
Delivery Routes for COVID-19 Vaccines


Graduate School of Medical Science and Engineering, Korea Advanced Institute of Science and Technology (KAIST), Daejeon 34141, Korea
*
Author to whom correspondence should be addressed.
Vaccines 2021, 9(5), 524; https://doi.org/10.3390/vaccines9050524
Submission received: 15 April 2021
/
Revised: 12 May 2021
/
Accepted: 17 May 2021
/
Published: 19 May 2021
(This article belongs to the Special Issue Recent Advances in Mucosal Vaccines)
Abstract
:The novel coronavirus, SARS-CoV-2, which causes COVID-19, has resulted in a pandemic with millions of deaths. To eradicate SARS-CoV-2 and prevent further infections, many vaccine candidates have been developed. These vaccines include not only traditional subunit vaccines and attenuated or inactivated viral vaccines but also nucleic acid and viral vector vaccines. In contrast to the diversity in the platform technology, the delivery of vaccines is limited to intramuscular vaccination. Although intramuscular vaccination is safe and effective, mucosal vaccination could improve the local immune responses that block the spread of pathogens. However, a lack of understanding of mucosal immunity combined with the urgent need for a COVID-19 vaccine has resulted in only intramuscular vaccinations. In this review, we summarize the history of vaccines, current progress in COVID-19 vaccine technology, and the status of intranasal COVID-19 vaccines. Future research should determine the most effective route for vaccine delivery based on the platform and determine the mechanisms that underlie the efficacy of different delivery routes.
1. Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), a novel virus that causes coronavirus disease 2019 (COVID-19), is responsible for a pandemic that has resulted in millions of deaths across the globe. There is an urgent need for effective vaccines to prevent COVID-19 and eradicate SARS-CoV-2, and many companies are developing and testing new vaccines. These include not only traditional vaccines, such as inactivated and attenuated viruses or subunit vaccines, but also new RNA, DNA, and viral vector vaccines [1]. Despite advances in vaccine platform technology, research on the most effective delivery routes for vaccines is limited. Since SARS-CoV-2 infects via the respiratory tract, intranasal vaccination should be effective. However, poor understanding of mucosal vaccines has limited their development to the stage of human clinical trials.
While the angiotensin-converting enzyme (ACE2) receptor for SARS-CoV-2 is found throughout the respiratory tract and in the brain, placenta, and gut, the first line of defense against infection is the nasal epithelium [2]. Intramuscular immunization induces an immune response in the lower respiratory tract (LRT), but not in the upper respiratory tract (URT). In contrast, intranasal immunization provides immunity not only in the URT but also provides systemic immunity [3,4]. Mucosal IgA is known to protect against the shedding of nasal virus early in infection, while the level of systemic IgA is correlated with severe disease. However, mucosal immunity can be difficult to establish because the mucosa is frequently exposed to and becomes tolerant of foreign molecules. Additionally, innate mucosal defense systems such as proteolytic enzymes present a barrier to antigen absorption [5]. For the development of effective mucosal vaccines, we need a better understanding of the mucosal immune environment. Here we review recent progress on delivery routes for COVID-19 vaccines and discuss future perspectives.
2. Immune Responses against COVID-19
Understanding the immune responses to SARS-CoV-2 infection is important for designing vaccines for COVID-19. SARS-CoV-2 binds target cells through the ACE2 receptor [6], and then serine protease TMPRRSS2, neuropilin 1, and furins facilitate SARS-CoV-2 infection [7]. As these proteins are broadly expressed in multiple organs including the nasal airway, the lung, and the placenta, SARS-CoV-2 can spread through the body [8]. It is also possible that SARS-CoV-2 can infect immune cells since in vitro infection of monocyte-derived macrophages and dendritic cells (DCs) has been observed and viral RNA was detected in SPP1+ macrophages [9].
2.1. Innate Immune Response
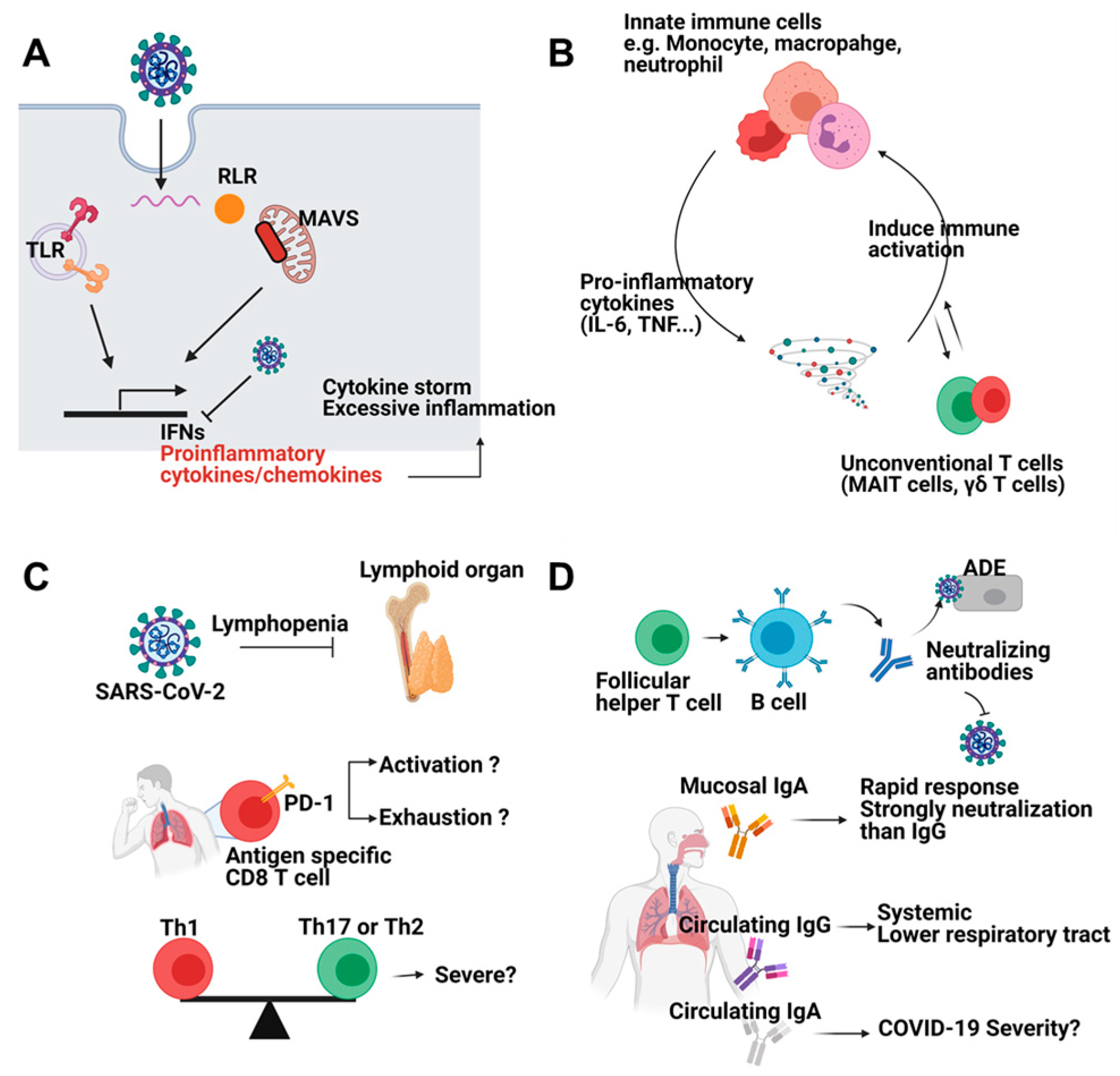
After entry of SARS-CoV-2, multiple pattern recognition receptors (PRRs) including Toll-like receptor 3 (TLR3), TLR7, retinoic acid-inducible gene 1 (RIG-I), and melanoma differentiation-associated gene 5 (MDA5) sense viral infection [7]. PRR recognition triggers phosphorylation of Interferon Regulatory Factor 3 (IRF3) and IRF7 which regulate type I interferon (IFN) and interferon-stimulated genes (ISGs) [10]. SARS-CoV-2-derived nonstructural proteins suppress type I IFN production [11]. As a result, SARS-CoV-2 induces low-level type I and II IFNs alongside high-level induction of proinflammatory cytokines and chemokines [12]. Although a normal immune response protects the host from infection, cytokine release syndrome (CRS), the so-called “cytokine storm”, is frequently observed in patients with severe COVID-19 (Figure 1A) [13].
