
Pathogenicity Islands in Uropathogenic Escherichia coli Clinical Isolate of the Globally Disseminated O25:H4-ST131 Pandemic Clonal Lineage: First Report from Egypt

Abstract
:1. Introduction
2. Results and Discussion
2.1. Antimicrobial Susceptibility Profile
2.2. Genomic Analysis of UPEC Strain EC14142
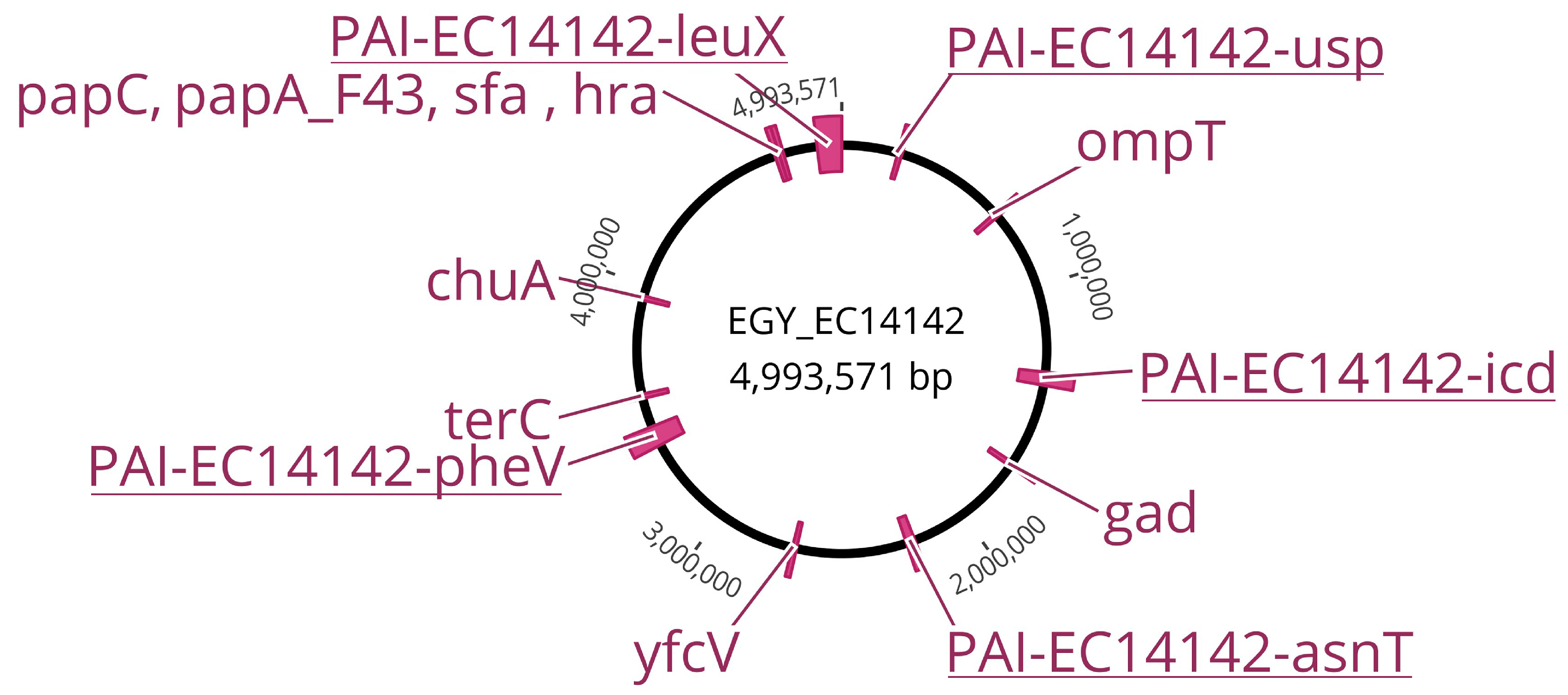
2.3. Distribution of Virulence Genes on EGY_EC14142
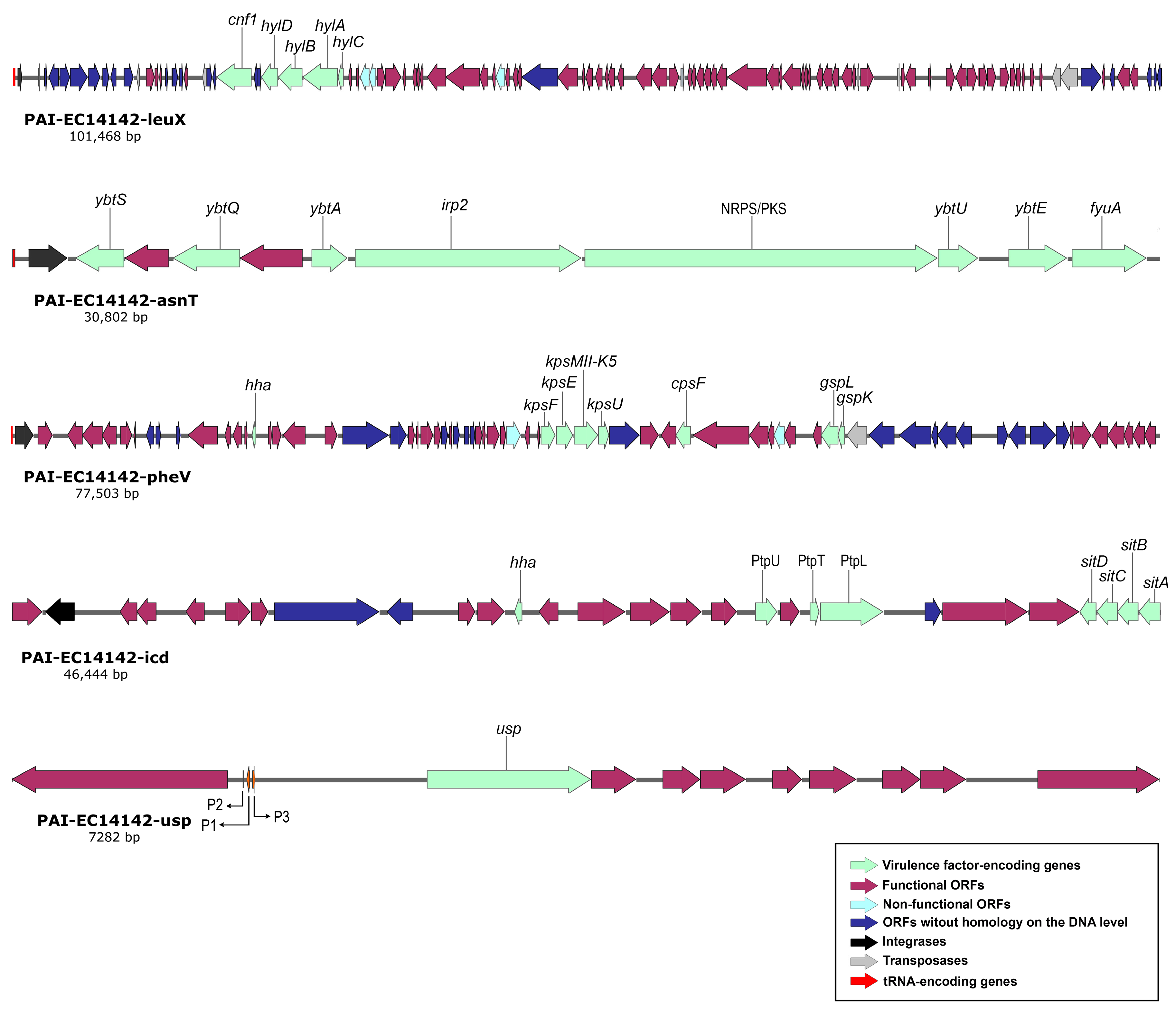
2.4. Structural Features of Pathogenicity Islands Integrated into EGY_EC14142 Chromosome
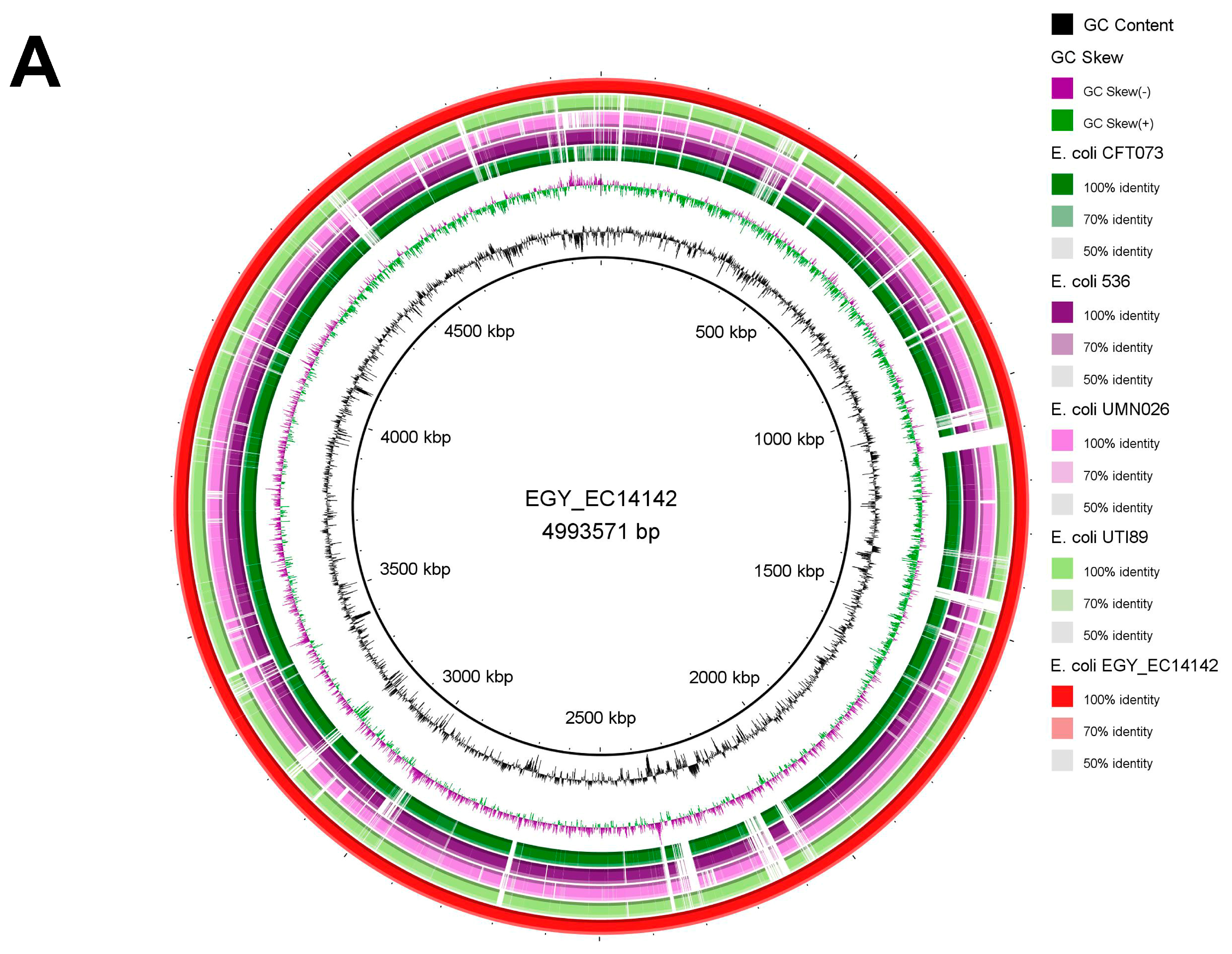
2.5. Genomic Comparisons of EGY_EC14142 with Closely Related UPEC Strains
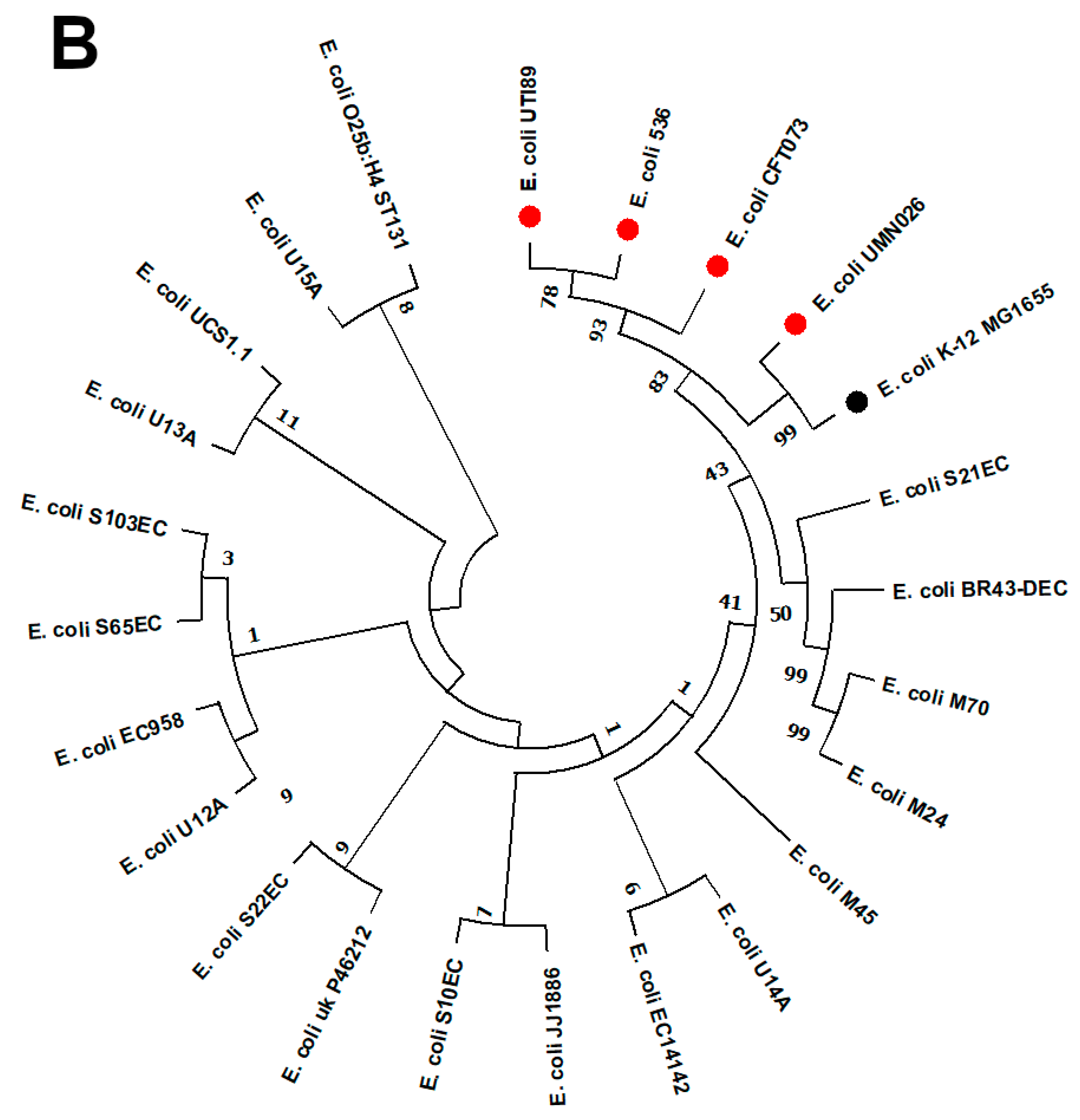
2.6. Phylogenomic Analysis of EC14142 Related to E. coli Belonging to ST131 Clade C
3. Materials and Methods
3.1. E. coli EC14142 Strain Isolation and Identification
3.2. Antimicrobial Susceptibility Testing
3.3. DNA Extraction and Whole Genome Sequencing (WGS)
3.4. Construction of EGY_EC14142 Chromosome and Identification of Its PAIs
3.5. Genomic Similarity and Phylogenetic Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bien, J.; Sokolova, O.; Bozko, P. Role of uropathogenic Escherichia coli virulence factors in development of urinary tract infection and kidney damage. Int. J. Nephrol. 2012, 2012, 681473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, H.; Hensel, M. Pathogenicity islands in bacterial pathogenesis. Clin. Microbiol. Rev. 2004, 17, 14–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelschlaeger, T.A.; Dobrindt, U.; Hacker, J. Pathogenicity islands of uropathogenic E. coli and the evolution of virulence. Int. J. Antimicrob. Agents 2002, 19, 517–521. [Google Scholar] [CrossRef]
- Middendorf, B.; Blum-Oehler, G.; Dobrindt, U.; Muhldorfer, I.; Salge, S.; Hacker, J. The pathogenicity islands (PAIs) of the uropathogenic Escherichia coli strain 536: Island probing of PAI II536. J. Infect. Dis. 2001, 183, S17–S20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacker, J.; Bender, L.; Ott, M.; Wingender, J.; Lund, B.; Marre, R.; Goebel, W. Deletions of chromosomal regions coding for fimbriae and hemolysins occur in vitro and in vivo in various extraintestinal Escherichia coli isolates. Microb. Pathog. 1990, 8, 213–225. [Google Scholar] [CrossRef]
