
MicroRNA-Mediated Regulation of the Virus Cycle and Pathogenesis in the SARS-CoV-2 Disease

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification of SARS-CoV-2-Derived miRNA Precursors
2.2. Identification of SARS-CoV-2 Mature miRNAs
2.3. Prediction of Biological Pathways Regulated by SARS-CoV-2 miRNAs
2.4. Identification of Human miRNAs Targeting SARS-CoV-2-Specific Genome Sequences
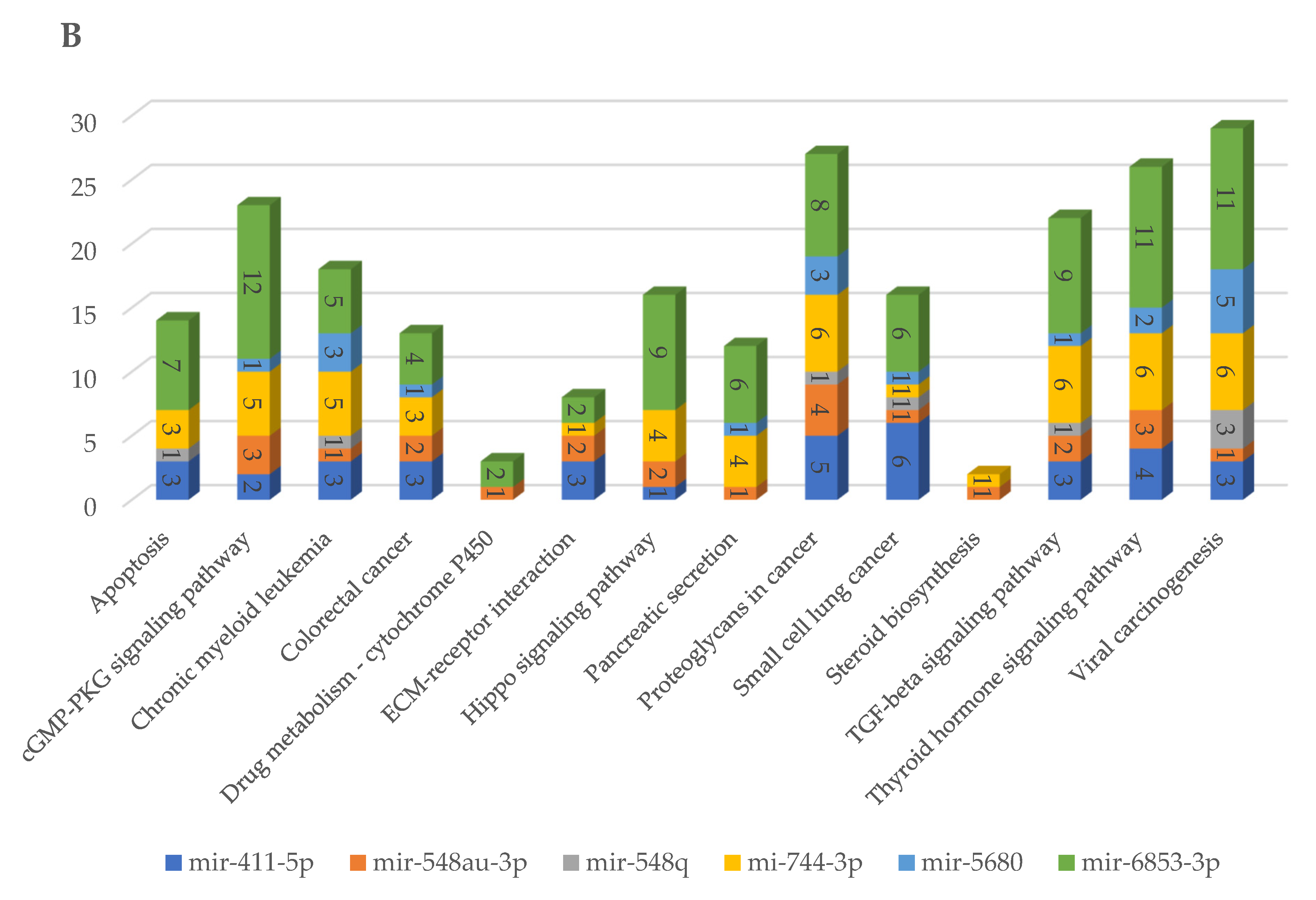
2.5. Prediction of Biological Pathways Regulated by miRNAs Able to Target Viral Genome
2.6. Analysis of SARS-CoV-2 Variants of Concern (VOCs)
3. Discussion
4. Materials and Methods
4.1. Prediction of SARS-CoV-2-Derived miRNA Precursors (pre-miRNAs) Specific Genome Sequences
4.2. Methods for Selection of V-miRNAs
4.3. Prediction of Molecular Pathway Regulated by V-miRNAs
4.4. Prediction of Human miRNAs Targeting the SARS-CoV-2 Genome
4.5. Prediction of Tissue-Specific Expression of miRNAs and Their Involvement in Human Disease
4.6. Prediction of Molecular Pathways Regulated by hsa-miRNAs
4.7. Analysis of SARS-CoV-2 Variants of Concern (VOCs)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gordon, D.E.; Hiatt, J.; Bouhaddou, M.; Rezelj, V.V.; Ulferts, S.; Braberg, H.; Jureka, A.S.; Obernier, K.; Guo, J.Z.; Batra, J.; et al. Comparative host-coronavirus protein interaction networks reveal pan-viral disease mechanisms. Science 2020, 370, eabe9403. [Google Scholar] [CrossRef]
- Sharma, A.; Tiwari, S.; Deb, M.K.; Marty, J.L. Severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2): A global pandemic and treatment strategies. Int. J. Antimicrob. Agents 2020, 56, 106054. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, J.-Y.; Yang, J.-S.; Kim, J.W.; Kim, V.N.; Chang, H. The Architecture of SARS-CoV-2 Transcriptome. Cell 2020, 181, 914–921.e10. [Google Scholar] [CrossRef]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Fu, L.; Wang, B.; Yuan, T.; Chen, X.; Ao, Y.; Fitzpatrick, T.; Li, P.; Zhou, Y.; Lin, Y.-F.; Duan, Q.; et al. Clinical characteristics of coronavirus disease 2019 (COVID-19) in China: A systematic review and meta-analysis. J. Infect. 2020, 80, 656–665. [Google Scholar] [CrossRef]
- Liu, J.; Xie, W.; Wang, Y.; Xiong, Y.; Chen, S.; Han, J.; Wu, Q. A comparative overview of COVID-19, MERS and SARS: Review article. Int. J. Surg. 2020, 81, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Qiu, J.; Aryal, S.; Hackett, J.L.; Wang, J. The RNA Architecture of the SARS-CoV-2 3′-Untranslated Region. Viruses 2020, 12, 1473. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Delorey, T.M.; Ziegler, C.G.K.; Heimberg, G.; Normand, R.; Yang, Y.; Segerstolpe, Å.; Abbondanza, D.; Fleming, S.J.; Subramanian, A.; Montoro, D.T.; et al. COVID-19 tissue atlases reveal SARS-CoV-2 pathology and cellular targets. Nature 2021, 595, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Kream, R.M.; Stefano, G.B. Long-Term Respiratory and Neurological Sequelae of COVID-19. Med. Sci. Monit. 2020, 26, e928996. [Google Scholar] [CrossRef]
