
Anti-Inflammatory Effects and Decreased Formation of Neutrophil Extracellular Traps by Enoxaparin in COVID-19 Patients

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Demographic Data
2.2. Increased NET Products in Plasma of COVID-19 Patients
2.3. Plasma of COVID-19 Patients Induced NETs
2.4. Reduction in NET and mRNA Expression by Enoxaparin Pretreatment In Vitro
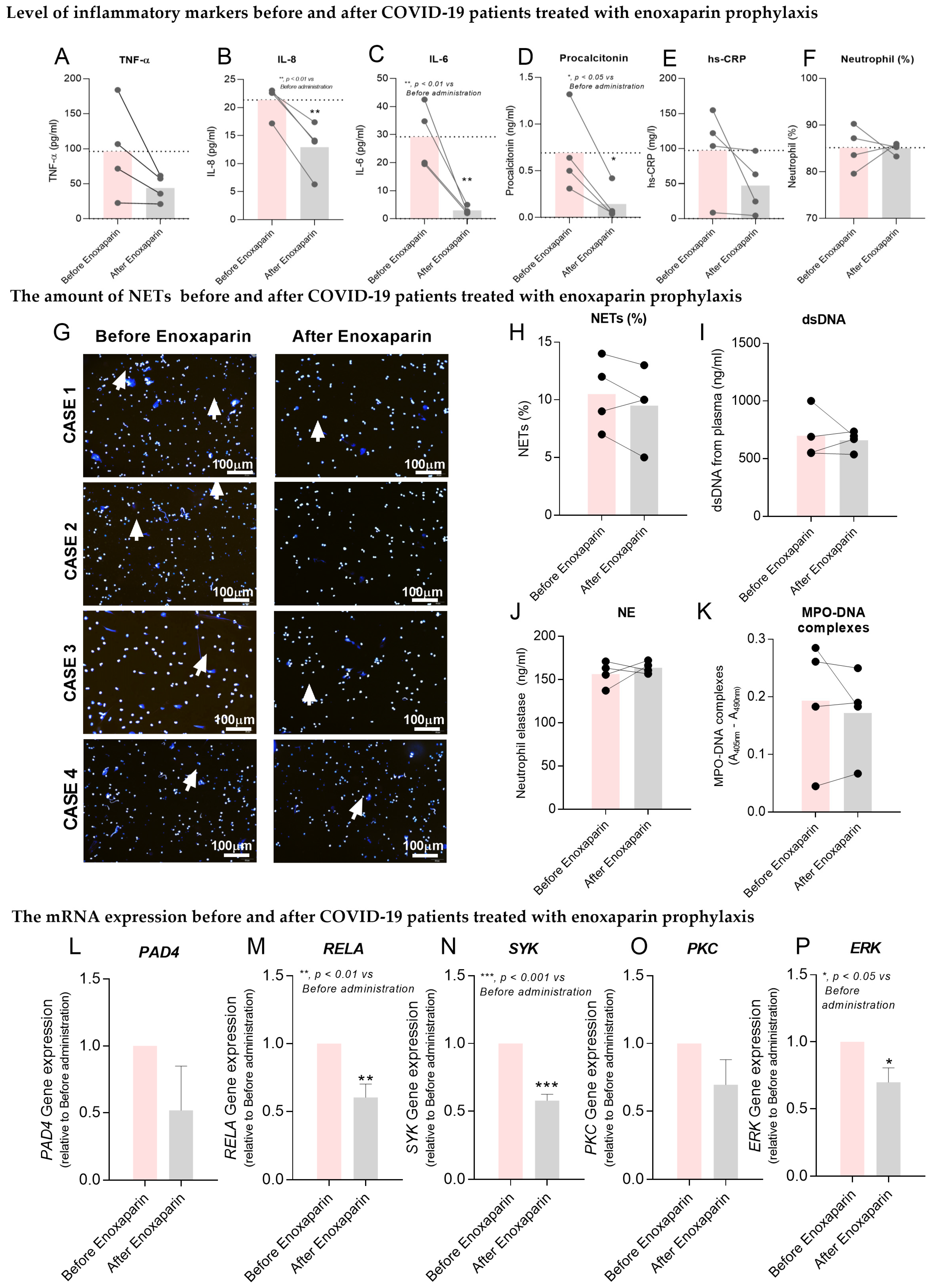
2.5. Effects of Enoxaparin Prophylaxis in COVID-19 Patients
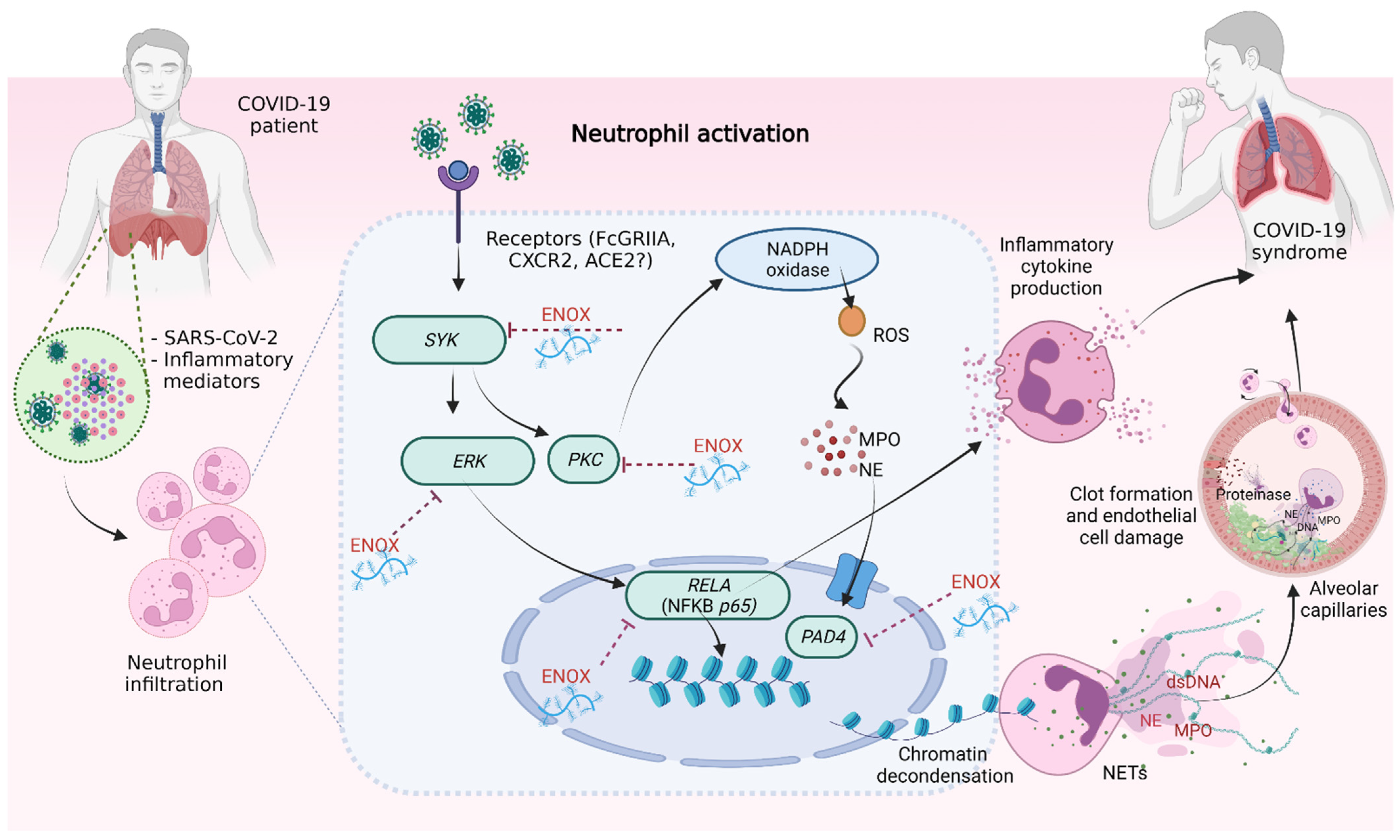
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Human Neutrophil Isolation
4.3. Detection of NETs
4.4. Quantification of NETs
4.5. Quantification of Cytokines and NET Products
4.6. Measurement of mRNA Expression
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scialo, F.; Daniele, A.; Amato, F.; Pastore, L.; Matera, M.G.; Cazzola, M.; Castaldo, G.; Bianco, A. ACE2: The Major Cell Entry Receptor for SARS-CoV-2. Lung 2020, 198, 867–877. [Google Scholar] [CrossRef]
- Beyerstedt, S.; Casaro, E.B.; Rangel, É.B. COVID-19: Angiotensin-converting enzyme 2 (ACE2) expression and tissue susceptibility to SARS-CoV-2 infection. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Attaway, A.H.; Scheraga, R.G.; Bhimraj, A.; Biehl, M.; Hatipoğlu, U. Severe COVID-19 pneumonia: Pathogenesis and clinical management. BMJ 2021, 372, n436. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Bai, X.; Wang, J.; Yu, Q.; Liu, W.; Pu, J. Clinical characteristics of coronavirus disease 2019 in Gansu province, China. Ann. Palliat. Med. 2020, 9, 1404–1412. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, Q.; Yin, Y.; Zhang, Y.; Cao, Y.; Lin, X. Excessive Neutrophils and Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2020, 11, 2063. [Google Scholar] [CrossRef] [PubMed]
- Moisa, E.; Corneci, D.; Negoita, S.; Filimon, C.; Serbu, A.; Negutu, M. Dynamic Changes of the Neutrophil-to-Lymphocyte Ratio, Systemic Inflammation Index, and Derived Neutrophil-to-Lymphocyte Ratio Independently Predict Invasive Mechanical Ventilation Need and Death in Critically Ill COVID-19 Patients. Biomedicines 2021, 9, 1656. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.M.; Ahmad, U.; Usman, U.; Ali, M.; Shaukat, A.; Gul, N. Neutrophil/lymphocyte ratio-A marker of COVID-19 pneumonia severity. Int. J. Clin. Pract. 2021, 75, e13698. [Google Scholar] [CrossRef]