The main sources of proinflammatory cytokines are monocytes, macrophages, and neutrophils that are recruited and activated by cytokines such as IL-6 and tumor necrosis factor (TNF) or by chemokines. This positive feedback loop increases the severity of the disease [14]. For example, proinflammatory macrophages produce IL-8, which is a chemokine for the recruitment of neutrophils. Neutrophil recruitment correlates with epithelial cell damage and apoptosis which might cause tissue damage and acute respiratory distress syndrome [15]. Programmed death ligand 1 (PD-L1)-expressing neutrophils correlate with severe symptoms while HLA-DRhiCD11chi monocytes show an opposite trend [16]. Mucosal-associated invariant T (MAIT) cells and γδ T cells also correlate with the severity of COVID-19 [17,18]. Although the primary factors leading to uncontrolled inflammation are unknown, the hyperactivated innate immune response is responsible for severe symptoms (Figure 1B).
2.2. Adaptive Immune Response
Consistent with other viral infections, T cells and B cells are a critical component of COVID-19, and T cells are phenotypically different depending on the severity of the disease [19]. COVID-19 patients show lymphopenia, with decreased CD4 T cells, CD8 T cells, and B cells, as well as a preference for natural killer cells over CD8 T cells [20,21]. Many viral infections induce transient lymphopenia [22], but SARS-CoV-2 induces a severe, persistent lymphopenia [19] that may be associated with proinflammatory cytokines or activation-induced expression of proapoptotic molecules [20,23]. CD8 T cells from COVID-19 patients express more inhibitory receptors, such as PD-1 and TIM-3, which correlate with terminal differentiation and functional cell exhaustion [23]. SARS-CoV-2-specific CD8 T cells are CD38+PD-1+ [24]. While it is not clear whether these markers represent cell exhaustion or activation, one study suggests that PD-1-expressing CD8 T cells are functional in COVID-19 patients [25]. CD4 T cells show tendencies similar to CD8 T cells. T helper 1 (Th1) cells correlate with mild disease and CCR6+ CD4 T cells correlate strongly with severe disease (Figure 1C) [26,27]. Subsets of T cells, including follicular helper T cell (Tfh) which support B cell responses, correlate with normal antibody-mediated protection [28]. Interestingly, some people who have not been exposed to SARS-CoV-2 have SARS-CoV-2-specific CD4 T cells, which cross-react with common cold viruses [29]. While this protective immunity disappears within 12 months of infection [30], the potential benefits of this cross-reactivity should be examined.
Antibody-mediated neutralization and antibody-dependent cellular cytotoxicity (ADCC) are critical for the clearing of viruses and prevention of infection. Patients with a mild case of COVID-19 have higher levels of immunoglobulins and more activation markers on B cells than patients with severe disease, but there are more CCR2+ plasma cells recruited to the lungs of patients with severe disease. Although B cell populations differ depending on the severity of the disease, it is not known whether these changes in cell populations affect the severity of the disease [31]. After acute infection, IgM appears first, followed about 4 weeks later by increasing levels of IgG, which may persist for 16–24 months [32]. Patients with mild-to-moderate symptoms mount a robust antibody response, suggesting that this response provides protective immunity [33]. On the contrary, an opposite result showing asymptomatic patients had lower level of IgG than symptomatic patients was also reported [34]. These contradictory results should be addressed. Unlike to IgG, serum IgA might be associated with severe disease [35,36]. However, IgA appears first after infection and has a stronger neutralization capacity than IgG in COVID-19 [37]. Suboptimal antibody levels can induce antibody-dependent enhancement (ADE), which facilitates viral infection; thus, modulating the antibody response could be beneficial to patients (Figure 1D) [38].
3. Current Advances in COVID-19 Vaccines
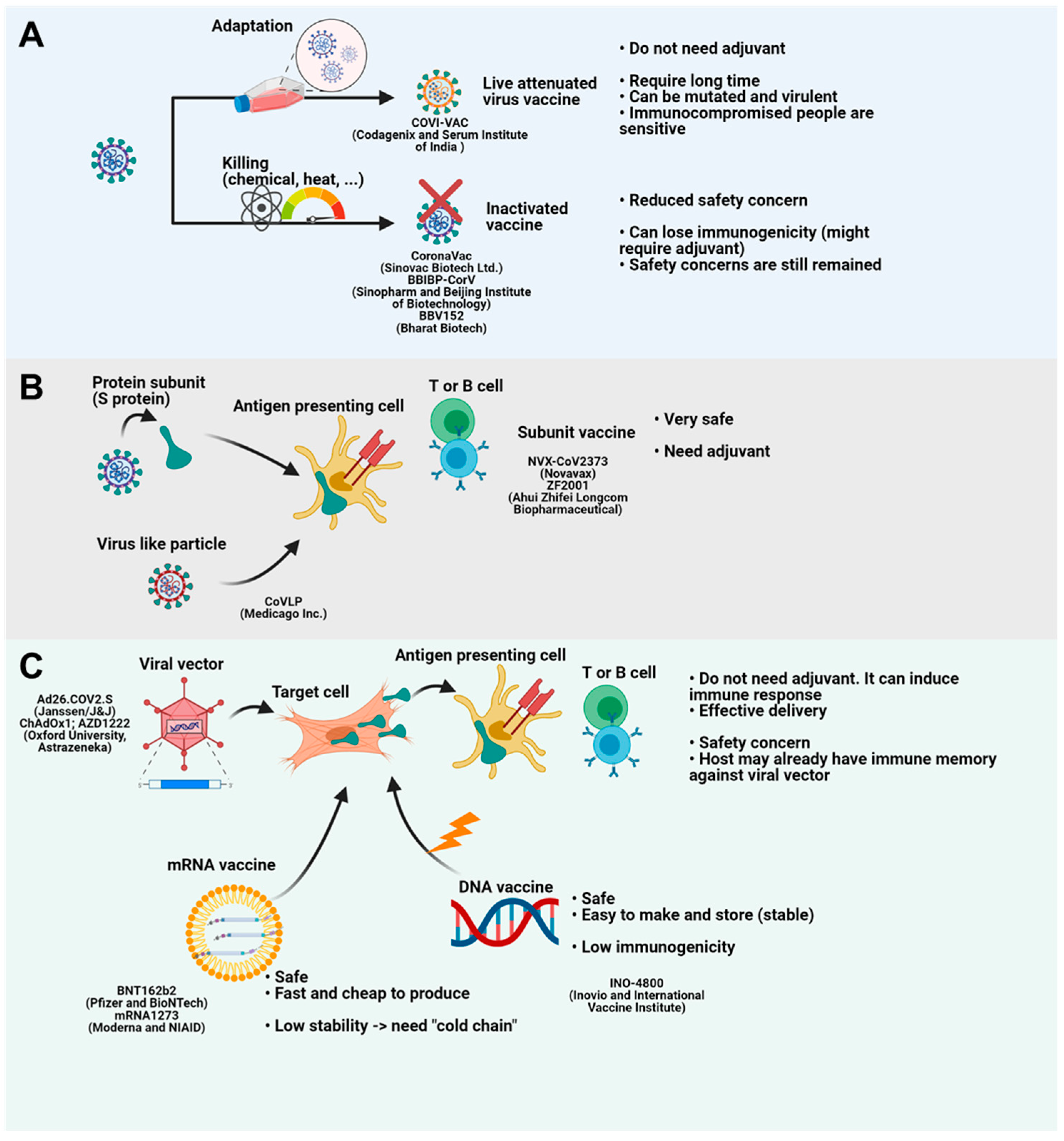
More than 100 vaccines for COVID-19 have been or are being tested (Table 1). In contrast to T cells, which frequently target the NSP7, NSP13, and N proteins of SARS-CoV-2 [39], antibodies predominantly target the N protein and the receptor-binding domain (RBD) of the spike (S) protein [40]. Similarly, vaccines typically target the S protein [1]. Traditionally, vaccines are either attenuated or inactivated pathogens or protein subunits from the pathogen. As early as 1880, attenuated pathogen vaccines were developed by Pasteur [1], and attenuated virus vaccines now prevent measles, mumps, and rubella [41]. However, attenuating strains takes years, so this type of vaccine cannot be used to tackle the current pandemic, and there are also safety concerns. Attenuated pathogens can mutate, becoming more virulent, and immunocompromised people might be susceptible to the attenuated strain [1]. Despite their limitations, live attenuated SARS-CoV-2 vaccines for COVID-19 have been developed, and they have the advantage that they do not require adjuvants. COVI-VAC, developed by Codagenix and the Serum Institute of India (Pune, India), is being evaluated in a phase I clinical trial (NCT04619628). Okamura et al. showed that a SARS-CoV-2 vaccine candidate in a Syrian hamster model protected animals from infection and did not produce serious lung lesions (Figure 2A) [42].
