Can Immunization of Hens Provide Oral-Based Therapeutics against COVID-19?

 , , ,
, , , 
Abstract
:1. Introduction
2. Antibody Therapy
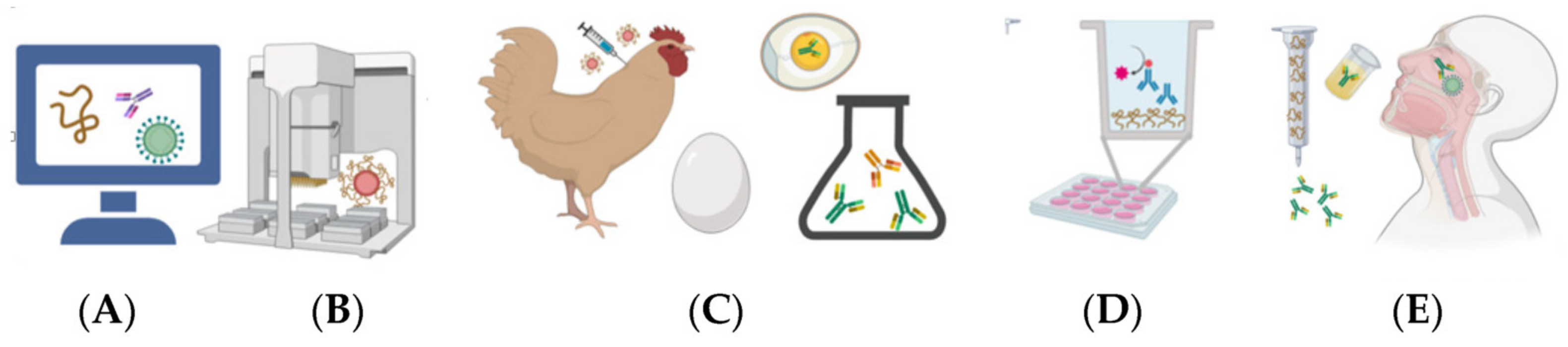
3. IgY Technology
4. Molecular Properties of IgY
5. Methods for Purification of IgY Antibodies
6. Advantages and Disadvantages of Avian IgY over Mammalian IgG
7. The Production Capacity of IgY Antibodies
8. Stability of IgY
9. Virus Protection Induced by IgY Antibodies
10. Peptide-Based Immunization against SARS-CoV-2
11. IgY as Promising and Safe Oral Treatment
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shereen, M.A.; Khan, S.; Kazmi, A.; Bashir, N.; Siddique, R. COVID-19 infection: Origin, transmission, and characteristics of human coronaviruses. J. Adv. Res. 2020, 24, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, S.-M.; Yu, X.-H.; Tang, S.-L.; Tang, C.-K. Coronavirus disease 2019 (COVID-19): Current status and future perspectives. Int. J. Antimicrob. Agents 2020, 55, 105951. [Google Scholar] [CrossRef] [PubMed]
- Gengler, I.; Wang, J.C.; Speth, M.M.; Sedaghat, A.R.; Sedaghat, A.R. Sinonasal pathophysiology of SARS-CoV -2 and COVID -19: A systematic review of the current evidence. Laryngoscope 2020, 5, 354–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitlin, L.; Cone, R.A.; Moench, T.R.; Whaley, K.J. Preventing infectious disease with passive immunization. Microbes Infect. 2000, 2, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.; Baum, A.; Pascal, K.E.; Russo, V.; Giordano, S.; Wloga, E.; Fulton, B.O.; Yan, Y.; Koon, K.; Patel, K.; et al. Studies in humanized mice and convalescent humans yield a SARS-CoV-2 antibody cocktail. Science 2020, 369, eabd0827. [Google Scholar] [CrossRef]
- Konwarh, R. Nanobodies: Prospects of Expanding the Gamut of Neutralizing Antibodies Against the Novel Coronavirus, SARS-CoV-2. Front. Immunol. 2020, 11, 1531. [Google Scholar] [CrossRef]
- Leo, O.; Cunningham, A.L.; Stern, P. Vaccine immunology. Perspect. Vaccinol. 2011, 1, 25–59. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.; Gray, D. Immunological Memory and Protective Immunity: Understanding Their Relation. Science 1996, 272, 54–60. [Google Scholar] [CrossRef]
- Dixit, R.; Herz, J.; Dalton, R.; Booy, R. Benefits of using heterologous polyclonal antibodies and potential applications to new and undertreated infectious pathogens. Vaccine 2016, 34, 1152–1161. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.P.; Goyffon, M. Antivenom serotherapy: Its applications, its limitations, its future. Bull. Soc. Pathol. Exot. 1991, 84, 286. [Google Scholar]
- Elbakri, A.; Nelson, P.N.; Abuodeh, R. The state of antibody therapy. Hum. Immunol. 2010, 71, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A. Antibody-based therapies for emerging infectious diseases. Emerg. Infect. Dis. 1996, 2, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Zolla-Pazner, S.; Pinter, A.; Mizuma, H. Potential use of serotherapy in the prevention and treatment of infection with the human immunodeficiency virus. J. Virol. Methods 1987, 17, 45–53. [Google Scholar] [CrossRef]
- Sewell, H.; Walker, F.; Sewell, F.W.H. Serotherapy for AIDS and pre-AIDS syndrome. Nature 1986, 320, 113. [Google Scholar] [CrossRef] [PubMed]
- Achard, C. Serotherapy in Tetanus. J. Nerv. Ment. Dis. 1921, 54, 340. [Google Scholar] [CrossRef]
- Klemperer, F. Ueber Natürliche Immunität und Ihre Verwerthung für die Immunisirungstherapie. Arch. Exp. Pathol. Pharmakol. 1893, 31, 356–382. [Google Scholar] [CrossRef]
