
Impact of Early Pandemic SARS-CoV-2 Lineages Replacement with the Variant of Concern P.1 (Gamma) in Western Bahia, Brazil

 , , , , , , ,
, , , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. RNA Extraction and RT-qPCR
2.3. Viral Genotyping by RT-qPCR
2.4. SARS-CoV-2 Genome SEQUENCING
2.5. Phylogenetic Analysis
2.6. Statistical Analyses
3. Results
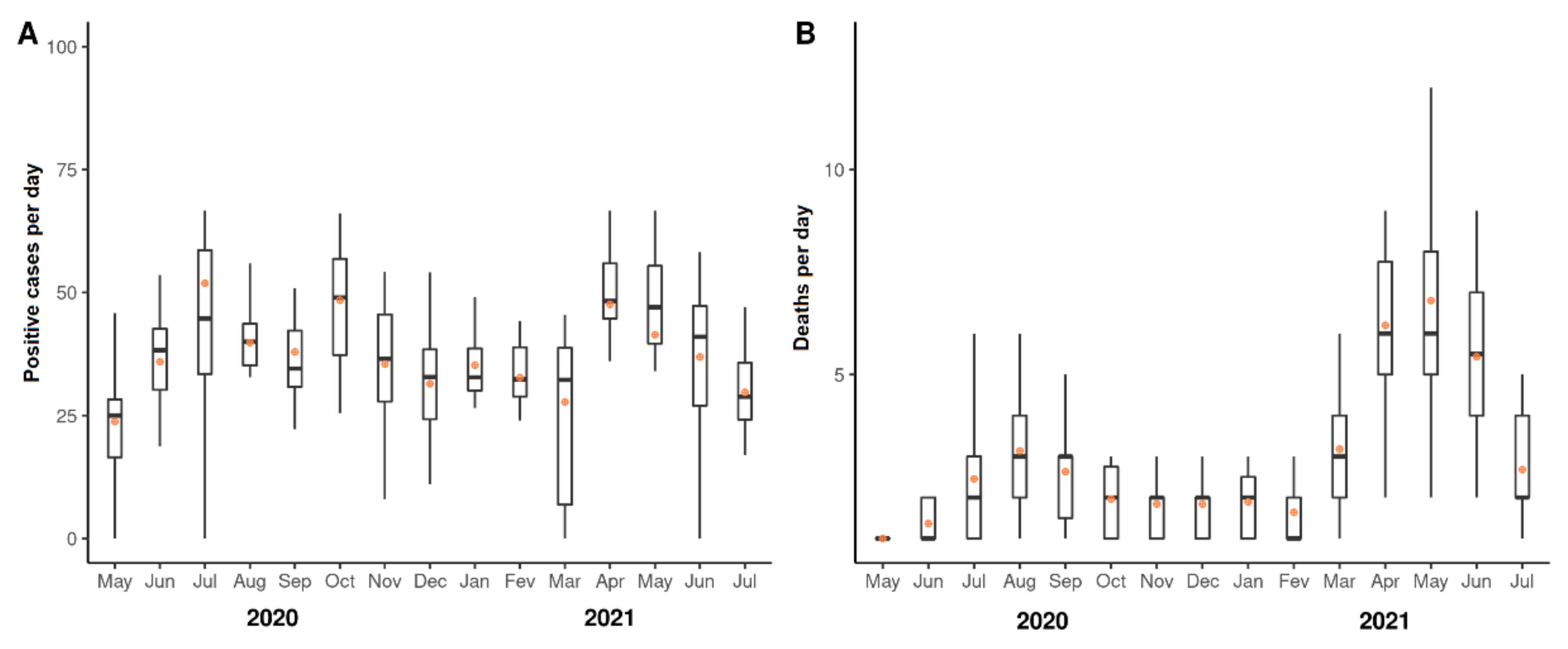
3.1. Fluctuations of Numbers of COVID-19 Cases and Deaths from May 2020 to July 2021
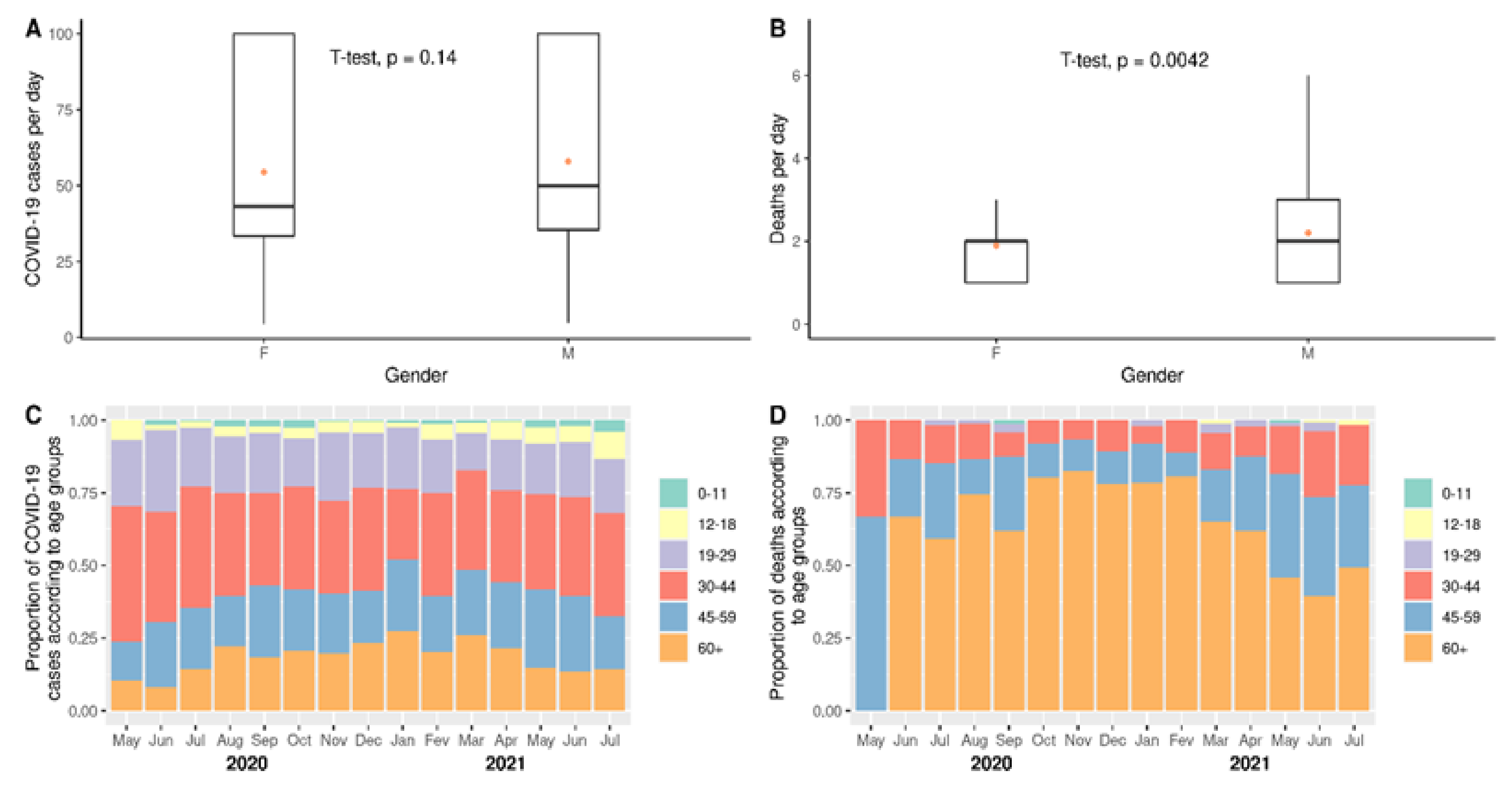
3.2. Impact of COVID-19 According to Gender or Age
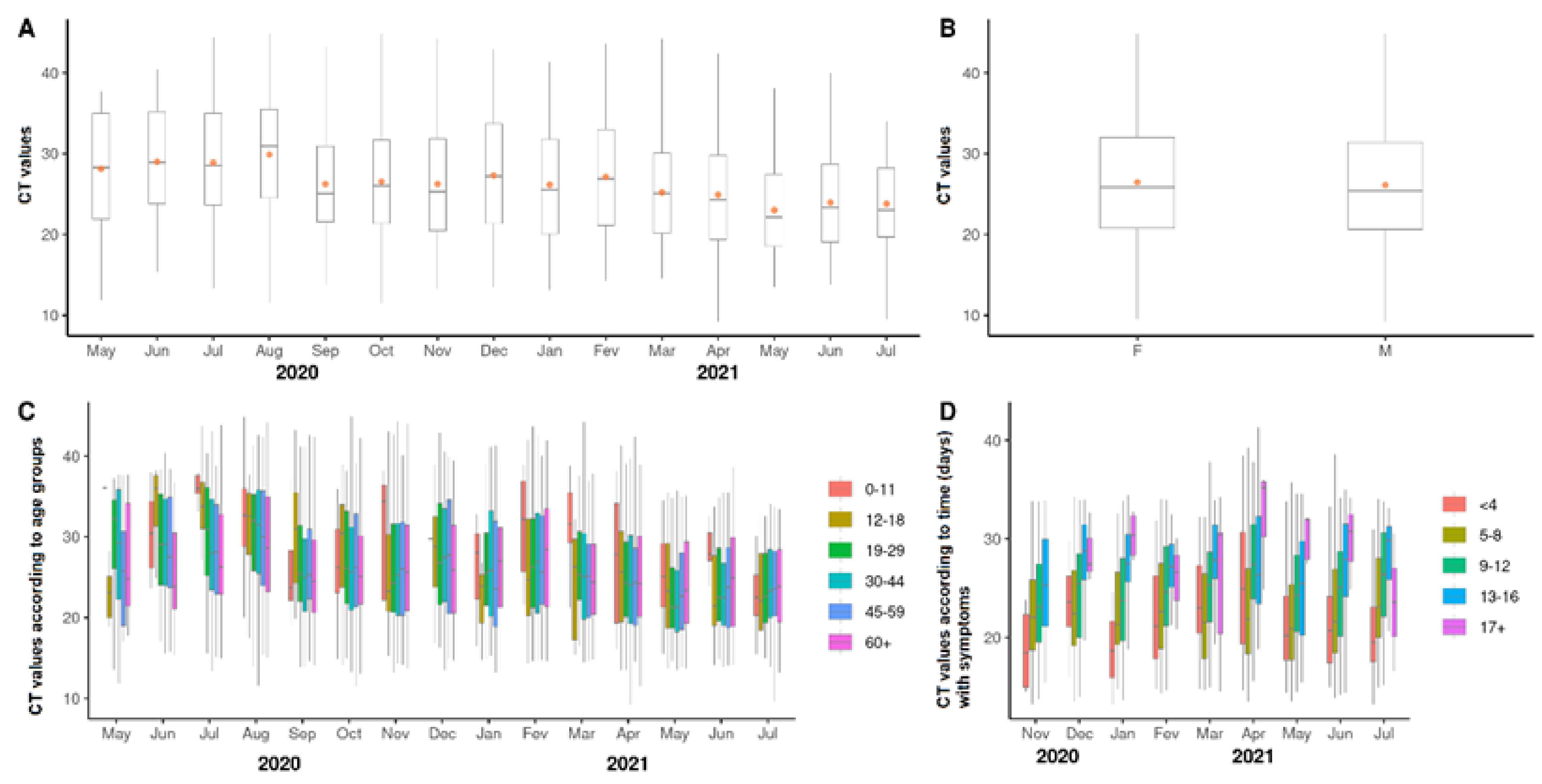
3.3. Viral Loads According to Cycle Threshold Values