- Twaddell, S.H.; Baines, K.J.; Grainge, C.; Gibson, P.G. The Emerging Role of Neutrophil Extracellular Traps in Respiratory Disease. Chest 2019, 156, 774–782. [Google Scholar] [CrossRef]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.A. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. Am. J. Pathol. 2011, 179, 199–210. [Google Scholar] [CrossRef]
- Bendib, I.; de Chaisemartin, L.; Granger, V.; Schlemmer, F.; Maitre, B.; Hüe, S. Neutrophil Extracellular Traps Are Elevated in Patients with Pneumonia-related Acute Respiratory Distress Syndrome. Anesthesiology 2019, 130, 581–591. [Google Scholar] [CrossRef]
- Connors, J.M.; Levy, J.H. COVID-19 and its implications for thrombosis and anticoagulation. Blood 2020, 135, 2033–2040. [Google Scholar] [CrossRef]
- Cuker, A.; Tseng, E.K.; Nieuwlaat, R.; Angchaisuksiri, P.; Blair, C.; Dane, K. American Society of Hematology 2021 guidelines on the use of anticoagulation for thromboprophylaxis in patients with COVID-19. Blood Adv. 2021, 5, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Drago, F.; Gozzo, L.; Li, L.; Stella, A.; Cosmi, B. Use of Enoxaparin to Counteract COVID-19 Infection and Reduce Thromboembolic Venous Complications: A Review of the Current Evidence. Front. Pharmacol. 2020, 11, 9886. [Google Scholar] [CrossRef] [PubMed]
- Mycroft-West, C.J.; Su, D.; Pagani, I.; Rudd, T.R.; Elli, S.; Gandhi, N.S. Heparin Inhibits Cellular Invasion by SARS-CoV-2: Structural Dependence of the Interaction of the Spike S1 Receptor-Binding Domain with Heparin. Thromb. Haemost. 2020, 120, 1700–1715. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, A.A.; Rovere-Querini, P.; D’Angelo, A.; Maugeri, N. Low molecular weight heparins prevent the induction of autophagy of activated neutrophils and the formation of neutrophil extracellular traps. Pharm. Res. 2017, 123, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, J.; Gao, M.; Fan, H.; Wang, Y.; Xu, X. Interleukin-8 as a Biomarker for Disease Prognosis of Coronavirus Disease-2019 Patients. Front. Immunol. 2021, 11, 602395. [Google Scholar] [CrossRef]