- Hädge, D.; Ambrosius, H. Evolution of low molecular weight immunoglobulins—IV IgY-like immunoglobulins of birds, reptiles and amphibians, precursors of mammalian IgA. Mol. Immunol. 1984, 21, 699–707. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, H.; Tian, Z.; Chen, S.; Schade, R. Chicken Monoclonal IgY Antibody: A Novel Antibody Development Strategy. Avian Biol. Res. 2010, 3, 97–106. [Google Scholar] [CrossRef]
- Spillner, E.; Braren, I.; Greunke, K.; Seismann, H.; Blank, S.; du Plessis, D. Avian IgY antibodies and their recombinant equivalents in research, diagnostics and therapy. Biologicals 2012, 40, 313–322. [Google Scholar] [CrossRef]
- Munhoz, L.S.; Vargas, G.D.; Fischer, G.; De Lima, M.; Esteves, P.A.; Hübner, S. Avian IgY antibodies: Characteristics and applications in immunodiagnostic. Ciênc. Rural 2014, 44, 153–160. [Google Scholar] [CrossRef]
- Criste, A.; Urcan, A.C.; Corcionivoschi, N. Avian IgY antibodies, ancestors of mammalian antibodies-production and application. Rom. Biotechnol. Lett. 2019, 25, 41–49. [Google Scholar] [CrossRef]
- Cray, C.; Villar, D. Cross-reactivity of anti-chicken IgY antibody with immunoglobulins of exotic avian species. Vet. Clin. Pathol. 2008, 37, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A.; Sjöquist, J. Chicken IgY: Utilizing the evolutionary difference. Comp. Immunol. Microbiol. Infect. Dis. 1990, 13, 199–201. [Google Scholar] [CrossRef]
- Amro, W.A.; Al-Qaisi, W.; Al-Razem, F. Production and purification of IgY antibodies from chicken egg yolk. J. Genet. Eng. Biotechnol. 2018, 16, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Sigino, H.; Nitoda, T.; Juneje, L.R. General cehmical composition of hen eggs. In Hen Eggs: Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press, Tailor & Francis Group: Boca Ratón, FL, USA, 1997; pp. 13–24. ISBN 0-8493-4005-5. [Google Scholar]
- Schade, R.; Calzado, E.G.; Sarmiento, R.; Chacana, P.A.; Porankiewicz-Asplund, J.; Terzolo, H.R. Chicken egg yolk antibodies (IgY-technology): A review of progress in production and use in research and human and veterinary medicine. ATLA Altern. Lab. Anim. 2005, 33, 129–154. [Google Scholar] [CrossRef] [PubMed]
- Akita, E.M.; Nakai, S. Comparison of four purification methods for the production of immunoglobulins from eggs laid by hens immunized with an enterotoxigenic E. coli strain. J. Immunol. Methods 1993, 160, 207–214. [Google Scholar] [CrossRef]
- Hodek, P.; Trefil, P.; Simunek, J.; Hudecek, J.; Stiborova, M. Optimized protocol of chicken antibody (IgY) purification providing electrophoretically homogenous preparations. Int. J. Electrochem. Sci. 2013, 8, 113–124. [Google Scholar]
- Mehdi, M.; Dallal, S.; Kalantar, E.; Khorasani, L.K.; Hassan, M.; Tehrani, H.; Zamani, Z.; Zamani, M.Z. Identification and extraction of chicken egg yolk immunoglobulin from egg by polyethylene glycol (PEG) precipitation. J. Med. Bacteriol. 2015, 4, 13–18. [Google Scholar]
- Pauly, D.; Chacana, P.A.; Calzado, E.G.; Brembs, B.; Schade, R. Igy technology: Extraction of chicken antibodies from egg yolk by polyethylene glycol (PEG) precipitation. J. Vis. Exp. 2011, e3084. [Google Scholar] [CrossRef]
- Polson, A.; von Wechmar, M.B.; van Regenmortel, M.H.V. Isolation of viral IgY antibodies from yolks of immunized hens. Immunol. Commun. 1980, 9, 475–493. [Google Scholar] [CrossRef]
- Gutiérrez Calzado, E.J.; Chacana, P.A.; Pauly, D.; Schade, R. IgY-technology, the immunization of laying hen and the extraction of antibodies from egg yolk by polyethylene glycol (PEG) precipitation. Rev. CENIC. Cienc. Biol. 2010, 41, 1–10. [Google Scholar]