- Boldrini, M.; Canoll, P.D.; Klein, R.S. How COVID-19 Affects the Brain. JAMA Psychiatry 2021, 78, 682–683. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.; Shao, C.; Huang, J.; Gan, J.; Huang, X.; Bucci, E.; Piacentini, M.; Ippolito, G.; Melino, G. COVID-19 infection: The perspectives on immune responses. Cell Death Differ. 2020, 27, 1451–1454. [Google Scholar] [CrossRef] [Green Version]
- Alkhatib, M.; Svicher, V.; Salpini, R.; Ambrosio, F.A.; Bellocchi, M.C.; Carioti, L.; Piermatteo, L.; Scutari, R.; Costa, G.; Artese, A.; et al. SARS-CoV-2 variants and their relevant mutational profiles: Update summer 2021. Microbiol. Spectr. 2021, 9, e0109621. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; COVID-19 Genomics UK (COG-UK) Consortium; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Ballesteros, N.; Muñoz, M.; Patiño, L.H.; Hernández, C.; González-Casabianca, F.; Carroll, I.; Santos-Vega, M.; Cascante, J.; Angel, A.; Feged-Rivadeneira, A.; et al. Deciphering the introduction and transmission of SARS-CoV-2 in the Colombian Amazon Basin. PLoS Negl. Trop. Dis. 2021, 15, e0009327. [Google Scholar] [CrossRef] [PubMed]
- Falcone, M.; Tiseo, G.; Valoriani, B.; Barbieri, C.; Occhineri, S.; Mazzetti, P.; Vatteroni, M.L.; Suardi, L.R.; Riccardi, N.; Pistello, M.; et al. Efficacy of bamlanivimab/etesevimab and casirivimab/imdevimab in preventing progression to severe COVID-19 and role of variants of concern. Infect. Dis. Ther. 2021, 10, 2479–2488. [Google Scholar] [CrossRef]
- Hoffmann, M.; Arora, P.; Groß, R.; Seidel, A.; Hörnich, B.F.; Hahn, A.S.; Krüger, N.; Graichen, L.; Hofmann-Winkler, H.; Kempf, A.; et al. SARS-CoV-2 variants B.1.351 and P.1 escape from neutralizing antibodies. Cell 2021, 184, 2384–2393. [Google Scholar] [CrossRef]
- Gao, S.-J.; Guo, H.; Luo, G. Omicron variant (B.1.1.529) of SARS-CoV-2, a global urgent public health alert! J. Med. Virol. 2021. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Barbu, M.G.; Condrat, C.E.; Thompson, D.C.; Bugnar, O.L.; Cretoiu, D.; Toader, O.D.; Suciu, N.; Voinea, S.C. MicroRNA Involvement in Signaling Pathways During Viral Infection. Front. Cell Dev. Biol. 2020, 8, 143. [Google Scholar] [CrossRef] [Green Version]