- Alfaro, C.; Teijeira, A.; Oñate, C.; Pérez, G.; Sanmamed, M.F.; Andueza, M.P. Tumor-Produced Interleukin-8 Attracts Human Myeloid-Derived Suppressor Cells and Elicits Extrusion of Neutrophil Extracellular Traps (NETs). Clin. Cancer Res. 2016, 22, 3924–3936. [Google Scholar] [CrossRef] [Green Version]
- Arcanjo, A.; Logullo, J.; Menezes, C.C.B.; de Souza Carvalho Giangiarulo, T.C.; dos Reis, M.C.; de Castro, G.M.M. The emerging role of neutrophil extracellular traps in severe acute respiratory syndrome coronavirus 2 (COVID-19). Sci. Rep. 2020, 10, 19630. [Google Scholar] [CrossRef]
- World Health Organization. COVID-19 Clinical Management: Living Guidance, 25 January 2021; World Health Organization: Geneva, Switzerland, 2021; Contract No.: WHO/2019-nCoV/clinical/2021.1. [Google Scholar]
- Laforge, M.; Elbim, C.; Frère, C.; Hémadi, M.; Massaad, C.; Nuss, P. Tissue damage from neutrophil-induced oxidative stress in COVID-19. Nat. Rev. Immunol. 2020, 20, 515–516. [Google Scholar] [CrossRef]
- Azzouz, D.; Khan, M.A.; Palaniyar, N. ROS induces NETosis by oxidizing DNA and initiating DNA repair. Cell Death Discov. 2021, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshari, R.S.; Jyoti, A.; Dubey, M.; Kothari, N.; Kohli, M.; Bogra, J. Cytokines induced neutrophil extracellular traps formation: Implication for the inflammatory disease condition. PLoS ONE 2012, 7, e48111. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Arfman, T.; Wichapong, K.; Reutelingsperger, C.P.M.; Voorberg, J.; Nicolaes, G.A.F. PAD4 takes charge during neutrophil activation: Impact of PAD4 mediated NET formation on immune-mediated disease. J. Thromb. Haemost. 2021, 19, 1607–1617. [Google Scholar] [CrossRef]
- Strich, J.R.; Ramos-Benitez, M.J.; Randazzo, D.; Stein, S.R.; Babyak, A.; Davey, R.T. Fostamatinib Inhibits Neutrophils Extracellular Traps Induced by COVID-19 Patient Plasma: A Potential Therapeutic. J. Infect. Dis. 2020, 223, 981–984. [Google Scholar] [CrossRef]
- Dowey, R.; Cole, J.; Thompson, A.A.R.; Huang, C.; Whatmore, J.; Iqbal, A. Enhanced neutrophil extracellular trap formation in COVID-19 is inhibited by the PKC inhibitor ruboxistaurin. ERJ Open Res. 2022, 8, 00596-2021. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Fang, H.; Dang, E.; Xue, K.; Zhang, J.; Li, B. Neutrophil Extracellular Traps Promote Inflammatory Responses in Psoriasis via Activating Epidermal TLR4/IL-36R Crosstalk. Front. Immunol. 2019, 10, 746. [Google Scholar] [CrossRef] [Green Version]