- Tan, S.H.; Mohamedali, A.; Kapur, A.; Lukjanenko, L.; Baker, M.S. A novel, cost-effective and efficient chicken egg IgY purification procedure. J. Immunol. Methods 2012, 380, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Sesarman, A.; Mihai, S.; Chiriac, M.T.; Olaru, F.; Sitaru, A.G.; Thurman, J.M.; Zillikens, D.; Sitaru, C. Binding of avian IgY to type VII collagen does not activate complement and leucocytes and fails to induce subepidermal blistering in mice. Br. J. Dermatol. 2008, 158, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Hädge, D.; Fiebig, H.; Puskas, E.; Ambrosius, H. Evolution of low molecular weight immunoglobulins II. No antigenic cross-reactivity of human IgD, human IgG and IgG3 to chicfen IgY. Dev. Comp. Immunol. 1980, 4, 725–736. [Google Scholar] [CrossRef]
- Härtle, S.; Magor, K.E.; Göbel, T.W.; Davison, F.; Kaspers, B. Structure and evolution of avian immunoglobulins. In Avian Immunology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 103–120. ISBN 9780123969651. [Google Scholar]
- Zhang, X.; Calvert, R.A.; Sutton, B.J.; Doré, K.A. IgY: A key isotype in antibody evolution. Biol. Rev. 2017, 92, 2144–2156. [Google Scholar] [CrossRef]
- Fischer, M.; Hlinak, A.; Montag, T.; Claros, M.; Schade, R.; Ebner, D. Comparison of standard methods for purification of egg yolk antibodies. Tierarztl. Prax. Ausgabe G Grosstiere Nutztiere 1996, 24, 411–418. [Google Scholar]
- Hatta, H.; Horimoto, Y. Applications of egg yolk antibody (IgY) in diagnosis reagents and in prevention of diseases. In Eggs as Functional Foods and Nutraceuticals for Human Health; Royal Society of Chemistry: London, UK, 2019; pp. 305–328. ISBN 978-1-78801-213-3. [Google Scholar]
- Esmailnejad, A.; Hachesoo, B.A.; Nasab, E.H.; Shakoori, M. Storage stability of anti-Salmonella typhimurium immunoglobulin Y in immunized quail eggs stored at 4 °C. Indian J. Anim. Sci. 2019, 89, 1318–1321. [Google Scholar]
- Nilsson, E.; Stålberg, J.; Larsson, A. IgY stability in eggs stored at room temperature or at +4 °C. Br. Poult. Sci. 2012, 53, 42–46. [Google Scholar] [CrossRef]
- Fu, C.Y.; Huang, H.; Wang, X.M.; Liu, Y.G.; Wang, Z.G.; Cui, S.J.; Gao, H.L.; Li, Z.; Li, J.P.; Kong, X.G. Preparation and evaluation of anti-SARS coronavirus IgY from yolks of immunized SPF chickens. J. Virol. Methods 2006, 133, 112–115. [Google Scholar] [CrossRef]
- Somasundaram, R.; Choraria, A.; Antonysamy, M. An approach towards development of monoclonal IgY antibodies against SARS CoV-2 spike protein (S) using phage display method: A review. Int. Immunopharmacol. 2020, 85, 106654. [Google Scholar] [CrossRef]
- Nguyen, H.H.; Tumpey, T.M.; Park, H.-J.; Han, G.-Y.; Lee, J.; Byun, Y.-H.; Song, J.-M.; Tran, L.D.; Nguyen, V.D.; Kilgore, P.E.; et al. Avian antibodies to combat potential H5N1 pandemic and seasonal influenza. Influenza Respi. Viruses 2011, 5, 233–236. [Google Scholar]
- Yang, Y.E.; Wen, J.; Zhao, S.; Zhang, K.; Zhou, Y. Prophylaxis and therapy of pandemic H1N1 virus infection using egg yolk antibody. J. Virol. Methods 2014, 206, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Sheng, Y.; Yu, W.; Zhao, S.; Shan, H.; Zhang, Q.; Wang, Z. Comparison of chicken IgY and mammalian IgG in three immunoassays for detection of sulfamethazine in milk. Food Anal. Methods 2018, 11, 3452–3463. [Google Scholar] [CrossRef]
- Vöhringer, C.; Sander, E. Comparison of antibodies in chicken egg yolk (IgY) and rabbit (IgG) for quantitative strain detection of Colletotrichum falcatum and Fusarium subglutinans with ELISA. Z. Pflanzenkrankh. Pflanzenschutz 2001, 108, 39–48. [Google Scholar] [CrossRef]
- Sheng, Y.J.; Ni, H.J.; Zhang, H.Y.; Li, Y.H.; Wen, K.; Wang, Z.H. Production of chicken yolk IgY to sulfamethazine: Comparison with rabbit antiserum IgG. Food Agric. Immunol. 2015, 26, 305–316. [Google Scholar] [CrossRef]