- Gal-Mor, O.; Finlay, B.B. Pathogenicity islands: A molecular toolbox for bacterial virulence. Cell. Microbiol. 2006, 8, 1707–1719. [Google Scholar] [CrossRef]
- Desvaux, M.; Dalmasso, G.; Beyrouthy, R.; Barnich, N.; Delmas, J.; Bonnet, R. Pathogenicity factors of genomic islands in intestinal and extraintestinal Escherichia coli. Front. Microbiol. 2020, 11, 2065. [Google Scholar] [CrossRef]
- Schneider, G.; Dobrindt, U.; Bruggemann, H.; Nagy, G.; Janke, B.; Blum-Oehler, G.; Buchrieser, C.; Gottschalk, G.; Emödy, L.; Hacker, J. The pathogenicity island-associated K15 capsule determinant exhibits a novel genetic structure and correlates with virulence in uropathogenic Escherichia coli strain 536. Infect. Immun. 2004, 72, 5993–6001. [Google Scholar] [CrossRef] [Green Version]
- Brzuszkiewicz, E.; Bruggemann, H.; Liesegang, H.; Emmerth, M.; Olschlager, T.; Nagy, G.; Albermann, K.; Wagner, C.; Buchrieser, C.; Emődy, L.; et al. How to become a uropathogen: Comparative genomic analysis of extraintestinal pathogenic Escherichia coli strains. Proc. Natl. Acad. Sci. USA 2006, 103, 12879–12884. [Google Scholar] [CrossRef] [Green Version]
- Zerr, D.M.; Miles-Jay, A.; Kronman, M.P.; Zhou, C.; Adler, A.L.; Haaland, W.; Weissman, S.J.; Elward, A.; Newland, J.G.; Zaoutis, T.; et al. Previous antibiotic exposure increases risk of infection with extended-spectrum-beta-lactamase- and AmpC-producing Escherichia coli and Klebsiella pneumoniae in pediatric patients. Antimicrob. Agents Chemother. 2016, 60, 4237–4243. [Google Scholar] [CrossRef]
- Moxon, C.A.; Paulus, S. Beta-lactamases in Enterobacteriaceae infections in children. J. Infect. 2016, 72, S41–S49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakaria, A.S.; Edward, E.A.; Mohamed, N.M. Genomic insights into a colistin-resistant uropathogenic Escherichia coli strain of O23:H4-ST641 lineage harboring mcr-1.1 on a conjugative IncHI2 plasmid from Egypt. Microorganisms 2021, 9, 799. [Google Scholar] [CrossRef] [PubMed]