- Zhan, S.; Wang, Y.; Chen, X. RNA virus-encoded microRNAs: Biogenesis, functions and perspectives on application. ExRNA 2020, 2, 15. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, Z.; Mallick, B.; Chakrabarti, J. Cellular versus viral microRNAs in host-virus interaction. Nucleic Acids Res. 2009, 37, 1035–1048. [Google Scholar] [CrossRef]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of hepatitis C virus RNA abundance by a liver-specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathans, R.; Chu, C.-Y.; Serquina, A.K.; Lu, C.-C.; Cao, H.; Rana, T.M. Cellular microRNA and P bodies modulate host-HIV-1 interactions. Mol. Cell 2009, 34, 696–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuovo, G.J.; Wu, X.; Volinia, S.; Yan, F.; di Leva, G.; Chin, N.; Nicol, A.F.; Jiang, J.; Otterson, G.; Schmittgen, T.D.; et al. Strong inverse correlation between microRNA-125b and human papillomavirus DNA in productive infection. Diagn. Mol. Pathol. 2010, 19, 135–143. [Google Scholar] [CrossRef]
- Sartorius, K.; Makarova, J.; Sartorius, B.; An, P.; Winkler, C.; Chuturgoon, A.; Kramvis, A. The regulatory role of MicroRNA in hepatitis-B virus-associated hepatocellular carcinoma (HBV-HCC) pathogenesis. Cells 2019, 8, 1504. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, P.; Biswas, S.C. Small non-coding RNAs: Do they encode answers for controlling SARS-CoV-2 in the future? Front. Microbiol. 2020, 11, 571553. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, F.; Argyris, E.; Chen, K.; Liang, Z.; Tian, H.; Huang, W.; Squires, K.; Verlinghieri, G.; Zhang, H. Cellular microRNAs contribute to HIV-1 latency in resting primary CD4+ T lymphocytes. Nat. Med. 2007, 13, 1241–1247. [Google Scholar] [CrossRef]
- Hussain, M.; Torres, S.; Schnettler, E.; Funk, A.; Grundhoff, A.; Pijlman, G.P.; Khromykh, A.A.; Asgari, S. West Nile virus encodes a microRNA-like small RNA in the 3′ untranslated region which up-regulates GATA4 mRNA and facilitates virus replication in mosquito cells. Nucleic Acids Res. 2012, 40, 2210–2223. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Asgari, S. MicroRNA-like viral small RNA from Dengue virus 2 autoregulates its replication in mosquito cells. Proc. Natl. Acad. Sci. USA 2014, 111, 2746–2751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liang, H.; Chen, X.; Ke, Y.; Zhou, Z.; Yang, M.; Zen, K.; Yang, R.; Liu, C.; Zhang, C.-Y. An Ebola virus-encoded microRNA-like fragment serves as a biomarker for early diagnosis of Ebola virus disease. Cell Res. 2016, 26, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, J.; Zhang, H.; Wang, M.; Gao, G.F.; Li, X. Ebola virus encodes a miR-155 analog to regulate importin-α5 expression. Cell. Mol. Life Sci. 2016, 73, 3733–3744. [Google Scholar] [CrossRef]