3.4. Substitution of SARS-CoV-2 Lineages and Its Impact on Local Health
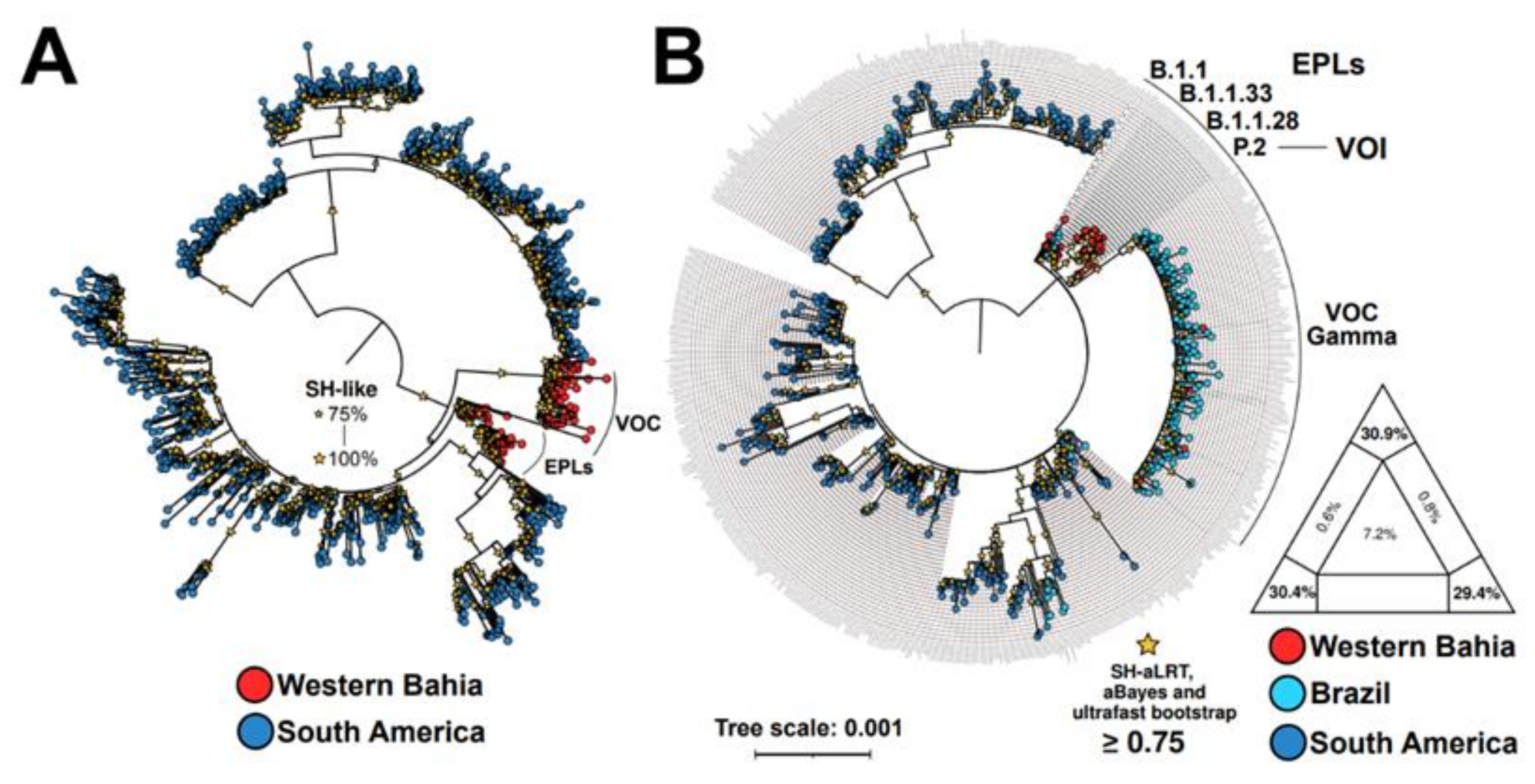
3.5. Viral Phylogeny
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Coronavirus (COVID-19) Dashboard|WHO Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 26 September 2022).
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Farias, J.P.; da Silva, P.S.; Fogaça, M.M.C.; Santana, I.V.; Luiz, W.B.; Birbrair, A.; Amorim, J.H. The COVID-19 humoral immunological status induced by CoronaVac and AstraZeneca vaccines significantly benefits from a booster shot with the Pfizer vaccine. J. Virol. 2022, 96, e0017722. [Google Scholar] [CrossRef]