- Fam, N.; Leflon-Guibout, V.; Fouad, S.; Aboul-Fadl, L.; Marcon, E.; Desouky, D.; El-Defrawy, I.; Abou-Aitta, A.; Klena, J.; Nicolas-Chanoine, M. CTX-M-15-producing Escherichia coli clinical isolates in Cairo (Egypt), including isolates of clonal complex ST10 and clones ST131, ST73, and ST405 in both community and hospital settings. Microb. Drug Resist. 2011, 17, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Cerero, L.; Navarro, M.D.; Bellido, M.; Martin-Pena, A.; Vinas, L.; Cisneros, J.M.; Gómez-Langley, S.L.; Sánchez-Monteseirín, H.; Morales, I.; Pascual, A.; et al. Escherichia coli belonging to the worldwide emerging epidemic clonal group O25b/ST131: Risk factors and clinical implications. J. Antimicrob. Chemother. 2014, 69, 809–814. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, R.; Johnston, B.; Lohse, C.; Porter, S.B.; Clabots, C.; Johnson, J.R. Escherichia coli sequence type 131 is a dominant, antimicrobial-resistant clonal group associated with healthcare and elderly hosts. Infect. Control Hosp. Epidemiol. 2013, 34, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.C.; Lai, C.H.; Lin, J.N.; Huang, C.K.; Liang, S.H.; Chen, W.F.; Shih, Y.C.; Lin, H.H.; Wang, J.L. Bacteremia caused by extended-spectrum-beta-lactamase-producing Escherichia coli sequence type ST131 and non-ST131 clones: Comparison of demographic data, clinical features, and mortality. Antimicrob. Agents Chemother. 2012, 56, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Abdelrahim, S.S.; Fouad, M.; Abdallah, N.; Ahmed, R.F.; Zaki, S. Comparative study of CTX-M-15 producing Escherichia coli ST131 clone isolated from urinary tract infections and acute diarrhoea. Infect. Drug Resist. 2021, 14, 4027–4038. [Google Scholar] [CrossRef]
- Paniagua-Contreras, G.L.; Hernandez-Jaimes, T.; Monroy-Perez, E.; Vaca-Paniagua, F.; Diaz-Velasquez, C.; Uribe-Garcia, A.; Vaca, S. Comprehensive expression analysis of pathogenicity genes in uropathogenic Escherichia coli strains. Microb. Pathog. 2017, 103, 1–7. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Rezatofighi, S.E.; Mirzarazi, M.; Salehi, M. Virulence genes and phylogenetic groups of uropathogenic Escherichia coli isolates from patients with urinary tract infection and uninfected control subjects: A case-control study. BMC Infect. Dis. 2021, 21, 361. [Google Scholar] [CrossRef]