- Pechelyulko, A.A.; Tarakanova, Y.N.; Dmitriev, D.A.; Massino, Y.S.; Kost, V.Y.; Rogozhin, E.A.; Segal, O.L.; Dmitriev, A.D. Comparative analysis of the efficiency of chicken and rabbit antibodies in competitive enzyme linked immunoassay for the detection of bovine beta-casomorphin 7. Appl. Biochem. Microbiol. 2019, 55, 704–710. [Google Scholar] [CrossRef]
- Krief, A.; Letesson, J.J.; Billen, D. Comparison between “IgY technology” from chickens and “IgG technology” from mice for production of tailor-made antibodies. Tetrahedron Lett. 2002, 43, 1843–1846. [Google Scholar] [CrossRef]
- Adachi, K.; Handharyani, E.; Sari, D.; Takama, K.; Fukuda, K.; Endo, I.; Yamamoto, R.; Sawa, M.; Tanaka, M.; Konishi, I.; et al. Development of neutralization antibodies against highly pathogenic H5N1 avian influenza virus using ostrich (Struthio camelus) yolk. Mol. Med. Rep. 2008, 1, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.; Larsson, A. Stability of chicken IgY antibodies freeze-dried in the presence of lactose, sucrose and threalose. J. Poult. Sci. 2007, 44, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; He, Z.; Liu, Y.; Ma, M.; Cai, Z. Mass spectrometry characterization for N-glycosylation of immunoglobulin Y from hen egg yolk. Int. J. Biol. Macromol. 2018, 108, 277–283. [Google Scholar] [CrossRef]
- Wang, B.; Yang, J.; Cao, S.; Wang, H.; Pan, X.; Zhu, J.; Zhou, Y.; Gao, L.; Li, W.; Li, M. Preparation of specific anti-Helicobacter pylori yolk antibodies and their antibacterial effects. Int. J. Clin. Exp. Pathol. 2014, 7, 6430–6437. [Google Scholar] [PubMed]
- Zajac, J.; Schubert, A.; Dyck, T.; Oelkrug, C. IgY antibodies for the prevention and treatment of Helicobacter pylori infections. J. Med. Microbiol. Diagn. 2017, 6. [Google Scholar] [CrossRef]
- Najdi, S.; Nikbakht Brujeni, G.; Sheikhi, N.; Chakhkar, S. Development of anti-Helicobacter pylori immunoglobulins Y (IgYs) in quail. Iran. J. Vet. Res. 2016, 17, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Hatta, H.; Ozeki, M.; Tsuda, K. Egg yolk antibody IgY and its application. In Hen Eggs: Basic and Applied Science; Yamamoto, T., Juneja, L.R., Hatta, H., Kim, M., Eds.; CRC Press: Taylor & Francis Group: Boca Raton, FL, USA, 1997; pp. 151–178. ISBN 0-8493-4005-5. [Google Scholar]
- Hatta, H.; Tsuda, K.; Akachi, S.; Kim, M.; Yamamoto, T. Productivity and some properties of egg yolk antibody (IgY) against human rotavirus compared with rabbit IgG. Biosci. Biotechnol. Biochem. 1993, 57, 450–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wei, Y.; Li, Y.; Wang, X.; Liu, Y.; Tian, D.; Jia, X.; Gong, R.; Liu, W.; Yang, L. IgY antibodies against Ebola virus possess post-exposure protection and excellent thermostability. bioRxiv 2020. [Google Scholar] [CrossRef]
- Shimizu, M.; Nagashima, H.; Hashimoto, K.; Suzuki, T. Egg yolk antibody (Ig Y) stability in aqueous solution with high sugar concentrations. J. Food Sci. 1994, 59, 763–765. [Google Scholar] [CrossRef]
- Guler, E.M.; Kesmen, M.; Dedeakayogullari, H.; Oskouei, Z.M.; Kilinc, A.; Suha Yalcin, A. Production of hen egg IgY liposomes against different salmonella species. FEBS J. 2013, 280, 481. [Google Scholar]
- Shimizu, M.; Miwa, Y.; Hashimoto, K.; Goto, A. Encapsulation of chicken egg yolk immunoglobulin G (IgY) by liposomes. Biosci. Biotechnol. Biochem. 1993, 57, 1445–1449. [Google Scholar] [CrossRef] [Green Version]
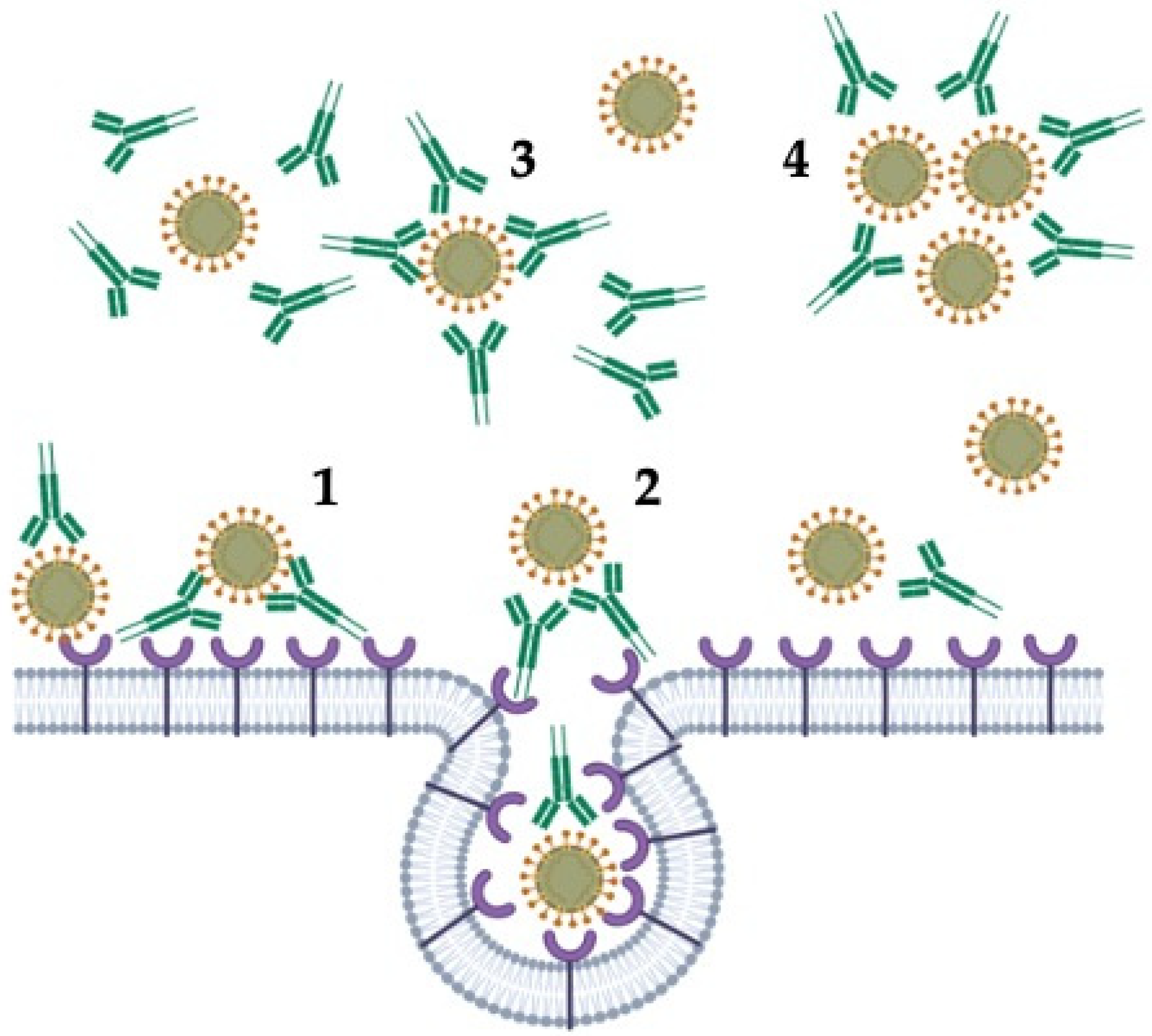
- VanBlargan, L.A.; Goo, L.; Pierson, T.C. Deconstructing the antiviral neutralizing-antibody response: Implications for vaccine development and immunity. Microbiol. Mol. Biol. Rev. 2016, 80, 989–1010. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.H.; Tumpey, T.M.; Park, H.J.; Byun, Y.H.; Tran, L.D.; Nguyen, V.D.; Kilgore, P.E.; Czerkinsky, C.; Katz, J.M.; Seong, B.L.; et al. Prophylactic and therapeutic efficacy of avian antibodies against influenza virus H5N1 and H1N1 in mice. PLoS ONE 2010, 5, e10152. [Google Scholar] [CrossRef]