To alleviate safety concerns about live attenuated vaccines, pathogens inactivated by heat, radiation, or chemical treatment have been developed as vaccines. However, inactivated pathogens can lose immunogenicity and typically require additional adjuvants [1]. Sinovac Biotech Ltd. developed CoronaVac, an inactivated virus vaccine for COVID-19. CoronaVac, in which the inactivated virus was adsorbed to the adjuvant aluminum hydroxide, induced neutralizing antibodies in mice, rats, and nonhuman primates [55]. A 3 μg dose was found to be safe and immunogenic for people aged 18–59 and for older people in a phase 1/2 clinical trial [43,56]. CoronaVac is now being evaluated in a phase 3 clinical trial (NCT04456595) [57]. BBIBP-CorV, which was developed by the Beijing Institute of Biotechnology and Sinopharm by adsorbing the virus to aluminum hydroxide [58], was found to be immunogenic, well-tolerated, and safe in a phase 1/2 clinical trial [44]. BBIBP-CorV is now being evaluated in a phase 3 clinical trial in multiple countries. Bharat Biotech developed the BBV152 vaccine which is adsorbed to Algel-IMDG (imidazoquinoline molecule chemisorbed on alum). BBV152 is effective in trafficking antigens to the draining lymph node without systemic circulation and has been demonstrated to be immunogenic, well-tolerated, and safe in mice, rats, Syrian hamsters, and nonhuman primates [59,60,61]. A phase 2 clinical trial was successful and BBV152 is now being evaluated in a phase 3 clinical trial (NCT04641481) (Figure 2A) [41].
Protein subunit vaccines contain synthesized or purified viral proteins that are injected and then processed and presented to adaptive immune cells by antigen-presenting cells (APCs). While subunit vaccines are safer than other vaccines, they require adjuvants and booster shots [1]. NVX-CoV2373, which is being developed by Novavax, contains the SARS-CoV-2 S protein within the matrix-M adjuvant. NVX-CoV2373 has been demonstrated to be immunogenic and safe in mice, baboons, and cynomolgus macaques [62,63], and it successfully induced a Th1 response and IgG in a phase 1/2 clinical trial [46]. This vaccine is now being evaluated in a phase 3 clinical trial. Ahui Zhifei Longcom Biopharmaceutical produced the ZF2001 vaccine, a dimeric RBD domain with aluminum hydroxide, which was immunogenic and well-tolerated in a phase 1/2 clinical trial and is being evaluated in a phase 3 clinical trial (NCT04646590) [51]. Medicago Inc. has developed CoVLP, a ‘virus-like particle’ (VLP) comprising empty virus particles containing SARS-CoV-2 S viral surface antigens, which is being evaluated in a phase 2/3 clinical trial (NCT04636697). Medicago also manufactured a plant-based quadrivalent virus-like particle (QVLP) that was successful against influenza infection (Figure 2B) [64].
Viral vectors including adenoviruses, modified vaccinia Ankara (MVA), and vesicular stomatitis virus (VSV) have been used successfully to produce vaccines against pathogens such as the Ebola virus [1]. However, the host may already have immunity against some viral vectors, which must be considered in determining the prime and boost doses. CanSino Biologics made a recombinant adenovirus type-5 (Ad5) vector vaccine expressing the S protein, Ad5-nCoV, that was fully immunogenic, and a single dose induced a significant immune response [49,65]. Sputnik V (Gam-COVID-Vac) is a combination vector vaccine of rAd26 and rAd5 containing the gene for the S protein, which had an efficacy rate of 91.6% in a preliminary report of a phase 3 clinical trial [49].
Ad5 is the most common adenovirus type that infects humans, and many people might have contracted the virus and may be immune to it; however, Ad26 is less common [1]. The Ad26.COV2.S vector containing the gene for the S protein, developed by Janssen, produced a durable immune response. A single dose of Ad26.COV2.S induced a humoral immune response in more than 90% of participants [66]. Ad26.COV2.S is being evaluated in a phase 2/3 clinical trial (NCT04614948). In phase 3 trial, single shot of Ad26.COV2.S was safe and effective [49]. Ad26.COV2.S is currently in use as the “J&J/Janssen” vaccine. To avoid pre-existing immunity, AstraZeneca and Oxford University used a chimpanzee adenovirus to deliver the gene for the S protein (ChAdOx1; AZD1222). AZD1222 was immunogenic and well-tolerated; however, older people showed greater tolerance than younger people in a phase 2/3 clinical trial [67]. A single dose of AZD1222 fully induced the T cell response [68], but after a second dose, the antibody response was stronger [69]. AZD1222 is currently in use as the “AstraZeneca” vaccine (Figure 2C).
The COVID-19 pandemic has led to the rapid development of DNA and RNA vaccines, which are currently being tested and used. Antigen-coding DNA is delivered intradermally or intramuscularly where local myocytes or nascent cells take up the DNA and synthesize the antigen. The antigen is taken up by antigen-presenting cells and then presented to adaptive immune cells. For effective delivery, DNA vaccines need to be electroporated into cells [1]. INO-4800, which contains DNA coding for the S protein of SARS-CoV-2 has been developed by Inovio and International Vaccine Institute. INO-4800 is delivered intradermally and a small electric pulse is applied to the skin leading to cellular uptake of the DNA. The DNA vaccine was immunogenic and well-tolerated in 100% of vaccinated subjects in a phase 1 clinical trial [52]. INO-4800 is being evaluated in an ongoing phase 2/3 clinical trial (INNOVATE; NCT04642638) (Figure 2C) [1].
RNA vaccines have been at the forefront of rapid vaccine development due to COVID-19. Pfizer and BioNTech produced BNT162b2, a vaccine formulated using mRNA for the S protein encased in lipid nanoparticles. Two doses of BNT162b2 showed 95% protection in a phase 2 clinical trial [53]. Although the phase 3 clinical trial (NCT04713553) is still recruiting, BNT162b2 was the first mRNA COVID-19 vaccine approved for use [70]. BNT162b2 is currently in use as the “Pfizer/BioNTech” COVID-19 vaccine. Interestingly, 12–37 days after the first dose of the vaccine, the viral loads of patients were significantly decreased [71]. The second mRNA vaccine is mRNA-1273, developed by Moderna and NIAID, which also contains mRNA coding for the S protein inside lipid nanoparticles. A preliminary report of a phase 2 trial showed that mRNA-1273 induced an immune response against SARS-CoV-2 [72]. Result of phase 3 clinical trial of mRNA-1273 showed 94.1% efficacy at preventing COVID-19 disease [50]. After a second dose of mRNA-1273, antibodies persisted for 6 months [73]. mRNA-1273 is currently in use as the “Moderna” vaccine (Figure 2C).
Several other vaccines are under development or still in the preclinical trial or clinical trial stage of development. Several vaccines are already approved and in current use.
4. Delivery Route of Vaccines
Most vaccines are injected intramuscularly [1] because the traditional subcutaneous route for vaccines that include an aluminum salt adjuvant resulted in severe adverse effects [74]. A clinical trial for a diphtheria toxin (DT) vaccine with a booster shot demonstrated that an intramuscular vaccination gave significantly fewer adverse effects than a subcutaneous injection [75]. Intramuscular injections of the H3N2 and H1N1 influenza vaccines were more immunogenic than a subcutaneous injection [76]. One vaccine study in rhesus macaques using the HIV-1 envelope glycoprotein (Env) in synthetic liposomes gave comparable immune responses for intramuscular and subcutaneous delivery [77]. The authors of a systematic review of delivery methods recommended intradermal rather than subcutaneous or intramuscular injection, when appropriate, because an intradermal injection requires a lower dose of the vaccine [78].
The route of immunization may induce different mechanisms of protection. An aerosol vaccination of the DNA prime-Ad5 booster vaccine for Simian immunodeficiency virus (SIV) antigens induced IgA-driven neutrophil-mediated phagocytosis, while intramuscular injection induced IgG-driven antibody-dependent monocyte-mediated phagocytosis [79]. However, these varied results came from different vaccine platforms suggesting that the efficacy of the vaccination route might be related to the platform technology, such as mRNA, DNA, or an adenoviral vector vaccine.
The discovery of tissue-resident memory T cells (Trm) showed that memory T cells residing in nonlymphoid tissue can respond immediately to pathogens [80,81]. The prime and pull strategy, which comprises a conventional parenteral vaccination and recruitment of activated T cells to the local area, establishes long-lasting local immune memory and effective protection against vaginal herpes simplex virus type 2 (HSV-2) infection [82]. Enhanced Trm accumulation requires repeated local antigen recognition [83]. Although circulating IgG can protect the host from mucosal infection, local IgA from plasma cells inhibits infection of local target cells. After influenza infection, local plasma cells secrete IgA to protect type I mucosal cells of the upper respiratory tract while IgG protects lung cells [84]. CD4 Trm cells recruit circulating memory B cells to secrete IgG and IgA in the type II mucosa [85]. These results suggest that local vaccination might be more effective than peripheral vaccination for mucosal infection.