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Munnink, B.B.O.; van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.S.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Eurosurveillance 2020, 25, 2001005. [Google Scholar] [CrossRef]
- Triggle, C.R.; Bansal, D.; Ding, H.; Islam, M.M.; Farag, E.A.B.A.; Hadi, H.A.; Sultan, A.A. A Comprehensive Review of Viral Char-acteristics, Transmission, Pathophysiology, Immune Response, and Management of SARS-CoV-2 and COVID-19 as a Basis for Controlling the Pandemic. Front. Immunol. 2021, 12, 1–23. [Google Scholar]
- Marcin, F.D.; Martin, S.W.; Tomas, S.; Sara, C.; Shankar, M.; Lachmann, G.; Monneret, G.; Venet, F.; Bauer, M.; Brunkhorst, F.M.; et al. The COVID-19 puzzle: Deciphering pathophysiology and phenotypes of a new disease entity|Elsevier Enhanced Reader. Lancet Respir. Med. 2021, 9, 622–642. [Google Scholar]
- Trougakos, I.P.; Stamatelopoulos, K.; Terpos, E.; Tsitsilonis, O.E.; Aivalioti, E.; Paraskevis, D.; Kastritis, E.; Pavlakis, G.N.; Dimopoulos, M.A. Insights to SARS-CoV-2 life cycle, pathophysiology, and rationalized treatments that target COVID-19 clinical complications. J. Biomed. Sci. 2021, 28, 1–18. [Google Scholar] [CrossRef]
- COVID-19 Vaccine Effectiveness|CDC. Available online: https://www.cdc.gov/coronavirus/2019-ncov/vaccines/effectiveness/index.html (accessed on 26 June 2022).