- Taylor, A.; Foo, S.S.; Bruzzone, R.; Vu Dinh, L.; King, N.J.C.; Mahalingam, S. Fc receptors in antibody-dependent enhancement of viral infections. Immunol. Rev. 2015, 268, 340–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallach, M.G.; Webby, R.J.; Islam, F.; Walkden-Brown, S.; Emmoth, E.; Feinstein, R.; Gronvik, K.O. Cross-protection of chicken immunoglobulin Y antibodies against H5N1 and H1N1 viruses passively administered in mice. Clin. Vaccine Immunol. 2011, 18, 1083–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thu, H.M.; Myat, T.W.; Win, M.M.; Thant, K.Z.; Rahman, S.; Umeda, K.; Van Nguyen, S.; Icatlo, F.C.; Higo-Moriguchi, K.; Taniguchi, K.; et al. Chicken egg yolk antibodies (IgY) for prophylaxis and treatment of rotavirus diarrhea in human and animal neonates: A concise review. Korean J. Food Sci. Anim. Resour. 2017, 37, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fink, A.L.; Williams, K.L.; Harris, E.; Alvine, T.D.; Henderson, T.; Schiltz, J.; Nilles, M.L.; Bradley, D.S. Dengue virus specific IgY provides protection following lethal dengue virus challenge and is neutralizing in the absence of inducing antibody dependent enhancement. PLoS Negl. Trop. Dis. 2017, 11, e0005721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, K.L.; Fink, A.; Nilles, M.L.; Bradley, D.S. Dengue NS1-specific IgY antibodies neutralizes dengue infection without inducing antibody dependent enhancement. J. Immunol. 2017, 198 (Suppl. 1), 225.3. [Google Scholar]
- O’Donnell, K.L.; Espinosa, D.; Nilles, M.L.; Harris, E.; Bradley, D.S. Avian IgY antibodies recognize novel dengue NS1 epitopes with the ability to neutralize infection and reduce vascular leakage without inducing antibody dependent enhancement. Am. J. Trop. Med. Hyg. 2018, 99, 512. [Google Scholar]
- O’Donnell, K.L.; Meberg, B.; Schiltz, J.; Nilles, M.L.; Bradley, D.S. Zika virus-specific IgY results are therapeutic following a lethal zika virus challenge without inducing antibody-dependent enhancement. Viruses 2019, 11, 301. [Google Scholar] [CrossRef] [Green Version]
- Brocato, R.; Josleyn, M.; Ballantyne, J.; Vial, P.; Hooper, J.W. DNA vaccine-generated duck polyclonal antibodies as a postexposure prophylactic to prevent Hantavirus Pulmonary Syndrome (HPS). PLoS ONE 2012, 7, e35996. [Google Scholar] [CrossRef]
- Haese, N.; Brocato, R.L.; Henderson, T.; Nilles, M.L.; Kwilas, S.A.; Josleyn, M.D.; Hammerbeck, C.D.; Schiltz, J.; Royals, M.; Ballantyne, J.; et al. Antiviral biologic produced in DNA vaccine/goose platform protects hamsters against hantavirus pulmonary syndrome when administered post-exposure. PLoS Negl. Trop. Dis. 2015, 9, e0003803. [Google Scholar] [CrossRef] [Green Version]
- Motoi, Y.; Sato, K.; Hatta, H.; Morimoto, K.; Inoue, S.; Yamada, A. Production of rabies neutralizing antibody in hen’s eggs using a part of the G protein expressed in Escherichia coli. Vaccine 2005, 23, 3026–3032. [Google Scholar] [CrossRef]
- Conroy, N.; Vlack, S.; Williams, J.M.; Patten, J.J.; Horvath, R.L.; Lambert, S.B. Using serology to assist with complicated post-exposure prophylaxis for rabies and australian bat lyssavirus. PLoS Negl. Trop. Dis. 2013, 7, e2066. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Park, C.H.; Hatta, H. Production of polyclonal rabies virus antibodies in birds. In Current Laboratory Techniques in Rabies Diagnosis, Research and Prevention; Academic Press: Cambridge, MA, USA, 2015; Volume 2, pp. 305–316. [Google Scholar]