- Hakkim, A.; Fuchs, T.A.; Martinez, N.E.; Hess, S.; Prinz, H.; Zychlinsky, A. Activation of the Raf-MEK-ERK pathway is required for neutrophil extracellular trap formation. Nat. Chem. Biol. 2011, 7, 75–77. [Google Scholar] [CrossRef]
- Hu, R.; Han, C.; Pei, S.; Yin, M.; Chen, X. Procalcitonin levels in COVID-19 patients. Int. J. Antimicrob. Agents 2020, 56, 106051. [Google Scholar] [CrossRef]
- Smilowitz, N.R.; Kunichoff, D.; Garshick, M.; Shah, B.; Pillinger, M.; Hochman, J.S. C-reactive protein and clinical outcomes in patients with COVID-19. Eur. Heart J. 2021, 42, 2270–2279. [Google Scholar] [CrossRef]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Polverino, E.; Rosales-Mayor, E.; Dale, G.E.; Dembowsky, K.; Torres, A. The Role of Neutrophil Elastase Inhibitors in Lung Diseases. Chest 2017, 152, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengas, A.; Poletti, V.; Pacifico, L.; di Domizio, C.; Patelli, M.; Spiga, L. Acute lung inflammation: Neutrophil elastase versus neutrophils in the bronchoalveolar lavage--neutrophil elastase reflects better inflammatory intensity. Intensive Care Med. 1994, 20, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, B.A.; Colovos, C.; Lawson, M.A.; Osborn, Z.T.; Sackheim, A.M.; Mould, K.J. Increased Histone-DNA complexes and endothelial-dependent thrombin generation in severe COVID-19. Vascul. Pharmacol. 2021, 142, 106950. [Google Scholar] [CrossRef]
- Cortjens, B.; de Boer, O.J.; de Jong, R.; Antonis, A.F.; Sabogal Piñeros, Y.S.; Lutter, R. Neutrophil extracellular traps cause airway obstruction during respiratory syncytial virus disease. J. Pathol. 2016, 238, 401–411. [Google Scholar] [CrossRef]
- Zuo, Y.; Estes, S.K.; Ali, R.A.; Gandhi, A.A.; Yalavarthi, S.; Shi, H. Prothrombotic autoantibodies in serum from patients hospitalized with COVID-19. Sci. Transl. Med. 2020, 12, 570. [Google Scholar] [CrossRef]
- Chen, T.; Li, Y.; Sun, R.; Hu, H.; Liu, Y.; Herrmann, M. Receptor-Mediated NETosis on Neutrophils. Front. Immunol. 2021, 12, 775267. [Google Scholar] [CrossRef]
