

A Rapid and Simple Multiparameter Assay to Quantify Spike-Specific CD4 and CD8 T Cells after SARS-CoV-2 Vaccination: A Preliminary Report

 ,
,  ,
,  and
and
Abstract
:1. Introduction
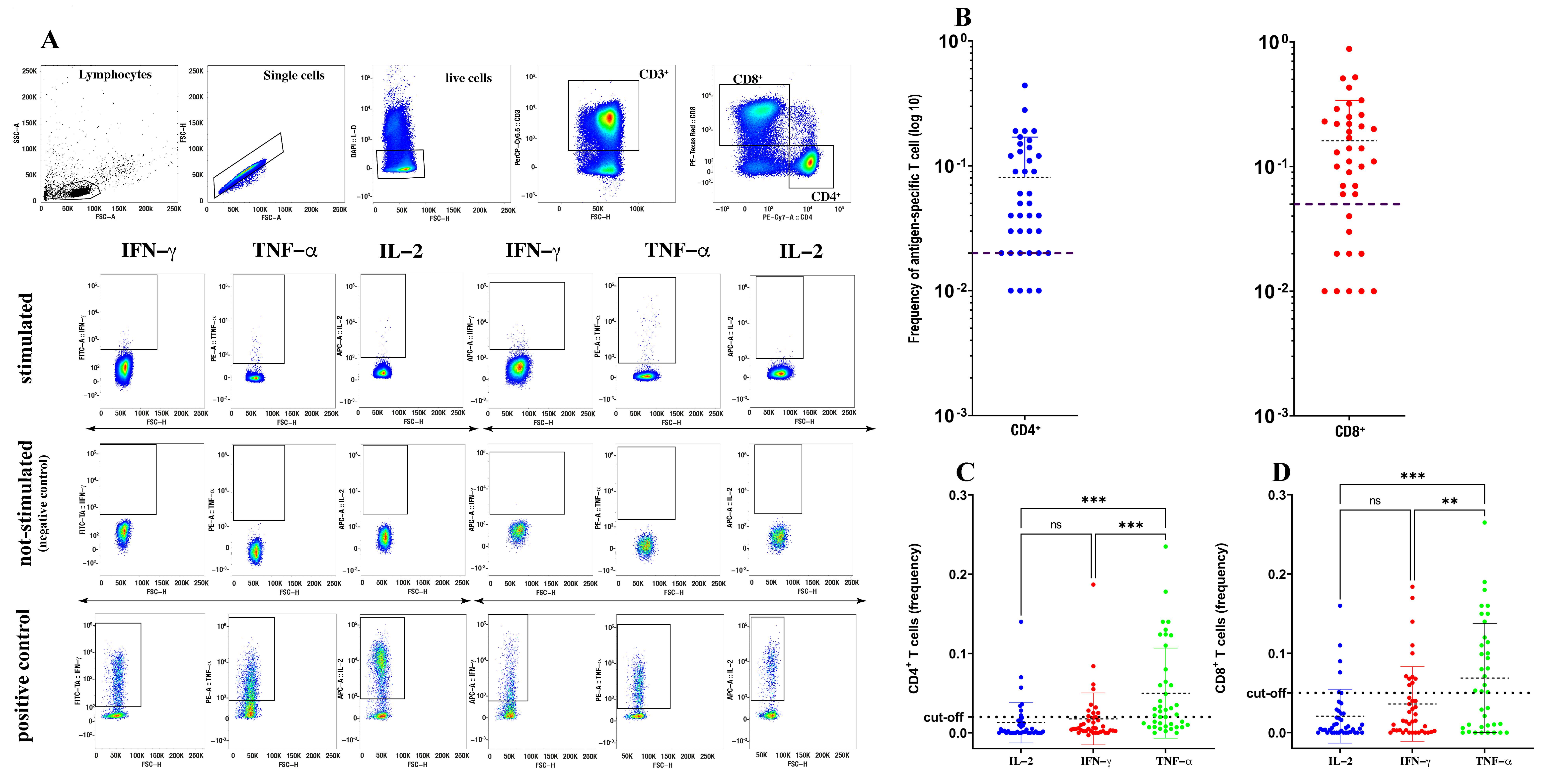
2. Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cucinotta, D.; Vanelli, M. WHO declares COVID-19 a pandemic. Acta Bio Med. Atenei Parm. 2020, 91, 157. [Google Scholar]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.T.; Girardin, R.C.; Dupuis, A.P.; Kulas, K.E.; Payne, A.F.; Wong, S.J.; Arinsburg, S.; Nguyen, F.T.; Mendu, D.R.; Firpo-Betancourt, A. Neutralizing antibody responses in COVID-19 convalescent sera. J. Infect. Dis. 2021, 223, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Fenrich, M.; Mrdenovic, S.; Balog, M.; Tomic, S.; Zjalic, M.; Roncevic, A.; Mandic, D.; Debeljak, Z.; Heffer, M. SARS-CoV-2 dissemination through peripheral nerves explains multiple organ injury. Front. Cell. Neurosci. 2020, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef] [PubMed]
- Sattler, A.; Angermair, S.; Stockmann, H.; Heim, K.M.; Khadzhynov, D.; Treskatsch, S.; Halleck, F.; Kreis, M.E.; Kotsch, K. SARS-CoV-2-specific T cell responses and correlations with COVID-19 patient predisposition. J. Clin. Investig. 2020, 130, 6477–6489. [Google Scholar] [CrossRef] [PubMed]
- Riou, C.; Schäfer, G.; du Bruyn, E.; Goliath, R.T.; Stek, C.; Mou, H.; Hung, D.; Wilkinson, K.A.; Wilkinson, R.J. Rapid, simplified whole blood-based multiparameter assay to quantify and phenotype SARS-CoV-2 specific T cells. medRxiv 2020. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.-D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.G.; Smits, K.; Joosten, S.A.; van Meijgaarden, K.E.; Satti, I.; Fletcher, H.A.; Caccamo, N.; Dieli, F.; Mascart, F.; McShane, H.; et al. Intracellular Cytokine Staining and Flow Cytometry: Considerations for Application in Clinical Trials of Novel Tuberculosis Vaccines. PLoS ONE 2015, 10, e0138042. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, S.; Krawczyk, A.; Knop, D.; Klump, H.; Heinold, A.; Heinemann, F.M.; Thümmler, L.; Temme, C.; Breyer, M.; Witzke, O.; et al. Cellular Immunity in COVID-19 Convalescents with PCR-Confirmed Infection but with Undetectable SARS-CoV-2-Specific IgG. Emerg.Infect. Dis. 2021, 27. [Google Scholar] [CrossRef]