- Gomez, L.A.; Totapalli, V.; Diraviyam, T.; Michael, A.; Vani, C. Isolation, characterization, comparative analysis of anti-rabies antibodies raised in chicken and rabbit. J. Pure Appl. Microbiol. 2017, 11, 323–327. [Google Scholar] [CrossRef]
- Motoi, Y.; Inoue, S.; Hatta, H.; Sato, K.; Morimoto, K.; Yamada, A. Detection of rabies-specific antigens by egg yolk antibody (IgY) to the recombinant rabies virus proteins produced in Escherichia coli. Jpn. J. Infect. Dis. 2005, 58, 115–118. [Google Scholar] [PubMed]
- Dai, Y.C.; Wang, Y.Y.; Zhang, X.F.; Tan, M.; Xia, M.; Wu, X.B.; Jiang, X.; Nie, J. Evaluation of anti-norovirus IgY from egg yolk of chickens immunized with norovirus P particles. J. Virol. Methods 2012, 186, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.C.; Zhang, X.F.; Tan, M.; Huang, P.; Lei, W.; Fang, H.; Zhong, W.; Jiang, X. A dual chicken IgY against rotavirus and norovirus. Antiviral Res. 2013, 97, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Ma, Y.; Lu, M.; Zhang, Y.; Li, A.; Liang, X.; Li, J. Efficient production of human norovirus-specific IgY in egg yolks by vaccination of hens with a recombinant vesicular stomatitis virus expressing VP1 protein. Viruses 2019, 11, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivalingam, G.N.; Shepherd, A.J. An analysis of B-cell epitope discontinuity. Mol. Immunol. 2012, 51, 304–309. [Google Scholar] [CrossRef]
- Khan, A.; Alam, A.; Imam, N.; Siddiqui, M.F.; Ishrat, R. Design of an epitope-based peptide vaccine against the Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2): A vaccine informatics approach. bioRxiv 2020. [Google Scholar] [CrossRef]
- Trier, N.H.; Hansen, P.R.; Houen, G. Production and characterization of peptide antibodies. Methods 2012, 56, 136–144. [Google Scholar] [CrossRef]
- Lee, B.S.; Huang, J.S.; Jayathilaka, L.P.; Lee, J.; Gupta, S. Antibody production with synthetic peptides. In High-Resolution Imaging of Cellular Proteins; Humana Press Inc.: New York, NY, USA, 2016; Volume 1474, pp. 25–47. [Google Scholar]
- Grant, G.A. Synthetic peptides for production of antibodies that recognize intact proteins. Curr. Protoc. Mol. Biol. 2002, 59, 11–16. [Google Scholar] [CrossRef]
- Hansen, P.R. Peptide-carrier conjugation. In Peptide Antibodies; Humana Press Inc.: New York, NY, USA, 2015; Volume 1348, pp. 51–57. [Google Scholar]
- Lee, B.S.; Huang, J.S.; Jayathilaka, G.D.L.P.; Lateef, S.S.; Gupta, S. Production of antipeptide antibodies. In Immunoelectron Microscopy; Humana Press Inc.: Totowa, NJ, USA, 2010; Volume 657, pp. 93–108. ISBN 9781607617822. [Google Scholar]
- Zhou, Z.; Zhou, R.; Tang, Z. Effects of topical application of immunoglobulin yolk on mutans streptococci in dental plaque. West China J. Stomatol. 2003, 21, 295–297. [Google Scholar]
- Takeuchi, S.; Motohashi, J.; Kimori, H.; Nakagawa, Y.; Tsurumoto, A. Effects of oral moisturising gel containing egg yolk antibodies against Candida albicans in older people. Gerodontology 2016, 33, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, K.; Sugano, N.; Shimada, T.; Shofiqur, R.A.K.M.; Ibrahim, E.S.M.; Isoda, R.; Umeda, K.; Van Sa, N.; Kodama, Y.; Ito, K. Effects of egg yolk antibody against Porphyromonas gingivalis gingipains in periodontitis patients. J. Oral Sci. 2007, 49, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizarro-Guajardo, M.; Díaz-González, F.; Álvarez-Lobos, M.; Paredes-Sabja, D. Characterization of chicken IgY specific to Clostridium difficile R20291 spores and the effect of oral administration in mouse models of initiation and recurrent disease. Front. Cell. Infect. Microbiol. 2017, 7, 365. [Google Scholar] [CrossRef]
- Larsson, A.; Carlander, D. Oral immunotherapy with yolk antibodies to prevent infections in humans and animals. Upsala J. Med. Sci. 2003, 108, 129–140. [Google Scholar]