- Ali, I.; Rafaque, Z.; Ahmed, I.; Tariq, F.; Graham, S.E.; Salzman, E.; Foxman, B.; Dasti, J.I. Phylogeny, sequence-typing and virulence profile of uropathogenic Escherichia coli (UPEC) strains from Pakistan. BMC Infect. Dis. 2019, 19, 620. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Tchesnokova, V.; Johnston, B.; Clabots, C.; Roberts, P.L.; Billig, M.; Riddell, K.; Rogers, P.; Qin, X.; Butler-Wu, S.; et al. Abrupt emergence of a single dominant multidrug-resistant strain of Escherichia coli. J. Infect. Dis. 2013, 207, 919–928. [Google Scholar] [CrossRef]
- Petty, N.K.; Ben Zakour, N.L.; Stanton-Cook, M.; Skippington, E.; Totsika, M.; Forde, B.M.; Phan, M.D.; Gomes Moriel, D.; Peters, K.M.; Davies, M.; et al. Global dissemination of a multidrug resistant Escherichia coli clone. Proc. Natl. Acad. Sci. USA 2014, 111, 5694–5699. [Google Scholar] [CrossRef] [Green Version]
- Schembri, M.A.; Zakour, N.L.; Phan, M.D.; Forde, B.M.; Stanton-Cook, M.; Beatson, S.A. Molecular characterization of the multidrug resistant Escherichia coli ST131 clone. Pathogens 2015, 4, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Cummins, E.A.; Snaith, A.E.; McNally, A.; Hall, R.J. The role of potentiating mutations in the evolution of pandemic Escherichia coli clones. Eur. J. Clin. Microbiol. Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Yazdanpour, Z.; Tadjrobehkar, O.; Shahkhah, M. Significant association between genes encoding virulence factors with antibiotic resistance and phylogenetic groups in community acquired uropathogenic Escherichia coli isolates. BMC Microbiol 2020, 20, 241. [Google Scholar] [CrossRef]
- Schwan, W.R.; Flohr, N.L.; Multerer, A.R.; Starkey, J.C. GadE regulates fliC gene transcription and motility in Escherichia coli. World J. Clin. Infect. Dis. 2020, 10, 14–23. [Google Scholar] [CrossRef]