- Kalimuddin, S.; Tham, C.Y.L.; Qui, M.; de Alwis, R.; Sim, J.X.Y.; Lim, J.M.E.; Tan, H.-C.; Syenina, A.; Zhang, S.L.; Le Bert, N.; et al. Early T cell and binding antibody responses are associated with COVID-19 RNA vaccine efficacy onset. Med 2021, 2, 682–688.e4. [Google Scholar] [CrossRef] [PubMed]
- Bange, E.M.; Han, N.A.; Wileyto, P.; Kim, J.Y.; Gouma, S.; Robinson, J.; Greenplate, A.R.; Hwee, M.A.; Porterfield, F.; Owoyemi, O.; et al. CD8+ T cells contribute to survival in patients with COVID-19 and hematologic cancer. Nat. Med. 2021, 27, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Herishanu, Y.; Avivi, I.; Aharon, A.; Shefer, G.; Levi, S.; Bronstein, Y.; Morales, M.; Ziv, T.; Shorer Arbel, Y.; Scarfò, L.; et al. Efficacy of the BNT162b2 mRNA COVID-19 vaccine in patients with chronic lymphocytic leukemia. Blood 2021, 137, 3165–3173. [Google Scholar] [CrossRef] [PubMed]
- Connolly, C.M.; Boyarsky, B.J.; Ruddy, J.A.; Werbel, W.A.; Christopher-Stine, L.; Garonzik-Wang, J.M.; Segev, D.L.; Paik, J.J. Absence of Humoral Response After Two-Dose SARS-CoV-2 Messenger RNA Vaccination in Patients with Rheumatic and Musculoskeletal Diseases: A Case Series. Ann. Intern. Med. 2021, 174, 1332–1334. [Google Scholar] [CrossRef] [PubMed]
- Spiera, R.; Jinich, S.; Jannat-Khah, D. Rituximab, but not other antirheumatic therapies, is associated with impaired serological response to SARS- CoV-2 vaccination in patients with rheumatic diseases. Ann. Rheum. Dis. 2021, 80, 1357. [Google Scholar] [CrossRef] [PubMed]
- Hagin, D.; Freund, T.; Navon, M.; Halperin, T.; Adir, D.; Marom, R.; Levi, I.; Benor, S.; Alcalay, Y.; Freund, N.T. Immunogenicity of Pfizer-BioNTech COVID-19 vaccine in patients with inborn errors of immunity. J. Allergy Clin. Immunol. 2021, 148, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Malipiero, G.; Moratto, A.; Infantino, M.; D’Agaro, P.; Piscianz, E.; Manfredi, M.; Grossi, V.; Benvenuti, E.; Bulgaresi, M.; Benucci, M.; et al. Assessment of humoral and cellular immunity induced by the BNT162b2 SARS-CoV-2 vaccine in healthcare workers, elderly people, and immunosuppressed patients with autoimmune disease. Immunol. Res. 2021, 1–8. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| G1 HD, Nab < 390 BAU/mL | G2 HD, 390 < Nab < 1040 BAU/mL | G3 HD, Nab > 1040 BAU/mL | G4 Patients Ocrelizumab or Primitive B Cell Deficiences | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Enrolled Subjects (%) | 9 | (22.5%) | 8 | (20%) | 11 | (27.5%) | 12 | (30%) | 40 | (100%) |