- Wilhelmson, M.; Carlander, D.; Kreuger, A.; Kollberg, H.; Larsson, A. Oral treatment with yolk antibodies for the prevention of C. albicans infections in chemotherapy treated children. A feasibility study. Food Agric. Immunol. 2005, 16, 41–45. [Google Scholar] [CrossRef]
- Gandhimathi, C.; Michael, A. Efficacy of oral passive immunotherapy against dental caries in humans using chicken egg yolk antibodies generated against streptococcus mutans. Int. J. Pharma Bio Sci. 2015, 6, B652–B663. [Google Scholar]
- Karlsson, M.; Kollberg, H.; Larsson, A. Chicken IgY: Utilizing the evolutionary advantage. World’s Poult. Sci. J. 2004, 60, 341–348. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Mine, Y. Using egg IgY antibodies for health, diagnostic and other industrial applications. In Improving the Safety and Quality of Eggs and Egg Products; Woodhead Publishing: Sawston, UK; Cambridge, UK, 2011; Volume 2, pp. 346–373. ISBN 9780857090720. [Google Scholar]
- Kovacs-Nolan, J.; Mine, Y. Avian egg antibodies: Basic and potential applications. Avian Poult. Biol. Rev. 2004, 15, 25–46. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, E.; Amini, A.; Wretlind, B.; Larsson, A. Pseudomonas aeruginosa infections are prevented in cystic fibrosis patients by avian antibodies binding Pseudomonas aeruginosa flagellin. J. Chromatogr. B 2007, 856, 75–80. [Google Scholar] [CrossRef]
- Nilsson, E.; Larsson, A.; Olesen, H.V.; Wejåker, P.E.; Kollberg, H. Good effect of IgY against Pseudomonas aeruginosa infections in cystic fibrosis patients. Pediatr. Pulmonol. 2008, 43, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Schuster, A.; Bend, J.; Høiby, N.; Verde, P.E.; Rottmann, A.; Larsson, A. WS12-5 Clinical study to evaluate an anti-Pseudomonas aeruginosa IgY gargling solution (EUDRACT 2011-000801-39). J. Cyst. Fibros. 2019, 18, S23. [Google Scholar] [CrossRef]
- Sudjarwo, S.A.; Indriyani, W.; Nasronudin; Sudjarwo, G.W. Production and characterization protein of anti HIV specific immunoglobulin Y for immunotherapy. J. Appl. Pharm. Sci. 2014, 4, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Horie, K.; Horie, N.; Abdou, A.M.; Yang, J.O.; Yun, S.S.; Chun, H.N.; Park, K.; Kim, M.; Hatta, H. Suppressive effect of functional drinking yogurt containing specific egg yolk immunoglobulin on Helicobacter pylori in humans. J. Dairy Sci. 2004, 87, 4073–4079. [Google Scholar] [CrossRef]
- Leiva, C.L.; Gallardo, M.J.; Casanova, N.; Terzolo, H.; Chacana, P. IgY-technology (egg yolk antibodies) in human medicine: A review of patents and clinical trials. Int. Immunopharmacol. 2020, 81, 106269. [Google Scholar] [CrossRef] [PubMed]
- Ku, Z.; Ye, X.; Salazar, G.T.; Zhang, N.; An, Z. Antibody therapies for the treatment of COVID-19. Antib. Ther. 2020, 3, 101–108. [Google Scholar] [CrossRef]
- Shetty, R.; Ghosh, A.; Honavar, S.G.; Khamar, P.; Sethu, S. Therapeutic opportunities to manage COVID-19/SARS-CoV-2 infection: Present and future. Indian J. Ophthalmol. 2020, 68, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.C.; Pande, V.; Sati, D.; Upreti, S.; Samant, M. Vaccination strategies to combat novel corona virus SARS-CoV-2. Life Sci. 2020, 256, 117956. [Google Scholar] [CrossRef]
- Amanat, F.; Krammer, F. SARS-CoV-2 vaccines: Status report. Immunity 2020, 52, 583–589. [Google Scholar] [CrossRef]
- Constantin, C.; Neagu, M.; Supeanu, T.; Chiurciu, V.; Spandidos, D. IgY-turning the page toward passive immunization in COVID-19 infection (Review). Exp. Ther. Med. 2020, 20, 151–158. [Google Scholar] [CrossRef]
- Albehwar, A.M.; Tammam, A.A.; Hassan, A.I. Efficacy of Antirabies IgG and IgY on Protection of Mice against Experimental Viral Infection as a Model for Emergency Intervention. Glob. J. Med. Res. 2016, 16. [Google Scholar]