- Miajlovic, H.; Smith, S.G. Bacterial self-defence: How Escherichia coli evades serum killing. FEMS Microbiol. Lett. 2014, 354, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.H.; Kikuchi, T.; Tokunaga, T.; Iyoda, S.; Iguchi, A. Diversity of the tellurite resistance gene operon in Escherichia coli. Front. Microbiol. 2021, 12, 681175. [Google Scholar] [CrossRef]
- Hochhut, B.; Wilde, C.; Balling, G.; Middendorf, B.; Dobrindt, U.; Brzuszkiewicz, E.; Gottschalk, G.; Carniel, E.; Hacker, J. Role of pathogenicity island-associated integrases in the genome plasticity of uropathogenic Escherichia coli strain 536. Mol. Microbiol. 2006, 61, 584–595. [Google Scholar] [CrossRef]
- Koh, E.I.; Robinson, A.E.; Bandara, N.; Rogers, B.E.; Henderson, J.P. Copper import in Escherichia coli by the yersiniabactin metallophore system. Nat. Chem. Biol. 2017, 13, 1016–1021. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, S.; Palur, R.V.; Sudhakar, K.U.; Rathinavelan, T. E. coli group 1 capsular polysaccharide exportation nanomachinary as a plausible antivirulence target in the perspective of emerging antimicrobial resistance. Front. Microbiol. 2017, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, E.; Nadal, I.; Donat, E.; Ribes-Koninckx, C.; Calabuig, M.; Sanz, Y. Reduced diversity and increased virulence-gene carriage in intestinal enterobacteria of coeliac children. BMC Gastroenterol. 2008, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Lallemand, M.; Login, F.H.; Guschinskaya, N.; Pineau, C.; Effantin, G.; Robert, X.; Shevchik, V.E. Dynamic interplay between the periplasmic and transmembrane domains of GspL and GspM in the type II secretion system. PLoS ONE 2013, 8, e79562. [Google Scholar] [CrossRef] [Green Version]
- Paytubi, S.; Dietrich, M.; Queiroz, M.H.; Juarez, A. Role of plasmid- and chromosomally encoded Hha proteins in modulation of gene expression in E. coli O157:H7. Plasmid 2013, 70, 52–60. [Google Scholar] [CrossRef]