- Nijhuis, R.H.T.; Russcher, A.; de Jong, G.J.; Jong, E.; Herder, G.J.M.; Remijn, J.A. Low prevalence of SARS-CoV-2 in plasma of COVID-19 patients presenting to the emergency department. J. Clin. Virol. 2020, 133, 104655. [Google Scholar] [CrossRef]
- Sae-Khow, K.; Tachaboon, S.; Wright, H.L.; Edwards, S.W.; Srisawat, N.; Leelahavanichkul, A. Defective Neutrophil Function in Patients with Sepsis Is Mostly Restored by ex vivo Ascorbate Incubation. J. Inflamm. Res. 2020, 13, 263–274. [Google Scholar] [CrossRef]
- Caudrillier, A.; Kessenbrock, K.; Gilliss, B.M.; Nguyen, J.X.; Marques, M.B.; Monestier, M. Platelets induce neutrophil extracellular traps in transfusion-related acute lung injury. J. Clin. Investig. 2012, 122, 2661–2671. [Google Scholar] [CrossRef]
- Robinson, S.; Zincuk, A.; Strøm, T.; Larsen, T.B.; Rasmussen, B.; Toft, P. Enoxaparin, effective dosage for intensive care patients: Double-blinded, randomised clinical trial. Crit. Care 2010, 14, R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawes, J. Comparison of the pharmacokinetics of enoxaparin (Clexane) and unfractionated heparin. Acta Chir. Scand. Suppl. 1990, 556, 68–74. [Google Scholar] [PubMed]
- Saisorn, W.; Saithong, S.; Phuengmaung, P.; Udompornpitak, K.; Bhunyakarnjanarat, T.; Visitchanakun, P. Acute Kidney Injury Induced Lupus Exacerbation Through the Enhanced Neutrophil Extracellular Traps (and Apoptosis) in Fcgr2b Deficient Lupus Mice With Renal Ischemia Reperfusion Injury. Front. Immunol. 2021, 12, 669162. [Google Scholar] [CrossRef] [PubMed]
- Saithong, S.; Saisorn, W.; Visitchanakun, P.; Sae-Khow, K.; Chiewchengchol, D.; Leelahavanichkul, A. A Synergy Between Endotoxin and (1→3)-Beta-D-Glucan Enhanced Neutrophil Extracellular Traps in Candida Administered Dextran Sulfate Solution Induced Colitis in FcGRIIB-/- Lupus Mice, an Impact of Intestinal Fungi in Lupus. J. Inflamm. Res. 2021, 14, 2333–2352. [Google Scholar] [CrossRef]