- Shanmugaraj, B.; Siriwattananon, K.; Wangkanont, K.; Phoolcharoen, W. Perspectives on monoclonal antibody therapy as potential therapeutic intervention for Coronavirus disease-19 (COVID-19). Asian Pac. J. Allergy Immunol. 2020, 38, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Madani, F.; Hsein, H.; Busignies, V.; Tchoreloff, P. An overview on dosage forms and formulation strategies for vaccines and antibodies oral delivery. Pharm. Dev. Technol. 2020, 25, 133–148. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
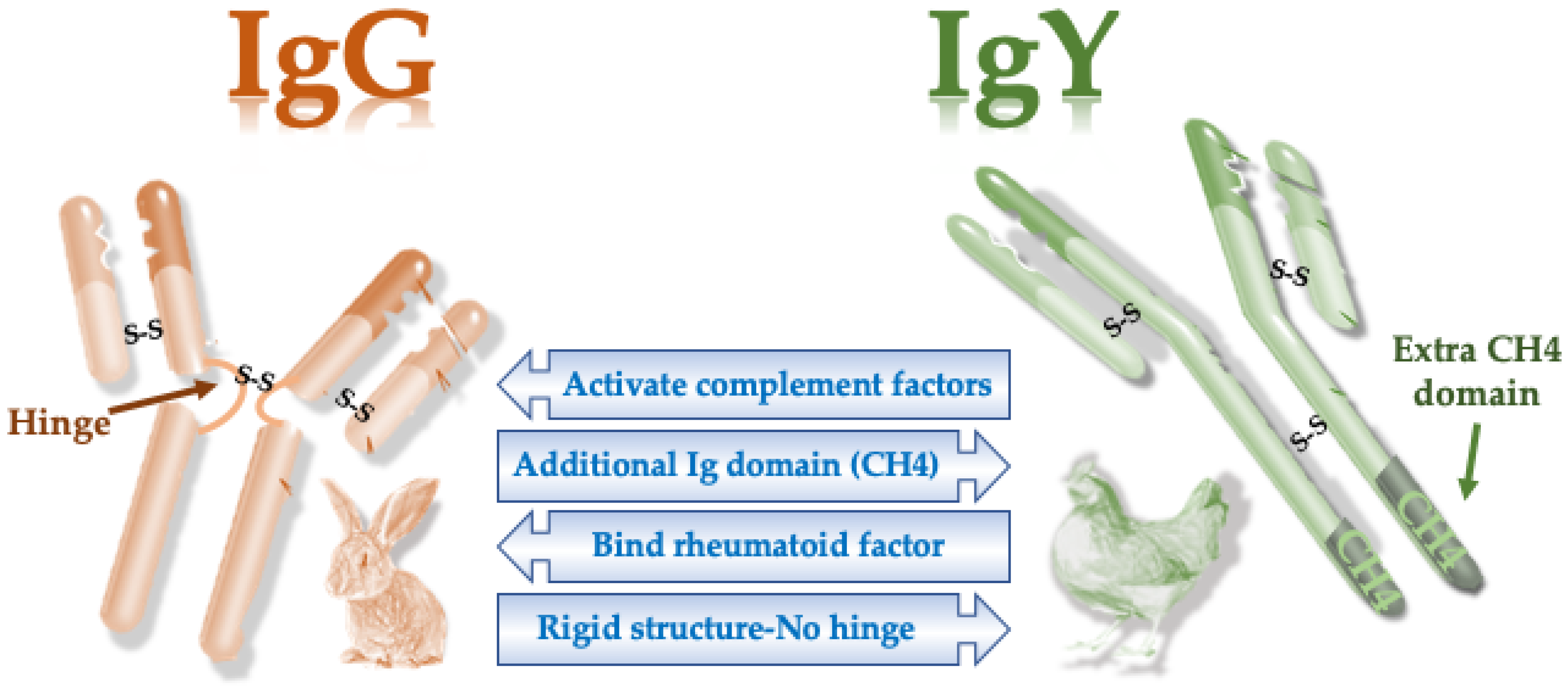
| Molecular Feature | Mammalian IgG | Avian IgY |
|---|---|---|
| Molecular weight | 150 kDa | 180 kDa |
| No. of constant domains | 4 | 3 |
| Isoelectric point | 6.1–8.5 | 5.7–7.6 |
| Hinge region | Present | Absent |
| Complement binding | Present | Absent |
| Rheumatoid factor binding | Present | Absent |
| Mediates anaphylaxis | No | Yes |
| Binding to protein A or G | Yes | No |
| Advantages | Disadvantages |
|---|---|
| Easy to produce and easily sampled | Mixed with other non-specific antibodies |
| Low cost, less invasive, a higher amount | Less used in the pharmaceutical industry |
| Can identify various epitopes on a given antigen | Cannot be purified by protein A or G |
| Detect small changes within antigens | Risk of different efficacy among batches |
| Remain stable at variable pH and temperatures | - |
| Can be produced in large batches | - |
| Can be safely used in the food industry | - |
| Perform better recognition of conserved mammalian epitopes than IgGs | - |
| No cross-reactivity with rheumatoid factors | - |
| No activation of the mammalian complement | - |
| High resistance to immunization with toxins | - |
| VIRUS | Model | Antigen | Effect | Reference |
|---|---|---|---|---|
| INFLUENZA A | Mice | whole inactivated viruses | In vivo protection against lethal challenge with H5N1 | [44,64,66] |
| ROTAVIRUS | Young children | Whole virion particle | Reduced the severity of clinical manifestation of diarrhea among IgY-treated subjects | [67] |
| DENGUE | Mice | Dengue Type 2 Antigen | Neutralizing but not enhancing virus infection | [68,69,70] |
| ZIKA | Mice | Inactivated PRVABC59 Zika virus | In vitro inhibition without inducing ADE | [71] |
| HANTAVIRUS | Hamster | M segment-DNA vaccine | In vivo efficacy after intranasal challenge | [72,73] |
| RABIES | Mice | Recombinant rG-F2 protein | Neutralized rabies virus infectivity | [74,75,76,77,78] |
| SARS | In vitro binding assay to Vero E6 cells | Inactivated SARS virus strain BJ01 | In vitro neutralization | [42] |
| NOROVIRUS | In vitro binding and blocking assay | Recombinant P particles | In vitro blocking activity | [79,80] |
| EBOLA | Mice | Recombinant antigens | In vivo protection | [59] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez de la Lastra, J.M.; Baca-González, V.; Asensio-Calavia, P.; González-Acosta, S.; Morales-delaNuez, A. Can Immunization of Hens Provide Oral-Based Therapeutics against COVID-19? Vaccines 2020, 8, 486. https://doi.org/10.3390/vaccines8030486
Pérez de la Lastra JM, Baca-González V, Asensio-Calavia P, González-Acosta S, Morales-delaNuez A. Can Immunization of Hens Provide Oral-Based Therapeutics against COVID-19? Vaccines. 2020; 8(3):486. https://doi.org/10.3390/vaccines8030486
Chicago/Turabian StylePérez de la Lastra, José M., Victoria Baca-González, Patricia Asensio-Calavia, Sergio González-Acosta, and Antonio Morales-delaNuez. 2020. "Can Immunization of Hens Provide Oral-Based Therapeutics against COVID-19?" Vaccines 8, no. 3: 486. https://doi.org/10.3390/vaccines8030486