- Porcheron, G.; Garenaux, A.; Proulx, J.; Sabri, M.; Dozois, C.M. Iron, copper, zinc, and manganese transport and regulation in pathogenic Enterobacteria: Correlations between strains, site of infection and the relative importance of the different metal transport systems for virulence. Front. Cell Infect. Microbiol. 2013, 3, 90. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.I.; Iqbal, Z.; Seleem, M.N.; Huang, D.; Sattar, A.; Hao, H.; Yuan, Z. Virulence and transcriptome profile of multidrug-resistant Escherichia coli from chicken. Sci. Rep. 2017, 7, 8335. [Google Scholar] [CrossRef] [Green Version]
- Rihtar, E.; Zgur Bertok, D.; Podlesek, Z. The uropathogenic specific protein gene usp from Escherichia coli and Salmonella bongori is a novel member of the TyrR and H-NS regulons. Microorganisms 2020, 8, 330. [Google Scholar] [CrossRef] [Green Version]
- Demirci, M. Unlu, O. and Tosun, A.I. Detection of O25b-ST131 clone, CTX-M-1 and CTX-M-15 genes via real-time PCR in Escherichia coli strains in patients with UTIs obtained from a university hospital in Istanbul. J. Infect Public Health 2019, 12, 640–644. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Supplement M100–S30; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Louvois, J. Factors influencing the assay of antimicrobial drugs in clinical samples by the agar plate diffusion method. J. Antimicrob. Chemother. 1982, 9, 253–256. [Google Scholar] [CrossRef]
- Iqbal, Z.; Caccamo, M.; Turner, I.; Flicek, P.; McVean, G. De novo assembly and genotyping of variants using colored de Bruijn graphs. Nat. Genet. 2012, 44, 226–232. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics/Disc Code | Disc Content, µg/Sensitivity Pattern | MIC, µg/mL/ Sensitivity Pattern |
|---|---|---|
| Amoxicillin-clavulanate/AMC | 30/R | 64/R |
| Cefepime/FEB | 30/R | 1024/R |
| Cefotaxime/CTX | 30/R | 1024/R |
| Ceftazidime/CAZ | 30/R | 1024/R |
| Ceftriaxone/CRO | 30/R | ≥4096/R |
| Colistin/CT | 25/I | 4/R |
| Ciprofloxacin/CIP | 5/R | 256/R |
| Doxycycline/DO | 30/R | 16/R |
| Imipenem/IPM | 10/S | 1/S |
| Gentamicin/CN | 10/I | 1/R |
| Levofloxacin/LEV | 5/R | 16/R |
| Meropenem/MEM | 10/S | 0.25/S |
| Piperacillin-tazobactam/TZP | 110/R | 64/R |
| Sulfamethoxazole-trimethoprim/SXT | 25/R | 16/R |
| Serotype a | Phylogroup b | Pathotype c | ST d | Clonotype e | Virulence Profile f | Resistance Profile g | ||