- Wright, H.L.; Makki, F.A.; Moots, R.J.; Edwards, S.W. Low-density granulocytes: Functionally distinct, immature neutrophils in rheumatoid arthritis with altered properties and defective TNF signalling. J. Leukoc. Biol. 2017, 101, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Krumbholz, M.; Schönermarck, U.; Back, W.; Gross, W.L.; Werb, Z. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, 11. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Mild (n = 10) | Moderate (n = 10) | Severe (n = 10) | p-Value |
|---|---|---|---|---|
| Gender (Female) | 42.6% | 33.3% | 16.7% | NA |
| (NA) | (NA) | (NA) | ||
| Mean Age (yr) | 32.71 | 50.00 | 48.67 | 0.29 |
| (16–49) | (34–71) | (18–84) | ||
| Mean Weight (kg) | 62.08 | 69.00 | 68.70 | 0.74 |
| (49–72) | (60–80) | (85–56) | ||
| Mean BMI (kg/m2) | 22.39 | 25.72 | 26.76 | 0.34 |
| (17.78–24.34) | (24.03–27.36) | (23.31–31.22) | ||
| Underlying diseases | ||||
| Diabetes | 0 | 2 | 1 | 0.263 |
| Hypertension | 0 | 1 | 1 | 0.521 |
| Dyslipidemia | 0 | 0 | 1 | 0.319 |
| Chronic kidney disease | 0 | 0 | 2 | 0.089 |
| Others (e.g., allergy) | 0 | 0 | 0 | 0.298 |
| Parameter | Mild (n = 10) | Moderate (n = 10) | Severe (n = 10) | p-Value |
|---|---|---|---|---|
| Red cell count | 5.08 | 4.82 | 5.05 | 0.26 |
| (4.59–5.81) | (4.08–5.35) | (4.11–6.55) | ||
| Hemoglobin | 13.41 | 13.58 | 13.33 | 0.78 |
| (10.2–15.2) | (10.3–15.2) | (12.2–15.5) | ||
| Hematocrit | 40.80 | 40.45 | 39.48 | 0.84 |
| (38.6–45.9) | (32.4–46.9) | (33.6–45.8) | ||
| MCV | 80.34 | 83.97 | 78.95 | 0.33 |
| (66.7–90.8) | (78.2–88.7) | (69.9–84.6) | ||
| MCH | 26.41 | 28.17 | 26.75 | 0.88 |
| (22.2–29.3) | (25.2–31.8) | (23.4–29.7) | ||
| MCHC | 32.87 | 33.60 | 33.85 | 0.01 |
| (32.3–33.3) | (31.8–36.3) | (31.0–36.3) | ||
| RDW | 12.94 | 12.62 | 12.87 | 0.96 |
| (11.6–14.4) | (11.0–14.0) | (12.1–15.2) | ||
| White cell count | 4428.57 | 4795.00 | 5628.33 | 0.15 |
| (2490–7550) | (3410–7650) | (3460–8220) | ||
| Neutrophil% | 46.34 | 60.63 | 64.48 | 0.30 |
| (34.8–58.8) | (45.3–82.1) | (37.8–77.7) | ||
| Neutrophil # | 2070.00 | 2913.33 | 3786.67 | 0.26 |
| (1230–3720) | (1960–5200) | (1310–6390) | ||
| Lymphocyte% | 44.14 | 30.82 | 26.42 | 0.17 |
| (31.8–52.0) | (10.6–44.6) | (9.6–53.2) | ||
| Lymphocyte # | 1918.57 | 1448.33 | 1353.33 | 0.82 |
| (1050–3260) | (360–2110) | (450–1840) | ||
| NLR | 1.12 | 2.86 | 3.58 | 0.07 |
| (0.67–1.85) | (1.02–7.78) | (0.71–8.09) | ||
| Monocyte % | 7.67 | 7.75 | 8.53 | 0.40 |