- Malik, S.S.; Azem-E-Zahra, S.; Kim, K.M.; Caetano-Anollés, G.; Nasir, A. Do Viruses Exchange Genes across Superkingdoms of Life? Front. Microbiol. 2017, 8, 2110. [Google Scholar] [CrossRef]
- Nasir, A.; Kim, K.M.; Caetano-Anolles, G. Giant viruses coexisted with the cellular ancestors and represent a distinct supergroup along with superkingdoms Archaea, Bacteria and Eukarya. BMC Evol. Biol. 2012, 12, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mughal, F.; Nasir, A.; Caetano-Anollés, G. The origin and evolution of viruses inferred from fold family structure. Arch. Virol. 2020, 165, 2177–2191. [Google Scholar] [CrossRef]
- Aydemir, M.N.; Aydemir, H.B.; Korkmaz, E.M.; Budak, M.; Cekin, N.; Pinarbasi, E. Computationally predicted SARS-COV-2 encoded microRNAs target NFKB, JAK/STAT and TGFB signaling pathways. Gene Rep. 2021, 22, 101012. [Google Scholar] [CrossRef]
- Abbas, S.H.; Pervez, M.T.; Ramzan, A.; Khan, D.A. In-silico analysis of Human miRNAs in SARS-CoV-2 Genome. BioSci. Rev. 2021, 3, 27–41. [Google Scholar] [CrossRef]
- Castrillón-Betancur, J.C.; Urcuqui-Inchima, S. Overexpression of miR-484 and miR-744 in Vero cells alters Dengue virus replication. Mem. Inst. Oswaldo Cruz 2017, 112, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Zhu, N.; Guo, W.; Wang, X.; Li, K.; Yan, J.; Jiang, C.; Han, S.; Xiang, H.; Wu, X.; et al. RNA-seq revealed a circular RNA-microRNA-mRNA regulatory network in Hantaan virus infection. Front. Cell. Infect. Microbiol. 2020, 10, 97. [Google Scholar] [CrossRef]
- Xia, L.-H.; Yan, Q.-H.; Sun, Q.-D.; Gao, Y.-P. MiR-411-5p acts as a tumor suppressor in non-small cell lung cancer through targeting PUM1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5546–5553. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, H.; Deng, M.; He, L.; Ping, F.; He, Y.; Fan, Z.; Cheng, B.; Xia, J. Upregulated miR-411-5p levels promote lymph node metastasis by targeting RYBP in head and neck squamous cell carcinoma. Int. J. Mol. Med. 2021, 47, 1. [Google Scholar] [CrossRef] [PubMed]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA Regulation of RNA Virus Replication and Pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef]
- Lagos, D.; Pollara, G.; Henderson, S.; Gratrix, F.; Fabani, M.; Milne, R.S.B.; Gotch, F.; Boshoff, C. miR-132 regulates antiviral innate immunity through suppression of the p300 transcriptional co-activator. Nat. Cell Biol. 2010, 12, 513–519. [Google Scholar] [CrossRef]
- Nersisyan, S.; Engibaryan, N.; Gorbonos, A.; Kirdey, K.; Makhonin, A.; Tonevitsky, A. Potential role of cellular miRNAs in coronavirus-host interplay. PeerJ 2020, 8, e9994. [Google Scholar] [CrossRef] [PubMed]