|---|---|---|---|---|---|---|---|---|
| Virulence Factor | Virulence Gene | Antimicrobial Class | Antimicrobial Resistance Genes | |||||
| O25:H4 | Group B2 | UPEC | 131 | CH40-30 | Heme binding outer membrane | chuA | β-lactams | blaOXA-1, blaCTX-M-15 |
| D-mannose-specific adhesin, type 1 fimbriae | fimH | Folate pathway antagonist | sul1, dfrA17 | |||||
| Glutamate decarboxylase | gad | Tetracyclines | tet(A) | |||||
| Heat-resistant agglutinin | hra | Fluoroquinolones | aac(6’)-Ib-cr | |||||
| Adhesin siderophore receptor | iha | Aminoglycosides | aadA5, aac(3) | |||||
| Ferric aerobactin synthetase | iucC | Macrolides | mph(A) | |||||
| Outer membrane protein (protease) T | ompT | |||||||
| Pilin subunit F43. P Fimbriae usher | papA_F43 | |||||||
| P fimbriae operon. Pilus assembly | papC | |||||||
| Secreted autotransporter toxin | sat | |||||||
| S-fimbrial adhesin | sfa | |||||||
| Tellurite resistance protein | terC | |||||||
| Outer membrane lipoprotein | traT | |||||||
| Putative chaperone-usher fimbria | yfcV | |||||||
| PAIs (GenBank Accession No.) | Size (bp) | G+C Content (%) | Position in Chromosome | No. of ORFs | Main Virulence Determinants Encoded within the PAI | Ref. | |
|---|---|---|---|---|---|---|---|
| Gene/Protein (Virulence Factor) | Function or Homology | ||||||
| PAI-EC14142-leuX (OM100123) | 101,468 | 46.3 | 4646–4,896,778 | 99 | cnf1 (cytotoxic necrotizing factor 1) | Engaged in cell necrosis | [26] |
| hylCABD (α-hemolysin) | Promotes passage of bacteria into the blood | [3] | |||||
| PAI-EC14142-asnT (OM100124) | 30,802 | 58.2 | 2,211,273–2,242,074 | 25 | fyuA (ferric yersiniabactin uptake A) | Acts as an outer-membrane receptor for iron uptake | [25] |
| irp2 (iron-repressible gene) | Involved in the biosynthesis of yersiniabactin siderophore | [2,3] | |||||
| ybtA, ybtE, ybtQ, ybtS, ybtU (yersiniabactin iron-capture island) | Involved in biosynthesis, regulation, and transfer of yersiniabactin siderophore | [2,3] | |||||
| NRPS/PKS (non-ribosomal peptide synthetase/polyketide synthase system) | Causes the breakage of DNA and cell cycle arrest in human cells | [9] | |||||