- Vaccine Efficacy, Effectiveness and Protection. Available online: https://www.who.int/news-room/feature-stories/detail/vaccine-efficacy-effectiveness-and-protection (accessed on 26 June 2022).
- Koelle, K.; Martin, M.A.; Antia, R.; Lopman, B.; Dean, N.E. The changing epidemiology of SARS-CoV-2. Science 2022, 375, 1116–1121. [Google Scholar] [CrossRef]
- Da Silva, S.J.R.; Pena, L. Collapse of the public health system and the emergence of new variants during the second wave of the COVID-19 pandemic in Brazil. One Health 2021, 13, 100287. [Google Scholar] [CrossRef] [PubMed]
- Xavier, D.R.; Lima e Silva, E.; Lara, F.A.; e Silva, G.R.R.; Oliveira, M.F.; Gurgel, H.; Barcelos, C. Involvement of political and so-cio-economic factors in the spatial and temporal dynamics of COVID-19 outcomes in Brazil: A population-based study. Lancet Reg. Health—Am. 2022, 10, 100221. [Google Scholar] [PubMed]
- Brusselaers, N.; Steadson, D.; Bjorklund, K.; Breland, S.; Stilhoff, S.J.; Ewing, A.; Bergmann, S.; Steineck, G. Evaluation of science advice during the COVID-19 pandemic in Sweden. Humanit. Soc. Sci. Commun. 2022, 9, 1–17. [Google Scholar]
- Riley, S.; Ainslie, K.E.C.; Eales, O.; Walters, C.E.; Wang, H.; Atchison, C.; Fronterre, C.; Diggle, P.J.; Ashby, D.; Donnelly, C.A.; et al. Resurgence of SARS-CoV-2: Detection by community viral surveillance. Science 2021, 372, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Wikle, N.B.; Tran, T.N.-A.; Gentilesco, B.; Leighow, S.M.; Albert, E.; Strong, E.R.; Brinda, K.; Inam, H.; Yang, F.; Hossain, S.; et al. SARS-CoV-2 epidemic after social and economic reopening in three U.S. states reveals shifts in age structure and clinical characteristics. Sci. Adv. 2022, 8, eabf9868. [Google Scholar] [CrossRef] [PubMed]
- McCormick, K.D.; Jacobs, J.L.; Mellors, J.W. The emerging plasticity of SARS-CoV-2. Science 2021, 371, 1306–1308. [Google Scholar] [CrossRef]
- Rocha, A.L.S.; Pinheiro, J.R.; Nakamura, T.C.; da Silva, J.D.S.; Rocha, B.G.S.; Klein, R.C.; Birbrair, A.; Amorim, J.H. Fomites and the environment did not have an important role in COVID-19 transmission in a Brazilian mid-sized city. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- A novel RT-qPCR Assay for Detection of SARS-CoV-2 Variants based on RhAmp Technology (IDT Technologies). Available online: https://www.protocols.io/view/a-novel-rt-qpcr-assay-for-detection-of-sars-cov-2-x54v9jwbmg3e/v1 (accessed on 2 May 2022).