- Yuen, C.-K.; Lam, J.-Y.; Wong, W.-M.; Mak, L.-F.; Wang, X.; Chu, H.; Cai, J.-P.; Jin, D.-Y.; To, K.K.-W.; Chan, J.F.-W.; et al. SARS-CoV-2 nsp13, nsp14, nsp15 and orf6 function as potent interferon antagonists. Emerg. Microbes Infect. 2020, 9, 1418–1428. [Google Scholar] [CrossRef]
- Sun, X.; Xu, M.; Liu, H.; Ming, K. MicroRNA-219 is downregulated in non-small cell lung cancer and inhibits cell growth and metastasis by targeting HMGA2. Mol. Med. Rep. 2017, 16, 3557–3564. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xu, J.; Hou, Y.; Leverenz, J.B.; Kallianpur, A.; Mehra, R.; Liu, Y.; Yu, H.; Pieper, A.A.; Jehi, L.; et al. Network medicine links SARS-CoV-2/COVID-19 infection to brain microvascular injury and neuroinflammation in dementia-like cognitive impairment. Alzheimers Res. Ther. 2021, 13, 110. [Google Scholar] [CrossRef] [PubMed]
- Barreda-Manso, M.A.; Nieto-Díaz, M.; Soto, A.; Muñoz-Galdeano, T.; Reigada, D.; Maza, R.M. In silico and in vitro analyses validate human MicroRNAs targeting the SARS-CoV-2 3′-UTR. Int. J. Mol. Sci. 2021, 22, 6094. [Google Scholar] [CrossRef] [PubMed]
- Schult, P.; Roth, H.; Adams, R.L.; Mas, C.; Imbert, L.; Orlik, C.; Ruggieri, A.; Pyle, A.M.; Lohmann, V. microRNA-122 amplifies hepatitis C virus translation by shaping the structure of the internal ribosomal entry site. Nat. Commun. 2018, 9, 2613. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, J.; Huang, M.; Xu, G.; Wei, W.; Qin, H. Inhibition of miR-148a-3p resists hepatocellular carcinoma progress of hepatitis C virus infection through suppressing c-Jun and MAPK pathway. J. Cell. Mol. Med. 2019, 23, 1415–1426. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, W.; Ye, W.; Jin, L.; He, J.; Lou, L. microRNAs: Novel players in hepatitis C virus infection. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Triboulet, R.; Mari, B.; Lin, Y.-L.; Chable-Bessia, C.; Bennasser, Y.; Lebrigand, K.; Cardinaud, B.; Maurin, T.; Barbry, P.; Baillat, V.; et al. Suppression of microRNA-silencing pathway by HIV-1 during virus replication. Science 2007, 315, 1579–1582. [Google Scholar] [CrossRef]
- Sironi, M.; Cagliani, R.; Forni, D.; Clerici, M. Evolutionary insights into host-pathogen interactions from mammalian sequence data. Nat. Rev. Genet. 2015, 16, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.B.; Simion, V.; Icli, B.; Pérez-Cremades, D.; Cheng, H.S.; Feinberg, M.W. Computational analysis of targeting SARS-CoV-2, viral entry proteins ACE2 and TMPRSS2, and interferon genes by host MicroRNAs. Genes 2020, 11, 1354. [Google Scholar] [CrossRef]