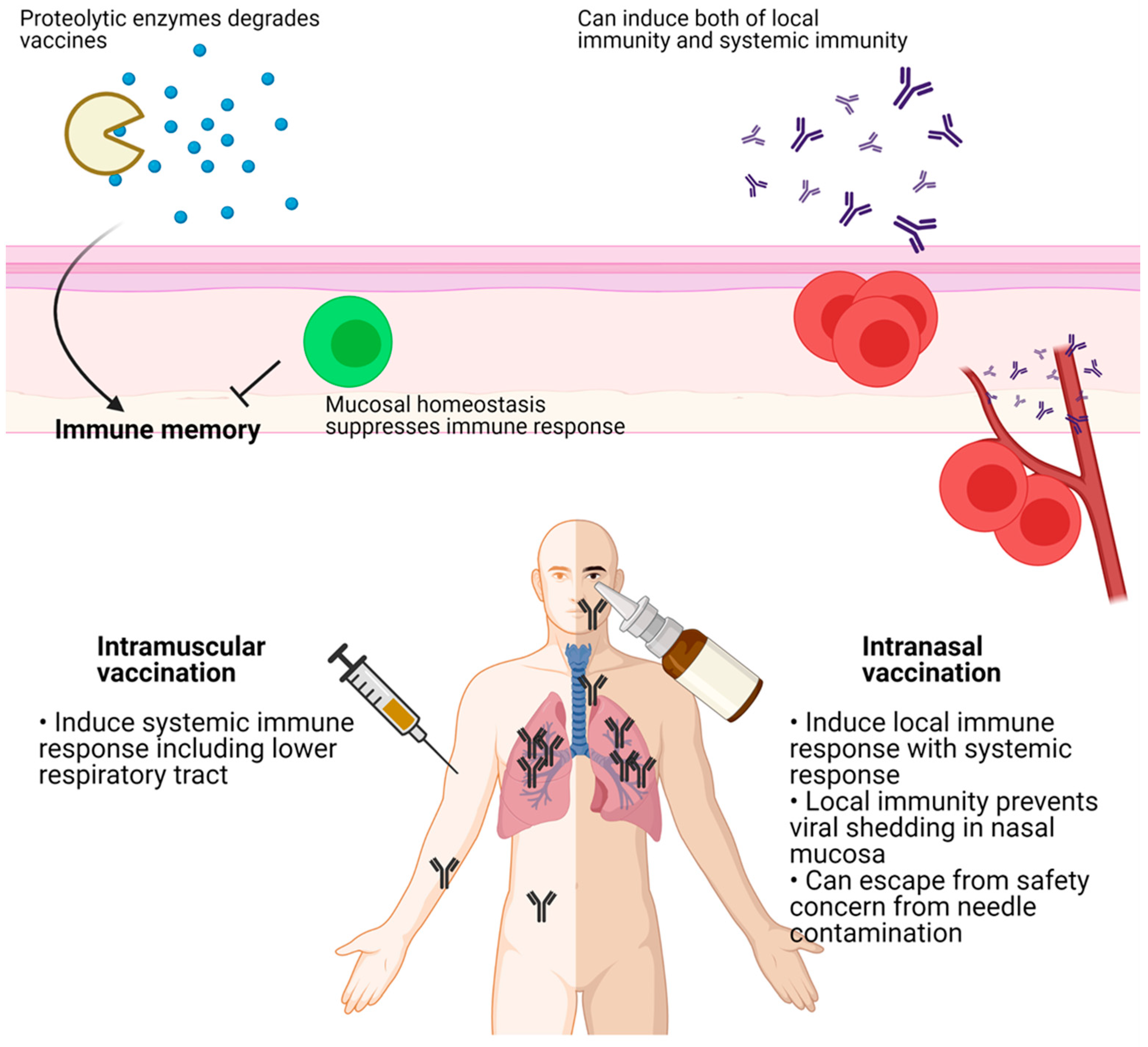
In addition to efficacy, mucosal vaccination is safer than peripheral vaccination because it avoids possible blood-borne infections from contaminated needles. While mucosal vaccination can induce a systemic immune response [86], the development of mucosal vaccines is difficult because little is known about mucosal immunity. The mucosal vaccine response could be attenuated since the mucosa maintains homeostasis in response to foreign invasion, and absorption of antigens through the mucosal barrier might be difficult as the antigens could be destroyed by proteolytic enzymes (Figure 3) [87]. Additionally, many vaccine platforms such as subunit and inactivated virus vaccines need adjuvants for an effective immune response. However, common adjuvants, including alum, could not induce the IgA and IgG class-switching and the recruitment of T and B cells to the mucosal area. Thus, infectious viral vectors that can trigger pathogen-associated molecular patterns (PAMPs) might be more suitable for mucosal vaccines [88].
5. Intranasal Vaccination of COVID-19 Vaccines
Currently, all vaccines for COVID-19 are injected intramuscularly. However, because SARS-CoV-2 mainly infects via the upper respiratory tract, the microenvironment of the nasal passage is important for immunity. For example, intranasal immunization of mice using the N protein resulted in a higher T cell response in lung bronchoalveolar lavage (BAL) than did intramuscular injection, while responses in the trachea and the oral cavity were similar. In contrast, T cell responses in the spleen, inguinal lymph nodes, and the brain were lower with intranasal immunization than with intramuscular injection [89]. This suggests that the route of immunization affects the immune responses of multiple organs, not only the target organ.
Several experiments using intranasal vaccinations have given generally favorable results. Intranasal vaccination using SARS-CoV induced Trm cells in the lungs while subcutaneous injection established memory T cells in the spleen, and airway Trm cells were important for protection from SARS-CoV infection [90]. Intramuscular ChAdOx1 injection in rhesus macaques reduced viral load in the bronchoalveolar lavage fluid (BALF) and prevented pneumonia, but did not prevent nasal shedding of the virus [3]. Intranasal ChAd-SARS-CoV-2-S (based on the simian Ad-36 vector) shows superior protection and mucosal immunity compared to the parenteral route in a murine model [91] and prevented viral shedding in rhesus macaques [4]. Intranasal and subcutaneous Ad5.SARS-CoV-2-S1 vaccination induced robust humoral and cellular immune responses in a murine model [92]. Administration of Ad5-S-nb2 in the nasal cavity also induced a systemic and a local antibody response that protected macaques [93]. Local administration of a lentivirus vector induced an IgA response that protected mice from SARS-CoV-2 [94]. Intranasal immunization with inactivated SARS-CoV with adjuvants was sufficient to induce local and serum antibodies in mice [95]. Similarly, nasal administration of a recombinant RBD vaccine with alum induced protective immunity against SARS-CoV-2 in mice [96]. These results suggest that intranasal administration of vaccines protects the host from SARS-CoV-2 infection and that local vaccination was sufficient to induce systemic humoral immune responses.
However, other experiments gave mixed results from intranasal vaccinations. Intranasal injection of human neutralizing antibodies (HuNAbs) and a DNA vaccine did not reduce the viral load in the nasal turbinate, but the lung was protected [97]. Intramuscular delivery of VSV-SARS2-EBOV vaccine demonstrated rapid protection compared to an intranasal injection [98]. However, these results contradict previous studies. While it is not yet clear whether intranasal vaccination is superior to intramuscular vaccination, it is evident that intranasal vaccination protects the host from infection. The efficacy of an intranasal vaccination may depend on the dosage or the vaccine platform. However, because nasal immunization can provide sterilizing immunity, preventing interhuman transmission, it might be an effective route to herd immunity [5]. While several clinical trials of intranasal vaccines, including AdCOVID (altimmune; Gaitersburg, USA), are currently in progress [1], further studies are needed to determine the most effective route for immunization.
6. Conclusions
The rapid development of vaccine technology has led to several vaccines for COVID-19. Some of the vaccines that have been approved for emergency use include mRNA-1273 (Moderna), AZD1222 (AstraZeneca), and Ad26.COV2.S (J&J/Janssen), which are delivered intramuscularly, and the results of their global use will provide important information on their safety and efficacy. As the local immune microenvironment is important for protection against infection and for preventing viral shedding, intranasal immunization is expected to be more effective than intramuscular immunization. While research has shown that in some cases intranasal immunization is better than an intramuscular injection, there are also contradictory results. Depending on the vaccine platform, the efficacy of intranasal immunization may differ. For example, a live viral vector may be more effective in inducing an immune response because it does not require an adjuvant. Since we do not fully understand mucosal immunity, future studies should focus on how the vaccine platform affects the mucosal immune response.
Author Contributions
Conceptualization: J.H.P. and H.K.L.; writing—original draft preparation: J.H.P. and H.K.L.; writing—review and editing: J.H.P. and H.K.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Mobile Clinic Module Project funded by KAIST and the 2020 Joint Research Project of Institutes of Science and Technology. This work was also supported by National Research Foundation of Korea (NRF-2021M3A9D3026428).
Institutional Review Board Statement
This review was completed with no requirement for Institutional Review.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data was generated.