- Silva, J.P.; de Lima, A.B.; Alvim, L.B.; Malta, F.S.; Mendonça, C.P.; Fonseca, P.L.C.; Moreira, F.R.R.; Queiroz, D.C.; Ferreira, J.G.G.; Ferreira, A.C.S.; et al. Delta Variant of SARS-CoV-2 Replacement in Brazil: A National Epidemiologic Surveillance Program. Viruses 2022, 14, 847. [Google Scholar] [CrossRef]
- Nascimento, V.A.D.; Corado, A.D.L.G.; Nascimento, F.O.D.; da Costa, Á.K.A.; Duarte, D.C.G.; Luz, S.L.B.; Goncalves, L.M.F.; de Jesus, M.S.; da Costa, C.F.; Delatorre, E.; et al. Genomic and phylogenetic characterisation of an imported case of sars-cov-2 in amazonas state, brazil. Mem. Inst. Oswaldo Cruz 2020, 115, 1–6. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE Team. Unipro UGENE: A unified bioinfor-matics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Gascuel, O. BIONJ: An improved version of the NJ algorithm based on a simple model of sequence data. Mol. Biol. Evol. 1997, 14, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, H. An Approximately Unbiased Test of Phylogenetic Tree Selection. Syst. Biol. 2002, 51, 492–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Coronavirus COVID-19 Dashboard. Available online: https://covid19.who.int (accessed on 8 December 2021).
- Boletins Epidemiológicos—COVID-19 Sesab. Available online: http://www.saude.ba.gov.br/temasdesaude/coronavirus/boletins-epidemiologicos-covid-19/ (accessed on 14 September 2022).
- Página Inicial—Português (Brasil). Available online: https://www.gov.br/turismo/pt-br (accessed on 14 September 2022).
- Planos Estaduais e Comunicados (Covid-19)|Sesab. Available online: http://www.saude.ba.gov.br/temasdesaude/coronavirus/planos-estaduais-e-comunicados-covid-19/ (accessed on 14 September 2022).
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Apalkova, Y.; Butovskaya, M.L.; Bronnikova, N.; Burkova, V.; Shackelford, T.K.; Fink, B. Assessment of Male Physical Risk-Taking Behavior in a Sample of Russian Men and Women. Evol. Psychol. Sci. 2018, 4, 314–321. [Google Scholar] [CrossRef]
- He, X.; Lau, E.H.Y.; Wu, P.; Deng, X.; Wang, J.; Hao, X.; Lau, Y.C.; Wong, J.Y.; Guan, Y.; Tan, X.; et al. Temporal dynamics in viral shedding and transmissibility of COVID-19. Nat. Med. 2020, 26, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Argyropoulos, K.V.; Serrano, A.; Hu, J.; Black, M.; Feng, X.; Shen, G.; Call, M.; Kim, M.J.; Lytle, A.; Belovarac, B.; et al. Association of Initial Viral Load in Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Patients with Outcome and Symptoms. Am. J. Pathol. 2020, 190, 1881–1887. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinheiro, J.R.; dos Reis, E.C.; Farias, J.P.; Fogaça, M.M.C.; da Silva, P.d.S.; Santana, I.V.R.; Rocha, A.L.S.; Vidal, P.O.; Simões, R.d.C.; Luiz, W.B.; et al. Impact of Early Pandemic SARS-CoV-2 Lineages Replacement with the Variant of Concern P.1 (Gamma) in Western Bahia, Brazil. Viruses 2022, 14, 2314. https://doi.org/10.3390/v14102314
Pinheiro JR, dos Reis EC, Farias JP, Fogaça MMC, da Silva PdS, Santana IVR, Rocha ALS, Vidal PO, Simões RdC, Luiz WB, et al. Impact of Early Pandemic SARS-CoV-2 Lineages Replacement with the Variant of Concern P.1 (Gamma) in Western Bahia, Brazil. Viruses. 2022; 14(10):2314. https://doi.org/10.3390/v14102314
Chicago/Turabian StylePinheiro, Josilene R., Esther C. dos Reis, Jéssica P. Farias, Mayanna M. C. Fogaça, Patrícia de S. da Silva, Itana Vivian R. Santana, Ana Luiza S. Rocha, Paloma O. Vidal, Rafael da C. Simões, Wilson B. Luiz, and et al. 2022. "Impact of Early Pandemic SARS-CoV-2 Lineages Replacement with the Variant of Concern P.1 (Gamma) in Western Bahia, Brazil" Viruses 14, no. 10: 2314. https://doi.org/10.3390/v14102314