
Acute HIV-1 and SARS-CoV-2 Infections Share Slan+ Monocyte Depletion—Evidence from an Hyperacute HIV-1 Case Report
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Controls
2.2. Flow Cytometry
2.3. Unsupervised Analysis
2.4. HIV-1 Read-Outs
2.5. SARS-CoV2 Plasma Viral Load
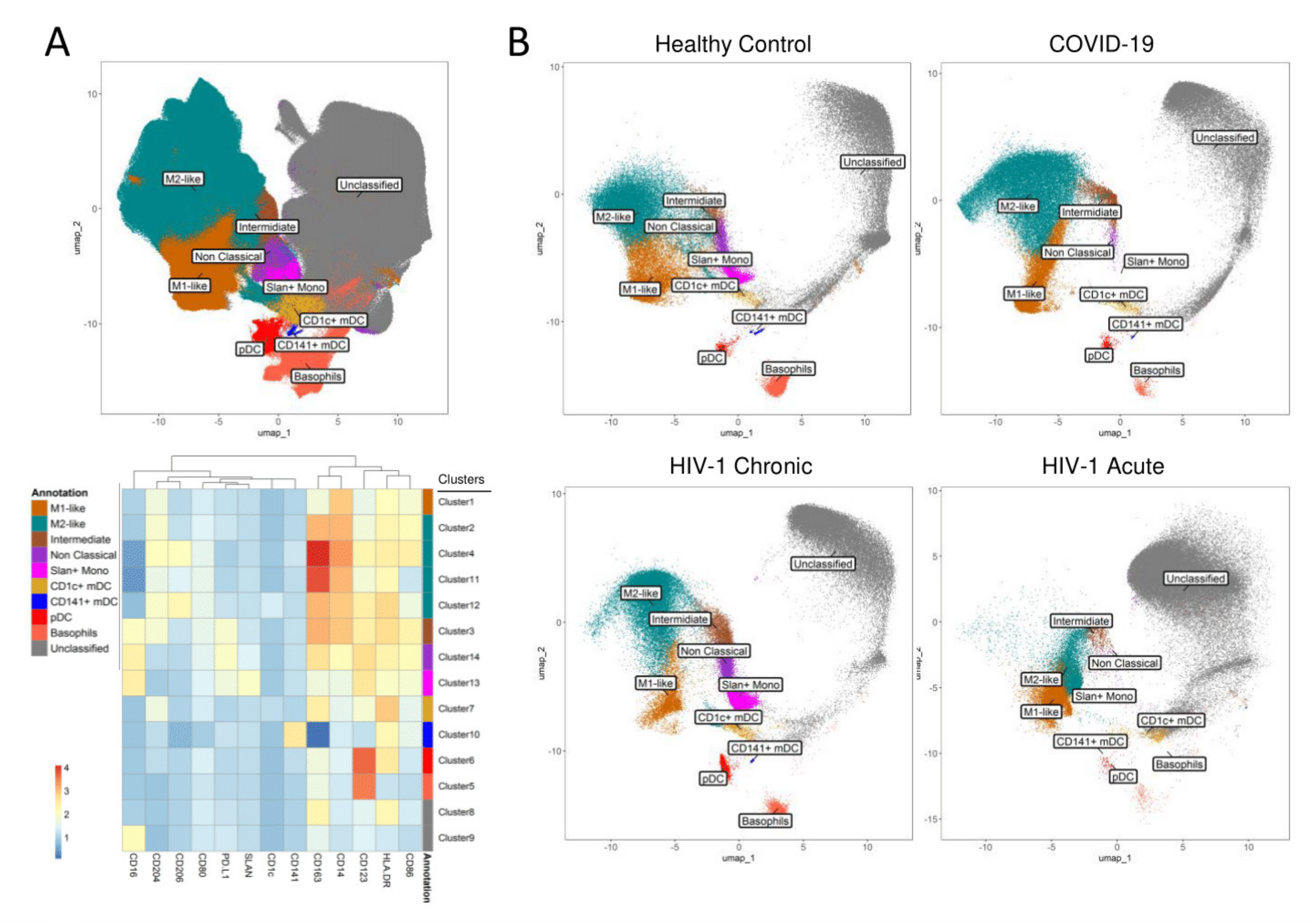
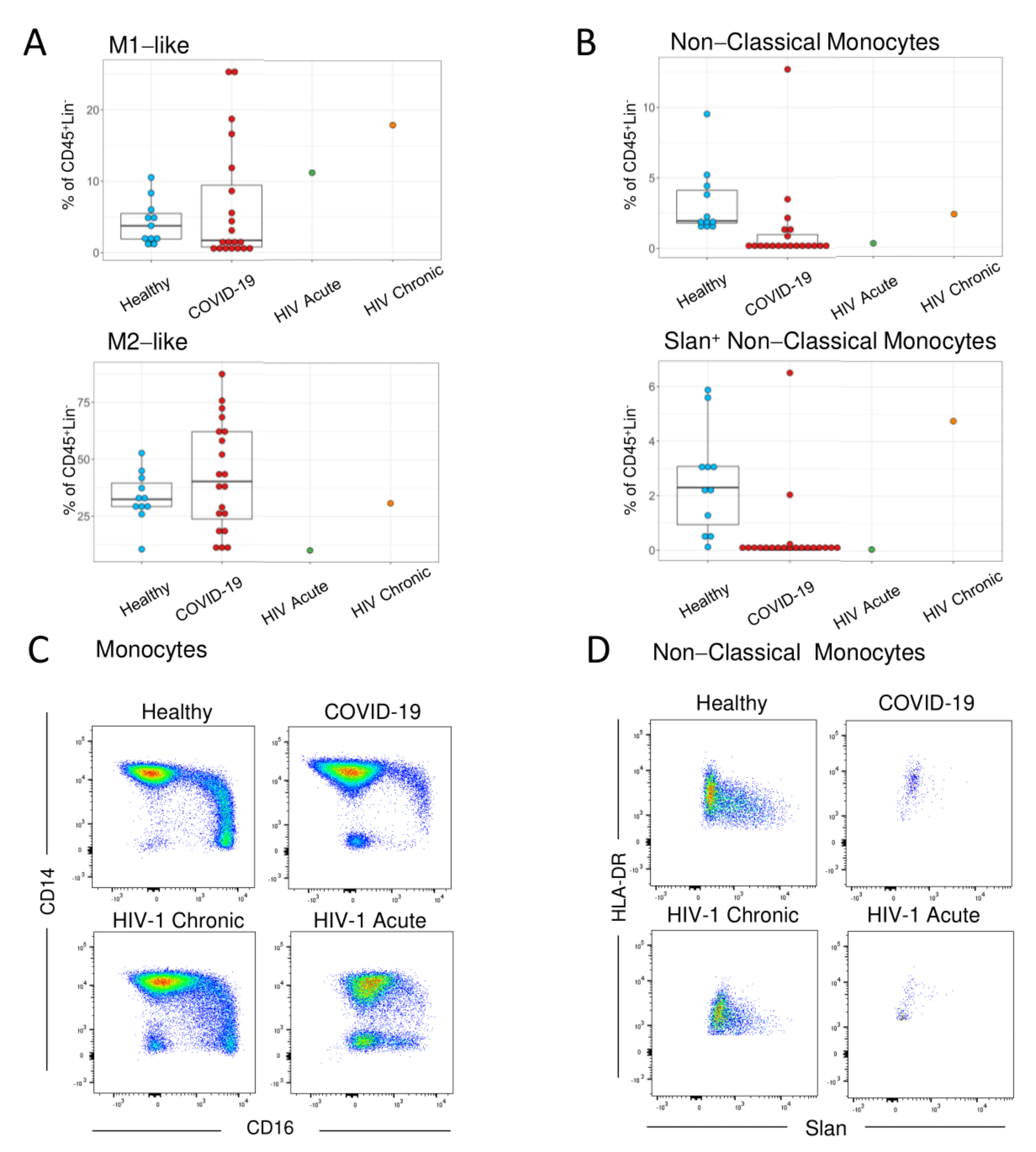
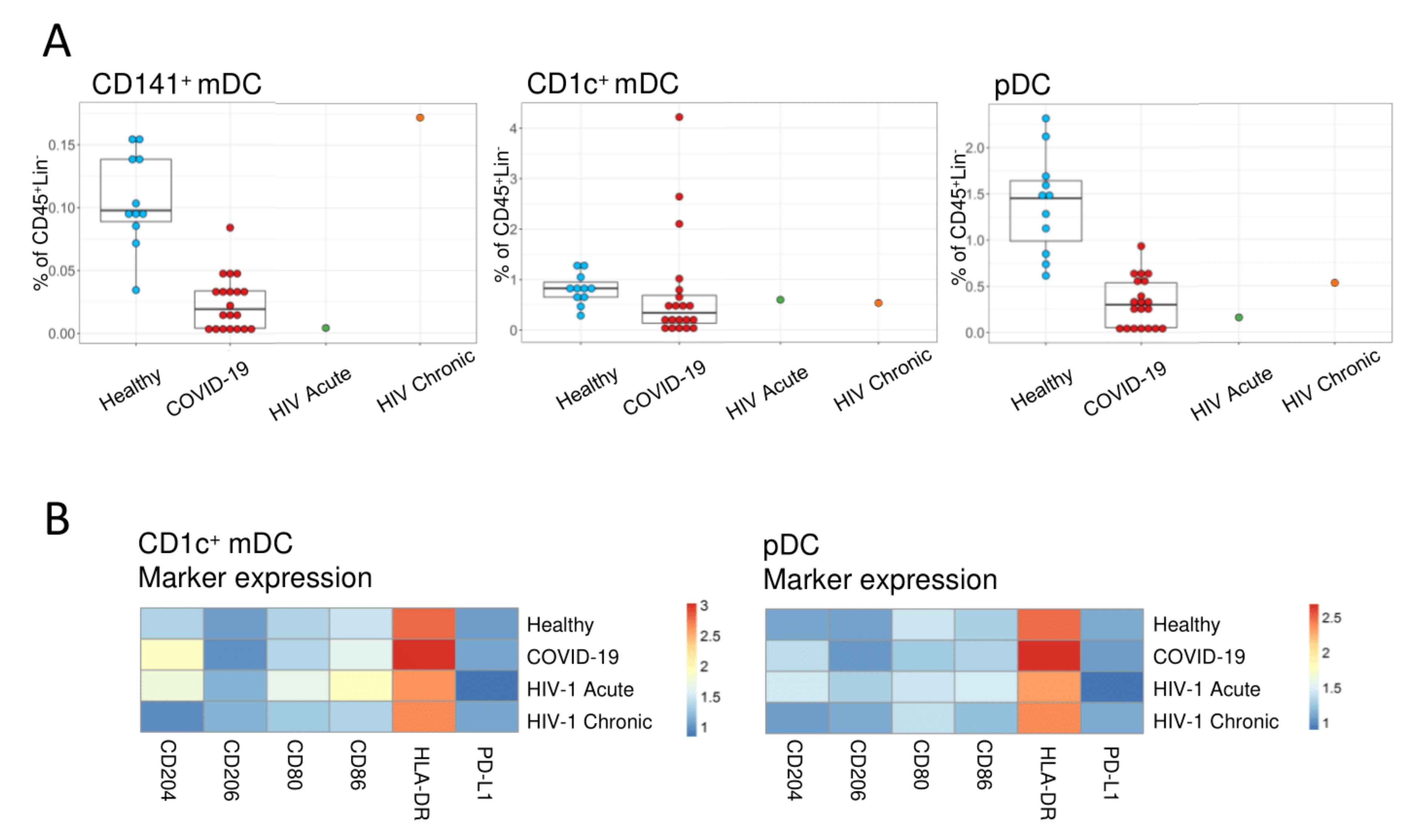
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jain, V.; Hartogensis, W.; Bacchetti, P.; Hunt, P.W.; Hatano, H.; Sinclair, E.; Epling, L.; Lee, T.H.; Busch, M.P.; McCune, J.M.; et al. Antiretroviral therapy initiated within 6 months of HIV infection is associated with lower