Acknowledgments
The authors would like to thank the members of the Laboratory of Host Defenses for helpful advice.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kyriakidis, N.C.; Lopez-Cortes, A.; Gonzalez, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Hikmet, F.; Mear, L.; Edvinsson, A.; Micke, P.; Uhlen, M.; Lindskog, C. The protein expression profile of ACE2 in human tissues. Mol. Syst. Biol. 2020, 16, e9610. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Lambe, T.; Spencer, A.; Belij-Rammerstorfer, S.; Purushotham, J.N.; Port, J.R.; Avanzato, V.A.; Bushmaker, T.; Flaxman, A.; Ulaszewska, M.; et al. ChAdOx1 nCoV-19 vaccine prevents SARS-CoV-2 pneumonia in rhesus macaques. Nature 2020, 586, 578–582. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Purushotham, J.; Schulz, J.; Holbrook, M.; Bushmaker, T.; Carmody, A.; Port, J.; Yinda, K.C.; Okumura, A.; Saturday, G.; et al. Intranasal ChAdOx1 nCoV-19/AZD1222 vaccination reduces shedding of SARS-CoV-2 D614G in rhesus macaques. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gallo, O.; Locatello, L.G.; Mazzoni, A.; Novelli, L.; Annunziato, F. The central role of the nasal microenvironment in the transmission, modulation, and clinical progression of SARS-CoV-2 infection. Mucosal. Immunol. 2021, 14, 305–316. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the human innate immune system. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef] [PubMed]
- Salamanna, F.; Maglio, M.; Landini, M.P.; Fini, M. Body localization of ACE-2: On the trail of the keyhole of SARS-CoV-2. Front. Med. 2020, 7, 594495. [Google Scholar] [CrossRef] [PubMed]
- Bost, P.; Giladi, A.; Liu, Y.; Bendjelal, Y.; Xu, G.; David, E.; Blecher-Gonen, R.; Cohen, M.; Medaglia, C.; Li, H.; et al. Host-viral infection maps reveal signatures of severe COVID-19 patients. Cell 2020, 181, 1475–1488.e12. [Google Scholar] [CrossRef] [PubMed]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Blanco, M.R.; Bruce, E.A.; Honson, D.D.; Chen, L.M.; Chow, A.; Bhat, P.; Ollikainen, N.; Quinodoz, S.A.; Loney, C.; et al. SARS-CoV-2 disrupts splicing, translation, and protein trafficking to suppress host defenses. Cell 2020, 183, 1325–1339.e21. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 2020, 181, 1036–1045.e9. [Google Scholar] [CrossRef] [PubMed]
- Mulchandani, R.; Lyngdoh, T.; Kakkar, A.K. Deciphering the COVID-19 cytokine storm: Systematic review and meta-analysis. Eur. J. Clin. Invest. 2021, 51, e13429. [Google Scholar] [CrossRef] [PubMed]
- Chua, R.L.; Lukassen, S.; Trump, S.; Hennig, B.P.; Wendisch, D.; Pott, F.; Debnath, O.; Thurmann, L.; Kurth, F.; Volker, M.T.; et al. COVID-19 severity correlates with airway epithelium-immune cell interactions identified by single-cell analysis. Nat. Biotechnol. 2020, 38, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, H.K. Re-analysis of single cell transcriptome reveals that the NR3C1-CXCL8-neutrophil axis determines the severity of COVID-19. Front. Immunol. 2020, 11, 2145. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Bassler, K.; Schlickeiser, S.; Zhang, B.; Kramer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 is marked by a dysregulated myeloid cell compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Flament, H.; Rouland, M.; Beaudoin, L.; Toubal, A.; Bertrand, L.; Lebourgeois, S.; Rousseau, C.; Soulard, P.; Gouda, Z.; Cagninacci, L.; et al. Outcome of SARS-CoV-2 infection is linked to MAIT cell activation and cytotoxicity. Nat. Immunol. 2021, 22, 322–335. [Google Scholar] [CrossRef]
- Lei, L.; Qian, H.; Yang, X.; Zhang, X.; Zhang, D.; Dai, T.; Guo, R.; Shi, L.; Cheng, Y.; Zhang, B.; et al. The phenotypic changes of gammadelta T cells in COVID-19 patients. J. Cell. Mol. Med. 2020, 24, 11603–11606. [Google Scholar] [CrossRef]
- Chen, Z.; Wherry, E.J. T cell responses in patients with COVID-19. Nat. Rev. Immunol. 2020, 20, 529–536. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex immune dysregulation in COVID-19 patients with severe respiratory failure. Cell Host Microbe 2020, 27, 992–1000.e3. [Google Scholar] [CrossRef]
- Mazzoni, A.; Salvati, L.; Maggi, L.; Capone, M.; Vanni, A.; Spinicci, M.; Mencarini, J.; Caporale, R.; Peruzzi, B.; Antonelli, A.; et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J. Clin. Invest. 2020, 130, 4694–4703. [Google Scholar] [CrossRef]
- McClain, M.T.; Park, L.P.; Nicholson, B.; Veldman, T.; Zaas, A.K.; Turner, R.; Lambkin-Williams, R.; Gilbert, A.S.; Ginsburg, G.S.; Woods, C.W. Longitudinal analysis of leukocyte differentials in peripheral blood of patients with acute respiratory viral infections. J. Clin. Virol. 2013, 58, 689–695. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’Andrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369. [Google Scholar] [CrossRef]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Stralin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef] [PubMed]
- Rha, M.S.; Jeong, H.W.; Ko, J.H.; Choi, S.J.; Seo, I.H.; Lee, J.S.; Sa, M.; Kim, A.R.; Joo, E.J.; Ahn, J.Y.; et al. PD-1-Expressing SARS-CoV-2-specific CD8(+) T cells are not exhausted, but functional in patients with COVID-19. Immunity 2021, 54, 44–52.e3. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Invest. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Juno, J.A.; Tan, H.X.; Lee, W.S.; Reynaldi, A.; Kelly, H.G.; Wragg, K.; Esterbauer, R.; Kent, H.E.; Batten, C.J.; Mordant, F.L.; et al. Humoral and circulating follicular helper T cell responses in recovered patients with COVID-19. Nat. Med. 2020, 26, 1428–1434. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Edridge, A.W.D.; Kaczorowska, J.; Hoste, A.C.R.; Bakker, M.; Klein, M.; Loens, K.; Jebbink, M.F.; Matser, A.; Kinsella, C.M.; Rueda, P.; et al. Seasonal coronavirus protective immunity is short-lasting. Nat. Med. 2020, 26, 1691–1693. [Google Scholar] [CrossRef]
- Kim, C.W.; Oh, J.E.; Lee, H.K. Single cell transcriptomic re-analysis of immune cells in bronchoalveolar lavage fluids reveals the correlation of B cell characteristics and disease severity of patients with SARS-CoV-2 infection. Immune Netw. 2021, 21, e10. [Google Scholar] [CrossRef]
- Liu, W.; Fontanet, A.; Zhang, P.H.; Zhan, L.; Xin, Z.T.; Baril, L.; Tang, F.; Lv, H.; Cao, W.C. Two-year prospective study of the humoral immune response of patients with severe acute respiratory syndrome. J. Infect. Dis. 2006, 193, 792–795. [Google Scholar] [CrossRef] [Green Version]
- Wajnberg, A.; Amanat, F.; Firpo, A.; Altman, D.R.; Bailey, M.J.; Mansour, M.; McMahon, M.; Meade, P.; Mendu, D.R.; Muellers, K.; et al. Robust neutralizing antibodies to SARS-CoV-2 infection persist for months. Science 2020, 370, 1227–1230. [Google Scholar] [CrossRef]