| PAI-EC14142-pheV (OM100125) | 77,503 | 47.9 | 3,357,439–3,434,941 | 79 | kpsE, kpsF, kpsU (capsule polysaccharide export inner-membrane system) | Promotes polysaccharide translocation across the inner membrane to the cell surface | [33] |
| kpsMII-K5 (K5 variant of group II capsule) | Involved in capsular polysaccharide production | [33] | |||||
| cpsF (capsular polysaccharide biosynthesis gene) | Prevents the activation of host phagocytic activity | [32] | |||||
| gspL, gspK (general secretion pathway) | Exports proteins from the bacterial cytoplasm | [34] | |||||
| hha (high hemolytic activity modulator) | Modulates expression of bacterial virulence factors | ||||||
| PAI-EC14142-icd (OM100126) | 46,444 | 51.1 | 1,340,973–1,387,416 | 61 | iss (increased serum survival) | Involved in serum resistance and evasion from phagocytosis | [28] |
| PtpT, PtpU, PtpL (phage tail proteins) | Increase resistance and virulence | [37] | |||||
| SitDCBA (iron/manganese transport system) | Involved in the transportation of Fe2+ and Mn2+ ions | [36] | |||||
| PAI-EC14142-usp (OM100127) | 7282 | 44 | 228,116–235,397 | 10 | usp (uropathogenic-specific gene) | Associated with pyelonephritis and bacteremia in an infected host | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, A.S.; Edward, E.A.; Mohamed, N.M. Pathogenicity Islands in Uropathogenic Escherichia coli Clinical Isolate of the Globally Disseminated O25:H4-ST131 Pandemic Clonal Lineage: First Report from Egypt. Antibiotics 2022, 11, 1620. https://doi.org/10.3390/antibiotics11111620
Zakaria AS, Edward EA, Mohamed NM. Pathogenicity Islands in Uropathogenic Escherichia coli Clinical Isolate of the Globally Disseminated O25:H4-ST131 Pandemic Clonal Lineage: First Report from Egypt. Antibiotics. 2022; 11(11):1620. https://doi.org/10.3390/antibiotics11111620
Chicago/Turabian StyleZakaria, Azza S., Eva A. Edward, and Nelly M. Mohamed. 2022. "Pathogenicity Islands in Uropathogenic Escherichia coli Clinical Isolate of the Globally Disseminated O25:H4-ST131 Pandemic Clonal Lineage: First Report from Egypt" Antibiotics 11, no. 11: 1620. https://doi.org/10.3390/antibiotics11111620