| (5.4–9.9) | (4.7–10.3) | (5.4–9.4) | ||
| Monocyte # | 331.43 | 385.00 | 465.00 | 0.14 |
| (170–410) | (220–790) | (290–730) | ||
| Eosinophils % | 1.46 | 0.53 | 0.23 | 0.10 |
| (0–2.8) | (0–1.7) | (0–0.8) | ||
| Eosinophils # | 65.71 | 33.33 | 11.67 | 0.05 |
| (0–130) | (0–130) | (0–40) | ||
| Basophils % | 0.39 | 0.27 | 0.18 | 0.50 |
| (0–0.7) | (0–0.7) | (0–0.4) | ||
| Basophils # | 14.29 | 15.00 | 10.00 | 0.47 |
| (0–30) | (0–50) | (0–30) | ||
| Platelet count | 205,857.14 | 207,500.00 | 199,666.67 | 0.30 |
| (34,000–327,000) | (138,000–298,000) | (144,000–282,000) | ||
| MPV | 10.68 | 10.70 | 11.32 | 0.52 |
| (10.3–12.1) | (9.3–11.8) | (10.1–13.3) |
| Parameter | Mild (n = 10) | Moderate (n = 10) | Severe (n = 10) | p-Value |
|---|---|---|---|---|
| AST | 18.00 | 31.83 | 62.00 | 0.004 |
| (12–25) | (18–68) | (26–182) | ||
| ALT | 15.86 | 39.83 | 45.00 | 0.04 |
| (6–38) | (19–81) | (13–109) | ||
| ALP | 57.71 | 66.50 | 73.33 | 0.22 |
| (45–97) | (89–43) | (37–137) | ||
| Total billirubin | 0.52 | 0.57 | 0.75 | 1.00 |
| (0.29–1.05) | (0.28–0.91) | (0.53–1.06) | ||
| Direct billirubin | 0.20 | 0.23 | 0.36 | 0.24 |
| (0.11–0.37) | (0.11–0.39) | (0.18–0.67) | ||
| Serum total protein | 8.15 | 7.80 | 8.00 | 0.98 |
| (7.9–8.4) | (7.6–8.0) | (7.8–8.4) | ||
| Serum albumin | 4.30 | 3.98 | 3.77 | 0.40 |
| (4.1–4.7) | (3.6–4.3) | (3.1–4.4) | ||
| Serum creatinine | 0.89 | 0.78 | 1.34 | 0.04 |
| (0.61–1.39) | (0.50–0.96) | (0.81–2.20) | ||
| Blood urea nitrogen | 12.57 | 10.50 | 14.33 | 0.62 |
| (7–20) | (7–14) | (10–22) | ||
| serum IL-6 | NA | NA | 1680 | NA |
| (NA) | (NA) | 1680 | ||
| serum hs-CRP | NA | 43.61 | 102.85 | 0.14 |
| (NA) | (16.95–110.32) | (21.38–230.37) | ||
| serum procalcitonin | NA | 0.05 | 0.28 | 0.08 |
| (NA) | (0.02–0.12) | (0.05–1.01) | ||
| serum ferritin | NA | 687.65 | 1464.24 | 1.00 |
| (NA) | (346.5–884.1) | (620–4609) | ||
| serum LDH | NA | 242.33 | 375.33 | 0.23 |
| (NA) | (200–299) | (198–556) | ||
| d Na | 139.33 | 137.67 | 136.83 | 0.38 |
| (136–142) | (133–142) | (129–141) | ||
| d K | 3.55 | 3.43 | 3.42 | 0.88 |
| (3.2–3.9) | (3.0–3.8) | (2.7–3.8) | ||
| d Cl | 106.33 | 102.50 | 90.17 | 0.07 |
| (104–109) | (99–107) | (21–106) | ||
| d HCO3 | 25.17 | 25.33 | 21.67 | 0.03 |