- Long, Q.X.; Tang, X.J.; Shi, Q.L.; Li, Q.; Deng, H.J.; Yuan, J.; Hu, J.L.; Xu, W.; Zhang, Y.; Lv, F.J.; et al. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nat. Med. 2020, 26, 1200–1204. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, S.; Lee, Y.; Grubbs, G.; Coyle, E.M.; Klenow, L.; Akasaka, O.; Koga, M.; Adachi, E.; Saito, M.; Nakachi, I.; et al. Longitudinal antibody repertoire in “mild” versus “severe” COVID-19 patients reveals immune markers associated with disease severity and resolution. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Yu, H.Q.; Sun, B.Q.; Fang, Z.F.; Zhao, J.C.; Liu, X.Y.; Li, Y.M.; Sun, X.Z.; Liang, H.F.; Zhong, B.; Huang, Z.F.; et al. Distinct features of SARS-CoV-2-specific IgA response in COVID-19 patients. Eur. Respir. J. 2020, 56. [Google Scholar] [CrossRef] [PubMed]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claer, L.; Quentric, P.; Fadlallah, J.; Devilliers, H.; Ghillani, P.; et al. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Sci. Transl. Med. 2021, 13. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Yang, Y. The potential danger of suboptimal antibody responses in COVID-19. Nat. Rev. Immunol. 2020, 20, 339–341. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Hu, Y.; Xu, M.; Chen, Z.; Yang, W.; Jiang, Z.; Li, M.; Jin, H.; Cui, G.; Chen, P.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine (CoronaVac) in healthy adults aged 60 years and older: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, Y.; Wang, Y.; Wang, H.; Yang, Y.; Gao, G.F.; Tan, W.; Wu, G.; Xu, M.; Lou, Z.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: A randomised, double-blind, placebo-controlled, phase 1/2 trial. Lancet Infect. Dis. 2021, 21, 39–51. [Google Scholar] [CrossRef]
- Ella, R.; Reddy, S.; Jogdand, H.; Sarangi, V.; Ganneru, B.; Prasad, S.; Das, D.; Raju, D.; Praturi, U.; Sapkal, G.; et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBV152: Interim results from a double-blind, randomised, multicentre, phase 2 trial, and 3-month follow-up of a double-blind, randomised phase 1 trial. Lancet Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1–2 trial of a SARS-CoV-2 recombinant spike protein nanoparticle vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S.; et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cardenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and efficacy of single-dose Ad26.COV2.S vaccine against Covid-19. N. Engl. J. Med. 2021. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Tebas, P.; Yang, S.; Boyer, J.D.; Reuschel, E.L.; Patel, A.; Christensen-Quick, A.; Andrade, V.M.; Morrow, M.P.; Kraynyak, K.; Agnes, J.; et al. Safety and immunogenicity of INO-4800 DNA vaccine against SARS-CoV-2: A preliminary report of an open-label, phase 1 clinical trial. EClinicalMedicine 2021, 31, 100689. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- De Assis, R.R.; Jain, A.; Nakajima, R.; Jasinskas, A.; Felgner, J.; Obiero, J.M.; Norris, P.J.; Stone, M.; Simmons, G.; Bagri, A.; et al. Analysis of SARS-CoV-2 antibodies in COVID-19 convalescent blood using a coronavirus antigen microarray. Nat. Commun. 2021, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Rota, P.A.; Moss, W.J.; Takeda, M.; de Swart, R.L.; Thompson, K.M.; Goodson, J.L. Measles. Nat. Rev. Dis. Primers 2016, 2, 16049. [Google Scholar] [CrossRef] [PubMed]
- Okamura, S.; Kashiwabara, A.; Suzuki, H.; Ueno, S.; Miyazato, P.; Takekawa, S.; Kamitani, W.; Yamanishi, K.; Ebina, H. Live attenuated SARS-CoV-2 vaccine candidate: Protective immunity without serious lung lesions in Syrian hamsters. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18–59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef]
- Palacios, R.; Patino, E.G.; de Oliveira, R.P.; Conde, M.; Batista, A.P.; Zeng, G.; Xin, Q.; Kallas, E.G.; Flores, J.; Ockenhouse, C.F.; et al. Double-blind, randomized, placebo-controlled phase III clinical trial to evaluate the efficacy and safety of treating healthcare professionals with the adsorbed COVID-19 (inactivated) vaccine manufactured by Sinovac—PROFISCOV: A structured summary of a study protocol for a randomised controlled trial. Trials 2020, 21, 853. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, Y.; Huang, B.; Deng, W.; Quan, Y.; Wang, W.; Xu, W.; Zhao, Y.; Li, N.; Zhang, J.; et al. Development of an inactivated vaccine candidate, BBIBP-CorV, with potent protection against SARS-CoV-2. Cell 2020, 182, 713–721.e9. [Google Scholar] [CrossRef]
- Ganneru, B.; Jogdand, H.; Daram, V.K.; Das, D.; Molugu, N.R.; Prasad, S.D.; Kannappa, S.V.; Ella, K.M.; Ravikrishnan, R.; Awasthi, A.; et al. Th1 skewed immune response of whole virion inactivated SARS CoV 2 vaccine and its safety evaluation. iScience 2021, 24, 102298. [Google Scholar] [CrossRef]
- Yadav, P.D.; Ella, R.; Kumar, S.; Patil, D.R.; Mohandas, S.; Shete, A.M.; Vadrevu, K.M.; Bhati, G.; Sapkal, G.; Kaushal, H.; et al. Immunogenicity and protective efficacy of inactivated SARS-CoV-2 vaccine candidate, BBV152 in rhesus macaques. Nat. Commun. 2021, 12, 1386. [Google Scholar] [CrossRef]
- Mohandas, S.; Yadav, P.D.; Shete-Aich, A.; Abraham, P.; Vadrevu, K.M.; Sapkal, G.; Mote, C.; Nyayanit, D.; Gupta, N.; Srinivas, V.K.; et al. Immunogenicity and protective efficacy of BBV152, whole virion inactivated SARS-CoV-2 vaccine candidates in the Syrian hamster model. iScience 2021, 24, 102054. [Google Scholar] [CrossRef]
- Guebre-Xabier, M.; Patel, N.; Tian, J.H.; Zhou, B.; Maciejewski, S.; Lam, K.; Portnoff, A.D.; Massare, M.J.; Frieman, M.B.; Piedra, P.A.; et al. NVX-CoV2373 vaccine protects cynomolgus macaque upper and lower airways against SARS-CoV-2 challenge. Vaccine 2020, 38, 7892–7896. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.H.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.D.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Ward, B.J.; Makarkov, A.; Seguin, A.; Pillet, S.; Trepanier, S.; Dhaliwall, J.; Libman, M.D.; Vesikari, T.; Landry, N. Efficacy, immunogenicity, and safety of a plant-derived, quadrivalent, virus-like particle influenza vaccine in adults (18–64 years) and older adults (>/=65 years): Two multicentre, randomised phase 3 trials. Lancet 2020, 396, 1491–1503. [Google Scholar] [CrossRef]
- Zhu, F.C.; Li, Y.H.; Guan, X.H.; Hou, L.H.; Wang, W.J.; Li, J.X.; Wu, S.P.; Wang, B.S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef]
- Sadoff, J.; Le Gars, M.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; van Damme, W.; Leroux-Roels, I.; et al. Interim results of a phase 1-2a trial of Ad26.COV2.S Covid-19 vaccine. N. Engl. J. Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, M.N.; Minassian, A.M.; Ewer, K.J.; Flaxman, A.L.; Folegatti, P.M.; Owens, D.R.; Voysey, M.; Aley, P.K.; Angus, B.; Babbage, G.; et al. Safety and immunogenicity of ChAdOx1 nCoV-19 vaccine administered in a prime-boost regimen in young and old adults (COV002): A single-blind, randomised, controlled, phase 2/3 trial. Lancet 2021, 396, 1979–1993. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Barrett, J.R.; Belij-Rammerstorfer, S.; Dold, C.; Ewer, K.J.; Folegatti, P.M.; Gilbride, C.; Halkerston, R.; Hill, J.; Jenkin, D.; Stockdale, L.; et al. Phase 1/2 trial of SARS-CoV-2 vaccine ChAdOx1 nCoV-19 with a booster dose induces multifunctional antibody responses. Nat. Med. 2021, 27, 279–288. [Google Scholar] [CrossRef]
- Lamb, Y.N. BNT162b2 mRNA COVID-19 vaccine: First approval. Drugs 2021, 81, 495–501. [Google Scholar] [CrossRef]
- Levine-Tiefenbrun, M.; Yelin, I.; Katz, R.; Herzel, E.; Golan, Z.; Schreiber, L.; Wolf, T.; Nadler, V.; Ben-Tov, A.; Kuint, J.; et al. Initial report of decreased SARS-CoV-2 viral load after inoculation with the BNT162b2 vaccine. Nat. Med. 2021. [Google Scholar] [CrossRef]
- Chu, L.; McPhee, R.; Huang, W.; Bennett, H.; Pajon, R.; Nestorova, B.; Leav, B.; mRNA-1273 Study Group. A preliminary report of a randomized controlled phase 2 trial of the safety and immunogenicity of mRNA-1273 SARS-CoV-2 vaccine. Vaccine 2021. [Google Scholar] [CrossRef] [PubMed]
- Doria-Rose, N.; Suthar, M.S.; Makowski, M.; O’Connell, S.; McDermott, A.B.; Flach, B.; Ledgerwood, J.E.; Mascola, J.R.; Graham, B.S.; Lin, B.C.; et al. Antibody persistence through 6 months after the second dose of mRNA-1273 vaccine for Covid-19. N. Engl. J. Med. 2021. [Google Scholar] [CrossRef]