- Gottwein, E.; Cullen, B.R. Viral and cellular microRNAs as determinants of viral pathogenesis and immunity. Cell Host Microbe 2008, 3, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.-M.; Wang, W.; Song, Z.-G.; Hu, Y.; Tao, Z.-W.; Tian, J.-H.; Pei, Y.-Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Tempel, S.; Tahi, F. A fast ab-initio method for predicting miRNA precursors in genomes. Nucleic Acids Res. 2012, 40, e80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkirtzou, K.; Tsamardinos, I.; Tsakalides, P.; Poirazi, P. MatureBayes: A probabilistic algorithm for identifying the mature miRNA within novel precursors. PLoS ONE 2010, 5, e11843. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SSEARCH | BLASTN | ||||
|---|---|---|---|---|---|
| Viral-Hairpins | Hsa-miRNAs | SR Mismatch | Viral-Harpins | Hsa-miRNAs | SR Mismatch |
| MD306 | miR-2114-5p | 0 | MD134 | miR-190b-5p | 1 |
| MD142 | miR-5680 | 0 | MR186 | miR-744-3p | 1 |
| MR304 | miR-411-5p | 1 | MD311 | miR-4699-3p | 1 |
| MR47 | miR-548au-3p | 1 | MD19 | miR-6730-5p | 1 |
| miR-548q | 1 | MD309 | miR-6838-5p | 1 | |
| MR231 | miR-5683 | 1 | MD309 | miR-181b-3p | 2 |
| MD197 | miR-6853-3p | 1 | MR252 | miR-545-3p | 2 |
| MD297 | miR-6867-5p | 1 | MR186 | miR-4420 | 2 |
| MR231 | miR-5683 | 1 | MD240 | miR-5011-3p | 2 |
| MD324 | miR-411-5p | 2 | |||
| MD195 MD324 | miR-548b-5p | 2 | |||
| MD51 | miR-1267 | 2 | |||
| SSEARCH | BLASTN | ||||
|---|---|---|---|---|---|
| Viral-Hairpins | Hsa-miRNAs | SR Mismatch | Viral-Harpins | Hsa-miRNAs | SR Mismatch |
| MR155 | miR-519c-3p | 0 | MR186 | miR-744-3p | 1 |
| MD142 | miR-5680 | 0 | MR165 | miR-153-5p | 2 |
| MR150 | miR-6074 | 0 | MD324 | miR-411-5p | 2 |
| MR252 | miR-147b-5p | 1 | MR304 | 2 | |
| MR155 | miR-365-5p | 1 | |||
| MR195 | miR-511-3p | 1 | |||
| MR47 | miR-548au-3p | 1 | |||
| miR-548q | 1 | ||||
| miR-548v | 1 | ||||
| MR285 | miR-6715b-5p | 1 | |||
| MD197 | miR-6853-3p | 1 | |||
| MD235 | miR-105-3p | 2 | |||
| MD142 | miR-147b-5p | 2 | |||
| MR3 | miR-4471 | 2 | |||
| Hsa-miRNA | Tissue | Disease | Target Sequence | Score |
|---|---|---|---|---|
| miR-298 | Adrenal glans, Arteries, Heart, Lung | Alzheimer’s Disease | 5′UTR | 91 |
| miR-644a | Arteries, Brain, Cortex Cerebellum, Heart, Lung | NSP14 | 92 | |
| miR-4742-3p | Adrenal glands, Brain, Cerebellum Cortex, Heart, Liver, Lung, Kidney | NSP14 | 94 | |
| miR-21-3p | Arteries, Blood, Lung, Thyroid | Cholesteatoma, Glioma, Larynx cancer, Oral Squamous Cell Carcinoma, Tongue Squamous Cell Carcinoma | NSP15 | 94 |
| miR-219a-1-3p | Adrenal glands, Brain, Cortex Cerebellum, Heart, Kidney, Lung | CNS tumors, HCC, Infratentorial cancer, LC, Sacral cordoma | SPIKE | 85 |
| miR-624-5p | Brain, Cerebellum, Heart, Lung | SPIKE | 90 | |
| miR-148a-3p | Blood, Liver, Lymphnodes, Intestines | Asthma, CCA, ESCR | ORF7a | 94 |
