Liu, J.; Wu, K.-H.; Hassell, N.; Benitez, A.J.; Wilson, M.M.; et al. Multiplex real-time reverse transcription PCR for influenza A virus, influenza B virus, and severe acute respiratory syndrome coronavirus 2. Emerg. Infect. Dis. 2021, 27, 1821–1830. [Google Scholar] [CrossRef]
- Vogels, C.B.F.; Breban, M.I.; Ott, I.M.; Alpert, T.; Petrone, M.E.; Watkins, A.E.; Kalinich, C.C.; Earnest, R.; Rothman, J.E.; Goes de Jesus, J.; et al. Multiplex qPCR discriminates variants of concern to enhance global surveillance of SARS-CoV-2. PLoS Biol. 2021, 19, e3001236. [Google Scholar] [CrossRef]
- Telwatte, S.; Martin, H.A.; Marczak, R.; Fozouni, P.; Vallejo-Gracia, A.; Kumar, G.R.; Murray, V.; Lee, S.; Ott, M.; Wong, J.K.; et al. Novel RT-ddPCR assays for measuring the levels of subgenomic and genomic SARS-CoV-2 transcripts. Methods 2021, 201, 15–25. [Google Scholar] [CrossRef]
- Dagotto, G.; Mercado, N.B.; Martinez, D.R.; Hou, Y.J.; Nkolola, J.P.; Carnahan, R.H.; Crowe, J.E., Jr.; Baric, R.S.; Barouch, D.H. Comparison of subgenomic and total RNA in SARS-CoV-2 challenged rhesus macaques. J. Virol. 2021, 95, e02370-20. [Google Scholar] [CrossRef] [PubMed]
- Dimcheff, D.E.; Valesano, A.L.; Rumfelt, K.E.; Fitzsimmons, W.J.; Blair, C.; Mirabelli, C.; Petrie, J.G.; Martin, E.T.; Bhambhani, C.; Tewari, M.; et al. Severe acute respiratory syndrome coronavirus 2 total and subgenomic RNA viral load in hospitalized patients. J. Infect. Dis. 2021, 224, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.; Kim, E.; Martínez-Colón, G.J.; Jagannathan, P.; Rustagi, A.; Parsonnet, J.; Bonilla, H.; Khosla, C.; Holubar, M.; Subramanian, A.; et al. SARS-CoV-2 subgenomic RNA kinetics in longitudinal clinical samples. Open Forum Infect. Dis. 2021, 8, ofab310. [Google Scholar] [CrossRef] [PubMed]
- Almehdi, A.M.; Khoder, G.; Alchakee, A.S.; Alsayyid, A.T.; Sarg, N.H.; Soliman, S.S.M. SARS-CoV-2 spike protein: Pathogenesis, vaccines, and potential therapies. Infection 2021, 49, 855–876. [Google Scholar] [CrossRef]
- Durmaz, B.; Abdulmajed, O.; Durmaz, R. Mutations observed in the SARS-CoV-2 spike glycoprotein and their effects in the interaction of virus with ACE-2 receptor. Medeni. Med. J. 2020, 35, 253–260. [Google Scholar] [CrossRef]
- Redondo, N.; Zaldívar-López, S.; Garrido, J.J.; Montoya, M. SARS-CoV-2 accessory proteins in viral pathogenesis: Knowns and unknowns. Front. Immunol. 2021, 12, 708264. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, S.; Fang, Y.; Bollinger, L.; Li, Y.; Lauck, M.; Postnikova, E.N.; Mazur, S.; Johnson, R.F.; Finch, C.L.; et al. Development and characterization of a cDNA-launch recombinant simian hemorrhagic fever virus expressing enhanced green fluorescent protein: ORF 2b’ is not required for in vitro virus replication. Viruses 2021, 13, 632. [Google Scholar] [CrossRef]
- Bennett, R.P.; Postnikova, E.N.; Eaton, B.P.; Cai, Y.; Yu, S.; Smith, C.O.; Liang, J.; Zhou, H.; Kocher, G.A.; Murphy, M.J.; et al. Sangivamycin is highly effective against SARS-CoV-2 in vitro and has favorable drug properties. JCI Insight 2022, 7, e153165. [Google Scholar] [CrossRef]
- Kimble, K.M.; Dickinson, S.E.; Biase, F.H. Extraction of total RNA from single-oocytes and single-cell mRNA sequencing of swine oocytes. BMC Res. Notes 2018, 11, 155. [Google Scholar] [CrossRef]
- Fan, G.; Qin, X.; Streblow, D.N.; Hoyos, C.M.; Hansel, D.E. Comparison of SARS-CoV-2 PCR-Based Detection Using Saliva or Nasopharyngeal Swab Specimens in Asymptomatic Populations. Microbiol. Spectr. 2021, 9, e0006221. [Google Scholar] [CrossRef]
- Speranza, E.; Williamson, B.N.; Feldmann, F.; Sturdevant, G.L.; Perez-Perez, L.; Meade-White, K.; Smith, B.J.; Lovaglio, J.; Martens, C.; Munster, V.J.; et al. Single-cell RNA sequencing reveals SARS-CoV-2 infection dynamics in lungs of African green monkeys. Sci. Transl. Med. 2021, 13, eabe8146. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Dankova, Z.; Novakova, E.; Skerenova, M.; Holubekova, V.; Lucansky, V.; Dvorska, D.; Brany, D.; Kolkova, Z.; Strnadel, J.; Mersakova, S.; et al. Comparison of SARS-CoV-2 detection by rapid antigen and by three commercial RT-qPCR tests: A study from Martin University Hospital in Slovakia. Int. J. Environ. Res. Public Health 2021, 18, 7037. [Google Scholar] [CrossRef] [PubMed]