- Cook, I.F. Evidence based route of administration of vaccines. Hum. Vaccines 2008, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Mark, A.; Carlsson, R.M.; Granstrom, M. Subcutaneous versus intramuscular injection for booster DT vaccination of adolescents. Vaccine 1999, 17, 2067–2072. [Google Scholar] [CrossRef]
- Cook, I.F.; Barr, I.; Hartel, G.; Pond, D.; Hampson, A.W. Reactogenicity and immunogenicity of an inactivated influenza vaccine administered by intramuscular or subcutaneous injection in elderly adults. Vaccine 2006, 24, 2395–2402. [Google Scholar] [CrossRef]
- Ols, S.; Yang, L.; Thompson, E.A.; Pushparaj, P.; Tran, K.; Liang, F.; Lin, A.; Eriksson, B.; Hedestam, G.B.K.; Wyatt, R.T.; et al. Route of vaccine administration alters antigen trafficking but not innate or adaptive immunity. Cell Rep. 2020, 30, 3964–3971.e7. [Google Scholar] [CrossRef]
- Schnyder, J.L.; de Pijper, C.A.; Garrido, H.M.G.; Daams, J.G.; Goorhuis, A.; Stijnis, C.; Schaumburg, F.; Grobusch, M.P. Fractional dose of intradermal compared to intramuscular and subcutaneous vaccination—A systematic review and meta-analysis. Travel Med. Infect. Dis. 2020, 37, 101868. [Google Scholar] [CrossRef]
- Ackerman, M.E.; Das, J.; Pittala, S.; Broge, T.; Linde, C.; Suscovich, T.J.; Brown, E.P.; Bradley, T.; Natarajan, H.; Lin, S.; et al. Route of immunization defines multiple mechanisms of vaccine-mediated protection against SIV. Nat. Med. 2018, 24, 1590–1598. [Google Scholar] [CrossRef] [PubMed]
- Wakim, L.M.; Waithman, J.; van Rooijen, N.; Heath, W.R.; Carbone, F.R. Dendritic cell-induced memory T cell activation in nonlymphoid tissues. Science 2008, 319, 198–202. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, T.; Wakim, L.M.; Eidsmo, L.; Reading, P.C.; Heath, W.R.; Carbone, F.R. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat. Immunol. 2009, 10, 524–530. [Google Scholar] [CrossRef]
- Shin, H.; Iwasaki, A. A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature 2012, 491, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Davies, B.; Prier, J.E.; Jones, C.M.; Gebhardt, T.; Carbone, F.R.; Mackay, L.K. Cutting edge: Tissue-resident memory T cells generated by multiple immunizations or localized deposition provide enhanced immunity. J. Immunol. 2017, 198, 2233–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renegar, K.B.; Small, P.A., Jr.; Boykins, L.G.; Wright, P.F. Role of IgA versus IgG in the control of influenza viral infection in the murine respiratory tract. J. Immunol. 2004, 173, 1978–1986. [Google Scholar] [CrossRef]
- Oh, J.E.; Iijima, N.; Song, E.; Lu, P.; Klein, J.; Jiang, R.; Kleinstein, S.H.; Iwasaki, A. Migrant memory B cells secrete luminal antibody in the vagina. Nature 2019, 571, 122–126. [Google Scholar] [CrossRef]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Mestecky, J.; Russell, M.W.; Elson, C.O. Perspectives on mucosal vaccines: Is mucosal tolerance a barrier? J. Immunol. 2007, 179, 5633–5638. [Google Scholar] [CrossRef] [Green Version]
- Boyaka, P.N. Inducing mucosal IgA: A challenge for vaccine adjuvants and delivery systems. J. Immunol. 2017, 199, 9–16. [Google Scholar] [CrossRef]
- Joag, V.; Wijeyesinghe, S.; Stolley, J.M.; Quarnstrom, C.F.; Dileepan, T.; Soerens, A.G.; Sangala, J.A.; O’Flanagan, S.D.; Gavil, N.V.; Hong, S.W.; et al. Cutting edge: Mouse SARS-CoV-2 epitope reveals infection and vaccine-elicited CD8 T cell responses. J. Immunol. 2021, 206, 931–935. [Google Scholar] [CrossRef]
- Zhao, J.; Zhao, J.; Mangalam, A.K.; Channappanavar, R.; Fett, C.; Meyerholz, D.K.; Agnihothram, S.; Baric, R.S.; David, C.S.; Perlman, S. Airway memory CD4(+) T cells mediate protective immunity against emerging respiratory coronaviruses. Immunity 2016, 44, 1379–1391. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.O.; Kafai, N.M.; Dmitriev, I.P.; Fox, J.M.; Smith, B.K.; Harvey, I.B.; Chen, R.E.; Winkler, E.S.; Wessel, A.W.; Case, J.B.; et al. A single-dose intranasal ChAd vaccine protects upper and lower respiratory tracts against SARS-CoV-2. Cell 2020, 183, 169–184.e13. [Google Scholar] [CrossRef]
- Kim, E.; Weisel, F.J.; Balmert, S.C.; Khan, M.S.; Huang, S.; Erdos, G.; Kenniston, T.W.; Carey, C.D.; Joachim, S.M.; Conter, L.J.; et al. A single subcutaneous or intranasal immunization with Adenovirus-based SARS-CoV-2 vaccine induces robust humoral and cellular immune responses in mice. Eur. J. Immunol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Wang, Q.; Shan, C.; Yang, C.; Feng, Y.; Wu, J.; Liu, X.; Zhou, Y.; Jiang, R.; Hu, P.; et al. An adenovirus-vectored COVID-19 vaccine confers protection from SARS-COV-2 challenge in rhesus macaques. Nat. Commun. 2020, 11, 4207. [Google Scholar] [CrossRef]
- Ku, M.W.; Bourgine, M.; Authie, P.; Lopez, J.; Nemirov, K.; Moncoq, F.; Noirat, A.; Vesin, B.; Nevo, F.; Blanc, C.; et al. Intranasal vaccination with a lentiviral vector protects against SARS-CoV-2 in preclinical animal models. Cell Host Microbe 2021, 29, 236–249.e6. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Zheng, B.; Yao, X.; Guan, Y.; Yuan, Z.H.; Zhong, N.S.; Lu, L.W.; Xie, J.P.; Wen, Y.M. Intranasal immunization with inactivated SARS-CoV (SARS-associated coronavirus) induced local and serum antibodies in mice. Vaccine 2005, 23, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Xu, Y.; Feng, J.; Hu, L.; Zhang, Y.; Zhang, B.; Guo, W.; Mai, R.; Chen, L.; Fang, J.; et al. Intranasal administration of a recombinant RBD vaccine induced protective immunity against SARS-CoV-2 in mouse. Vaccine 2021. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Chan, J.F.; Zhou, B.; Zhou, R.; Li, S.; Shan, S.; Liu, L.; Zhang, A.J.; Chen, S.J.; Chan, C.C.; et al. Robust SARS-CoV-2 infection in nasal turbinates after treatment with systemic neutralizing antibodies. Cell Host Microbe 2021. [Google Scholar] [CrossRef]
- Furuyama, W.; Shifflett, K.; Pinski, A.N.; Griffin, A.J.; Feldmann, F.; Okumura, A.; Gourdine, T.; Jankeel, A.; Lovaglio, J.; Hanley, P.W.; et al. Rapid protection from COVID-19 in nonhuman primates vaccinated intramuscularly but not intranasally with a single dose of a recombinant vaccine. bioRxiv 2021. [Google Scholar] [CrossRef]
Figure 1.
Immune responses against SARS-CoV-2 infection. (A) SARS-CoV-2 can be sensed by pattern recognition receptors (PRRs) such as Toll like receptors and RIG-I like receptors. Viral sensing induces interferons and proinflammatory cytokines and chemokines. However, SARS-CoV-2 inhibits induction of interferons. Reduced interferons and excessive proinflammatory cytokines cause cytokine storm and severe inflammation. (B) Innate immune cells including monocyte, macrophage, and neutrophils are the main source of proinflammatory cytokines such as IL-6 and TNF. Cytokines further activate immune cells. Unconventional T cells (MAIT cells, γδ T cells) also participate in positive feedback loop of inflammation. (C) SARS-CoV-2 infection leads to lymphopenia. CD8 T cells from COVID-19 patients showed exhausted feature. Whether these features represent activation or exhaustion should be addressed. Furthermore, balance among CD4 T cell subsets is a critical factor for disease severity. (D) Antibodies from B cells neutralize virus entry. Mucosal IgA is rapidly produced and shows superior neutralizing capacity compared to systemic IgG while circulating IgA is correlated with severe disease symptoms. Circulating IgG systemically protects host from virus. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).
Figure 1.