| (23–28) | (23–27) | (12–26) |
| Parameter | Mild (n = 10) | Moderate (n = 10) | Severe (n = 10) | p-Value |
|---|---|---|---|---|
| D-Dimer | NA | 364.36 | 1290.29 | 0.02 |
| (NA) | (308.95–411.6) | (794.55–1884.02) | ||
| PT | NA | 12.83 | 13.20 | 0.01 |
| (NA) | (11.9–14.3) | (12.8–13.5) | ||
| INR | NA | 1.12 | 1.15 | 0.01 |
| (NA) | (1.04–1.25) | (1.12–1.18) | ||
| APTT | NA | 25.30 | 31.40 | 0.30 |
| (NA) | (23.0–27.3) | (23.4–56.3) | ||
| Fibrinogen | NA | 5.22 | 4.66 | 0.41 |
| (NA) | (4.86–5.91) | (4.02–5.91) |
| Target Genes | Forward Primers | Reverse Primers |
|---|---|---|
| PAD4 | 5′-CGAAGACCCCCAAGGACT-3′ | 5′-AGGACAGTTTGCCCCGTG-3′ |
| NFKB | 5′-GCAGCACTACTTCTTGACCACC-3′ | 5′-TCTGCTCCTGAGCATTGACGTC-3′ |
| SYK | 5′-CGTATGAGCCAGAACTTGCACC-3′ | 5′-CTTTCGGTCCAGGTAAACCTCC-3′ |
| PKC | 5′-GAGGGACACATCAAGATTGCCG-3′ | 5′-CACCAATCCACGGACTTCCCAT-3′ |
| ERK | 5′-TGGCAAGCACTACCTGGATCAG-3′ | 5′-GCAGAGACTGTAGGTAGTTTCGG-3′ |
| ACTB | 5′-CACCATTGGCAATGAGCGGTTC-3′ | 5′-AGGTCTTTGCGGATGTCCACGT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saithong, S.; Saisorn, W.; Tovichayathamrong, P.; Filbertine, G.; Torvorapanit, P.; Wright, H.L.; Edwards, S.W.; Leelahavanichkul, A.; Hirankarn, N.; Chiewchengchol, D. Anti-Inflammatory Effects and Decreased Formation of Neutrophil Extracellular Traps by Enoxaparin in COVID-19 Patients. Int. J. Mol. Sci. 2022, 23, 4805. https://doi.org/10.3390/ijms23094805
Saithong S, Saisorn W, Tovichayathamrong P, Filbertine G, Torvorapanit P, Wright HL, Edwards SW, Leelahavanichkul A, Hirankarn N, Chiewchengchol D. Anti-Inflammatory Effects and Decreased Formation of Neutrophil Extracellular Traps by Enoxaparin in COVID-19 Patients. International Journal of Molecular Sciences. 2022; 23(9):4805. https://doi.org/10.3390/ijms23094805
Chicago/Turabian StyleSaithong, Supichcha, Wilasinee Saisorn, Punyot Tovichayathamrong, Grace Filbertine, Pattama Torvorapanit, Helen L. Wright, Steven W. Edwards, Asada Leelahavanichkul, Nattiya Hirankarn, and Direkrit Chiewchengchol. 2022. "Anti-Inflammatory Effects and Decreased Formation of Neutrophil Extracellular Traps by Enoxaparin in COVID-19 Patients" International Journal of Molecular Sciences 23, no. 9: 4805. https://doi.org/10.3390/ijms23094805