- Cuong, H.Q.; Hai, N.D.; Linh, H.T.; Anh, N.H.; Hieu, N.T.; Thang, C.M.; Thao, N.T.T.; Lan, P.T. Comparison of primer-probe sets among different master mixes for laboratory screening of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). BioMed Res. Int. 2020, 2020, 7610678. [Google Scholar] [CrossRef] [PubMed]
- Haddar, C.; Verhoeven, P.O.; Bourlet, T.; Pozzetto, B.; Pillet, S. Brief comparative evaluation of six open one-step RT-qPCR mastermixes for the detection of SARS-CoV-2 RNA using a Taqman probe. J. Clin. Virol. 2020, 132, 104636. [Google Scholar] [CrossRef]
- Matsuyama, S.; Nao, N.; Shirato, K.; Kawase, M.; Saito, S.; Takayama, I.; Nagata, N.; Sekizuka, T.; Katoh, H.; Kato, F.; et al. Enhanced isolation of SARS-CoV-2 by TMPRSS2-expressing cells. Proc. Natl. Acad. Sci. USA 2020, 117, 7001–7003. [Google Scholar] [CrossRef] [Green Version]
- Syed, A.M.; Taha, T.Y.; Tabata, T.; Chen, I.-P.; Ciling, A.; Khalid, M.M.; Sreekumar, B.; Chen, P.Y.; Hayashi, J.M.; Soczek, K.M.; et al. Rapid assessment of SARS-CoV-2 evolved variants using virus-like particles. Science 2021, 374, 1626–1632. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, T.; Cai, Y.; Lavine, C.L.; Peng, H.; Zhu, H.; Anand, K.; Tong, P.; Gautam, A.; Mayer, M.L.; et al. Membrane fusion and immune evasion by the spike protein of SARS-CoV-2 Delta variant. Science 2021, 374, 1353–1360. [Google Scholar] [CrossRef]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR-Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [Green Version]
- Chiara, M.; D’Erchia, A.M.; Gissi, C.; Manzari, C.; Parisi, A.; Resta, N.; Zambelli, F.; Picardi, E.; Pavesi, G.; Horner, D.S.; et al. Next generation sequencing of SARS-CoV-2 genomes: Challenges, applications and opportunities. Brief. Bioinform. 2021, 22, 616–630. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Regions | Primers and Probes | Sequences (5′–3′) |
|---|---|---|
| Genomic ORF1a | gORF1a-F gORF1a-R gORF1a-P | AGAAGATTGGTTAGATGATGATAGT TTCCATCTCTAATTGAGGTTGAACC FAM/TCCTCACTGCCGTCTTGTTGACCA/BHQ13 |
| Subgenomic S | sgS-F sgS-R sgS-P | CGATCTCTTGTAGATCTGTTCTC TAAGAACAAGTCCTGAGTTGA SUN/CCCTGCATA/ZEN/CACTAATTCTTTCACACGT/IABKFQ |
| Subgenomic ORF3a | sgORF3a-F sgORF3a-R sgORF3a-P | CGATCTCTTGTAGATCTGTTCTC CAACAGCAAGTTGCAAACAAA ABY/CGGATGGCTTATTGTTGGCGTTGC/QSY7 |
| Assay | Target Regions | Ct Mean | Ct SD |
|---|---|---|---|
| Singleplex | Genomic ORF1a | 16.620 | 0.033 |
| Subgenomic S | 17.669 | 0.130 | |
| Subgenomic ORF3a | 17.517 | 0.212 | |
| Duplex ORF1A/sgS | Genomic ORF1a | 17.397 | 0.213 |
| Subgenomic S | 19.160 | 0.184 | |
| Duplex ORF1a/sgORF3a | Genomic ORF1a | 16.981 | 0.181 |
| Subgenomic ORF3a | 18.399 | 0.103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhosle, S.M.; Tran, J.P.; Yu, S.; Geiger, J.; Jackson, J.D.; Crozier, I.; Crane, A.; Wada, J.; Warren, T.K.; Kuhn, J.H.; et al. Duplex One-Step RT-qPCR Assays for Simultaneous Detection of Genomic and Subgenomic RNAs of SARS-CoV-2 Variants. Viruses 2022, 14, 1066. https://doi.org/10.3390/v14051066
Bhosle SM, Tran JP, Yu S, Geiger J, Jackson JD, Crozier I, Crane A, Wada J, Warren TK, Kuhn JH, et al. Duplex One-Step RT-qPCR Assays for Simultaneous Detection of Genomic and Subgenomic RNAs of SARS-CoV-2 Variants. Viruses. 2022; 14(5):1066. https://doi.org/10.3390/v14051066
Chicago/Turabian StyleBhosle, Sushma M., Julie P. Tran, Shuiqing Yu, Jillian Geiger, Jennifer D. Jackson, Ian Crozier, Anya Crane, Jiro Wada, Travis K. Warren, Jens H. Kuhn, and et al. 2022. "Duplex One-Step RT-qPCR Assays for Simultaneous Detection of Genomic and Subgenomic RNAs of SARS-CoV-2 Variants" Viruses 14, no. 5: 1066. https://doi.org/10.3390/v14051066