Immune responses against SARS-CoV-2 infection. (A) SARS-CoV-2 can be sensed by pattern recognition receptors (PRRs) such as Toll like receptors and RIG-I like receptors. Viral sensing induces interferons and proinflammatory cytokines and chemokines. However, SARS-CoV-2 inhibits induction of interferons. Reduced interferons and excessive proinflammatory cytokines cause cytokine storm and severe inflammation. (B) Innate immune cells including monocyte, macrophage, and neutrophils are the main source of proinflammatory cytokines such as IL-6 and TNF. Cytokines further activate immune cells. Unconventional T cells (MAIT cells, γδ T cells) also participate in positive feedback loop of inflammation. (C) SARS-CoV-2 infection leads to lymphopenia. CD8 T cells from COVID-19 patients showed exhausted feature. Whether these features represent activation or exhaustion should be addressed. Furthermore, balance among CD4 T cell subsets is a critical factor for disease severity. (D) Antibodies from B cells neutralize virus entry. Mucosal IgA is rapidly produced and shows superior neutralizing capacity compared to systemic IgG while circulating IgA is correlated with severe disease symptoms. Circulating IgG systemically protects host from virus. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).

Figure 2.
Vaccine platforms for COVID-19. (A) Live attenuated vaccines are developed through repeated attenuation. Inactivated vaccines are generated by chemical or heat stress. Live attenuated vaccines do not require adjuvant. However, this technology takes a long time and has safety concerns. Inactivated vaccines have reduced danger compared to attenuated vaccines. However, they need adjuvant because they lose their immunogenicity. (B) Protein subunit vaccine or subunit-containing virus like particle (VLP) provides antigen to antigen presenting cells (APCs). APCs present antigen to immune cells and generate immune memory. Although these platforms are safe, they need adjuvant. (C) Viral vector vaccines deliver antigen-coding genes to target cells. Infected cells make and give antigen to APCs. Viral vectors do not need adjuvant because the vector itself is danger-associated molecular pattern (DAMP). Delivery is also easy. However, they have safety concerns and vectors can be eradicated by host immune memory if the host has experience of the viral vector. mRNA vaccine and DNA vaccine deliver mRNA and DNA to the target cells. mRNA and DNA generate antigens. While mRNA vaccines are safe, fast and cheap, they need “cold chain” because RNA is unstable. DNA vaccines are stable and safe. However, they are weak to induce proper immunity. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).
Figure 2.
Vaccine platforms for COVID-19. (A) Live attenuated vaccines are developed through repeated attenuation. Inactivated vaccines are generated by chemical or heat stress. Live attenuated vaccines do not require adjuvant. However, this technology takes a long time and has safety concerns. Inactivated vaccines have reduced danger compared to attenuated vaccines. However, they need adjuvant because they lose their immunogenicity. (B) Protein subunit vaccine or subunit-containing virus like particle (VLP) provides antigen to antigen presenting cells (APCs). APCs present antigen to immune cells and generate immune memory. Although these platforms are safe, they need adjuvant. (C) Viral vector vaccines deliver antigen-coding genes to target cells. Infected cells make and give antigen to APCs. Viral vectors do not need adjuvant because the vector itself is danger-associated molecular pattern (DAMP). Delivery is also easy. However, they have safety concerns and vectors can be eradicated by host immune memory if the host has experience of the viral vector. mRNA vaccine and DNA vaccine deliver mRNA and DNA to the target cells. mRNA and DNA generate antigens. While mRNA vaccines are safe, fast and cheap, they need “cold chain” because RNA is unstable. DNA vaccines are stable and safe. However, they are weak to induce proper immunity. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).

Figure 3.
Intramuscular vaccination induces systemic immune response including lower respiratory tract (LRT), but not upper respiratory tract (URT). In contrary, intranasal vaccination induces local immune response in LRT with systemic immunity. Local immunity prevents nasal shedding of viruses which is critical for herd immunity. In the mucosal area, delivered antigens can be degraded by proteolytic enzymes. Mucosal homeostasis inhibits immune response against vaccines. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).
Figure 3.
Intramuscular vaccination induces systemic immune response including lower respiratory tract (LRT), but not upper respiratory tract (URT). In contrary, intranasal vaccination induces local immune response in LRT with systemic immunity. Local immunity prevents nasal shedding of viruses which is critical for herd immunity. In the mucosal area, delivered antigens can be degraded by proteolytic enzymes. Mucosal homeostasis inhibits immune response against vaccines. Figure was created by biorender.com (BioRender, Toronto, ON, Canada).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Current advances in COVID-19 vaccines.
| Name | Company | Route | Technology | Clinical Trial | Approved | Grade 3-4 Side Effects |
|---|---|---|---|---|---|---|
| COVI-VAC | Codagenix (NewYork, USA) Serum Institute (Pune, India) | Intranasal | Live-attenuated virus | Phase I clinical trial (NCT04619628) | No | |
| CoronaVac | Sinovac Biotech Ltd. (Beijing, China) | Intramuscular | Inactivated virus | Phase III clinical trial (NCT04800133, NCT04651790, NCT04456595, NCT04508075, NCT04582344, NCT04617483, PHRR210210-003308) | Yes | Wu et al. [43] phase 1/2 trial No |
| BBIBP-CorV | Sinopharm Beijing Institute of Biotechnology (Beijing, China) | Intramuscular | Inactivated virus | Phase III clinical trial (NCT04510207, ChiCTR2000034780, NCT04612972, NCT04560881) | Yes | Xia et al. [44] phase 1/2 trial, No |
| BBV152 (Covaxin) | Bharat Biotech (Hyderabad, India) | Intramuscular | Inactivated virus | Phase III clinical trial (NCT04641481) | Yes | Ella et al. [45]. phase 2 trial, days 0–7, 28–35 No |
| NVX-CoV2373 | Novavax (Gaithersburg, USA) | Intramuscular | Protein subunit | Phase III clinical trial (NCT04583995, NCT04611802) | No | Keech et al. [46]. phase 1–2 trial 2% (severe adverse events) for groups D and E |
| ZF2001 | Ahui Zhifei Longcom Biopharmaceutical (Hefei, China) | Intramuscular | Protein subunit | Phase III clinical trial (NCT04646590) | Yes | Yang et al. [47]. phase 1 and 2 trial 10% (grade 3 or worse) for 50 μg group |
| CoVLP | Medicago Inc. (Quebec, ON, Canada) | Intramuscular | Virus-like particle | Phase III clinical trial (NCT04636697) | No | |
| Ad5-nCoV | Cansino Biologics (Tianjin, China) | Intramuscular | Adenovirus vector (Ad5) | Phase III clinical trial (NCT04526990, NCT04540419) | Yes | Zhu et al. [48] phase 2 trial 9% (grade 3) 1 × 1011 group, 1% (grade 3) 5 × 1010 group |
| Sputnik V | Gamaleya (Moscow, Russia) | Intramuscular | Adenovirus vector (Ad5 + Ad26) | Phase III clinical trial (NCT04640233, NCT04642339, NCT04656613, NCT04741061, NCT04564716, NCT4530396) | Yes | Logunov et al. [49]. phase 3 trial 0.38% (grade 3) |
| Ad26.COV2.S | Johnson and Johnson (Janssen) (Beerse, Belgium) | Intramuscular | Adenovirus vector (Ad26) | Phase III clinical trial (NCT04838795, NCT04505722, NCT04614948) | Yes | Sadoff et al. [50]. phase 3 trial 0.4% (serious adverse events) |
| AZD1222 | AstraZeneca (Cambridge, UK), Oxford university (Oxford, UK) | Intramuscular | ChAdOx1 | Phase III clinical trial (NCT04864561, NCT04800133, NCT04536051, NCT04516746, NCT04400838, NCT04540393) | Yes | Voysey et al. [51]. pooled four trials 0.7% (serious adverse events) |
| INO-4800 | Inovio (Plymouth Meeting, USA), International vaccine institute (Seoul, South Korea) | Intradermal | DNA vaccine | Phase III clinical trial (NCT04642638) | No | Tebas et al. [52]. phase 1 trial No |
| BNT162b2 | Pfizer (New York, USA), BioNTech (Mainz, Germany) | Intramuscular | mRNA vaccine | Phase III clinical trial (NCT04368728, NCT04805125, NCT04800133, NCT04816669, NCT04713553, NCT04754594) | Yes | Polack et al. [53]. phase 2/3 trial 4/43,252 (serious adverse events), 2/43,252 (died) |
| mRNA-1273 | Moderna (Cambridge, USA), NIAID (North Bathesda, USA) | Intramuscular | mRNA vaccine | Phase III clinical trial (NCT04860297, NCT04806113, NCT04649151, NCT04470427, NCT04796896, NCT04811664, NCT04805125) | Yes | Baden et al. [54]. phase 3 trial 1.5% (grade 3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, J.H.; Lee, H.K. Delivery Routes for COVID-19 Vaccines. Vaccines 2021, 9, 524. https://doi.org/10.3390/vaccines9050524
AMA Style
Park JH, Lee HK. Delivery Routes for COVID-19 Vaccines. Vaccines. 2021; 9(5):524. https://doi.org/10.3390/vaccines9050524
Chicago/Turabian StylePark, Jang Hyun, and Heung Kyu Lee. 2021. "Delivery Routes for COVID-19 Vaccines" Vaccines 9, no. 5: 524. https://doi.org/10.3390/vaccines9050524
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.