| Mean Age | 50 | 44 | 45 | 41 | 44 | |||||
| Range | 27–66 | 27–59 | 22–55 | 21–64 | 22–66 | |||||
| Male Gender (%) | 5 | (55.7%) | 3 | (37.5%) | 3 | (77.5%) | 3 | (27.3%) | 14 | (35%) |
| G4 Enrolled Patients | Pathology | CCI | Therapy | N Days | IgG mg/dL |
|---|---|---|---|---|---|
| ID20 | RRMS | 0 | Ocrelizumab | 180 | - |
| ID30 | RRMS | 0 | Ocrelizumab | 180 | - |
| ID31 | RRMS | 0 | Ocrelizumab | 180 | - |
| ID39 | RRMS | 1 | Ocrelizumab | 160 | - |
| ID40 | SPMS | 2 | Ocrelizumab | 180 | - |
| ID41 | RRMS | 0 | Ocrelizumab | 180 | - |
| ID25 | CVID | - | IgG Vein Kendrion | 28 | >600 |
| ID38 | CVID | - | Subcutaneous IgG Hyqvia Takeda | 28 | >600 |
| ID45 | XLA | - | Subcutaneous IgG Hyqvia Takeda | 28 | >600 |
| ID46 | CIVD | - | CSL Bhering Hizentra | 15 | >600 |
| ID47 | XLA | - | IgG Vein Kendrion | 28 | >600 |
| ID48 | PNH | - | CSL Bhering Hizentra | 15 | >600 |
| Enrolled Patients | NAb BAU/mL | Frequency (%) of CD4+ T Cells | Frequency (%) of CD8+ T Cells |
| ID20 | 0 | 0.0558 | 0.0174 |
| ID25 | 0 | 0.1933 | 0.2069 |
| ID30 | 0 | 0.0271 | 0.0136 |
| ID31 | 0 | 0.0600 | 0.5232 |
| ID38 | 0 | 0.0350 | 0.0256 |
| ID39 | 0 | 0.0021 | 0.1285 |
| ID40 | 0 | 0.0390 | 0.0230 |
| ID41 | 0 | 0.0867 | 0.0631 |
| ID45 | 0 | 0.0359 | 0.0969 |
| ID46 | 0 | 0.0120 | 0.5109 |
| ID47 | 0 | 0.1700 | 0.1440 |
| ID48 | 0 | 0.0120 | 0.2530 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shekarkar Azgomi, M.; La Manna, M.P.; Badami, G.D.; Ragonese, P.; Trizzino, A.; Dieli, F.; Caccamo, N. A Rapid and Simple Multiparameter Assay to Quantify Spike-Specific CD4 and CD8 T Cells after SARS-CoV-2 Vaccination: A Preliminary Report. Biomedicines 2021, 9, 1576. https://doi.org/10.3390/biomedicines9111576
Shekarkar Azgomi M, La Manna MP, Badami GD, Ragonese P, Trizzino A, Dieli F, Caccamo N. A Rapid and Simple Multiparameter Assay to Quantify Spike-Specific CD4 and CD8 T Cells after SARS-CoV-2 Vaccination: A Preliminary Report. Biomedicines. 2021; 9(11):1576. https://doi.org/10.3390/biomedicines9111576
Chicago/Turabian StyleShekarkar Azgomi, Mojtaba, Marco Pio La Manna, Giusto Davide Badami, Paolo Ragonese, Antonino Trizzino, Francesco Dieli, and Nadia Caccamo. 2021. "A Rapid and Simple Multiparameter Assay to Quantify Spike-Specific CD4 and CD8 T Cells after SARS-CoV-2 Vaccination: A Preliminary Report" Biomedicines 9, no. 11: 1576. https://doi.org/10.3390/biomedicines9111576