| miR-148b-3p | Adrenal glands, Brain, Heart, Lung | Asthma, Oral Squamous Cell Carcinoma | ORF7a | 94 |
| miR-152-3p | Arteries, Colon, Pituitary gland, Thyroid | ALL, Asthma, CCA, HCC | ORF7a | 94 |
| miR-589-5p | Adrenal glands, Heart, Liver, Lung, Nervous tissue | ORF7b | 94 |
| 5′UTR | |||||
| Wuhan | Alpha | Beta | Gamma | Delta | Omicron |
| miR-7851-3p (91) | nd | nd | nd | nd | nd |
| SPIKE | |||||
| Wuhan | Alpha | Beta | Gamma | Delta | Omicron |
| miR-122b-3p (71) | miR-122b-3p (80) | miR-122b-3p (87) | miR-122b-3p (87) | miR-122b-3p (80) | miR-122b-3p (79) |
| miR-182-3p (57) | miR-182-3p (57) | miR-182-3p (57) | miR-182-3p (57) | miR-182-3p (90) | miR-182-3p (57) |
| miR-21-3p (71) | miR-21-3p (80) | miR-21-3p (87) | miR-21-3p (87) | miR-21-3p (80) | miR-21-3p (79) |
| miR-424-5p (84) | miR-424-5p (84) | miR-424-5p (84) | miR-424-5p (89) | miR-424-5p (84) | miR-424-5p (84) |
| miR-497-5p (84) | miR-497-5p (84) | miR-497-5p (84) | miR-497-5p (89) | miR-497-5p (84) | miR-497-5p (84) |
| miR-597-3p (81) | miR-597-3p (81) | miR-597-3p (81) | miR-597-3p (81) | miR-597-3p (88) | miR-597-3p (81) |
| miR-6838-5p (84) | miR-6835-5p (68) | miR-6835-5p (68) | miR-6838-5p (89) | miR-6838-3p (51) | miR-6835-5p (68) |
| Nucleocapsid | |||||
| Wuhan | Alpha | Beta | Gamma | Delta | Omicron |
| miR-615-5p (79) | miR-615-5p (89) | miR-615-5p (79) | miR-615-5p (79) | miR-615-5p (79) | miR-615-5p (80) |
| 3′UTR | |||||
| Wuhan | Alpha | Beta | Gamma | Delta | Omicron |
| miR-3941 (84) | miR-3941 (85) | miR-3941 (85) | miR-3941 (85) | miR-3941 (85) | miR-3941 (80) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battaglia, R.; Alonzo, R.; Pennisi, C.; Caponnetto, A.; Ferrara, C.; Stella, M.; Barbagallo, C.; Barbagallo, D.; Ragusa, M.; Purrello, M.; et al. MicroRNA-Mediated Regulation of the Virus Cycle and Pathogenesis in the SARS-CoV-2 Disease. Int. J. Mol. Sci. 2021, 22, 13192. https://doi.org/10.3390/ijms222413192
Battaglia R, Alonzo R, Pennisi C, Caponnetto A, Ferrara C, Stella M, Barbagallo C, Barbagallo D, Ragusa M, Purrello M, et al. MicroRNA-Mediated Regulation of the Virus Cycle and Pathogenesis in the SARS-CoV-2 Disease. International Journal of Molecular Sciences. 2021; 22(24):13192. https://doi.org/10.3390/ijms222413192
Chicago/Turabian StyleBattaglia, Rosalia, Ruben Alonzo, Chiara Pennisi, Angela Caponnetto, Carmen Ferrara, Michele Stella, Cristina Barbagallo, Davide Barbagallo, Marco Ragusa, Michele Purrello, and et al. 2021. "MicroRNA-Mediated Regulation of the Virus Cycle and Pathogenesis in the SARS-CoV-2 Disease" International Journal of Molecular Sciences 22, no. 24: 13192. https://doi.org/10.3390/ijms222413192