T-cell activation and smaller HIV reservoir size. J. Infect. Dis. 2013, 208, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilliams, M.; Mildner, A.; Yona, S. Developmental and Functional Heterogeneity of Monocytes. Immunity 2018, 49, 595–613. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Naveed, M.; Naeem, M.; Ur Rahman, M.; Gul Hilal, M.; Kakakhel, M.A.; Ali, G.; Hassan, A. Review of potential risk groups for coronavirus disease 2019 (COVID-19). New Microbes New Infect. 2021, 41. [Google Scholar] [CrossRef]
- Costa, P.; Rusconi, S.; Mavilio, D.; Fogli, M.; Murdaca, G.; Pende, D.; Mingari, M.C.; Galli, M.; Moretta, L.; De Maria, A. Differential disappearance of inhibitory natural killer cell receptors during HAART and possible impairment of HIV-1-specific CD8 cytotoxic T lymphocytes. AIDS 2001, 15, 965–974. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, Q.; Chen, P.; Guo, N.; Song, A.; Huang, X.; Xia, W.; Li, L.; Moog, C.; Wu, H.; et al. Foxp3+Helios+ regulatory T cells are associated with monocyte subsets and their PD-1 expression during acute HIV-1 infection. BMC Immunol. 2019, 20, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockerham, L.R.; Siliciano, J.D.; Sinclair, E.; O’Doherty, U.; Palmer, S.; Yukl, S.A.; Strain, M.C.; Chomont, N.; Hecht, F.M.; Siliciano, R.F.; et al. CD4+ and CD8+ T cell activation are associated with HIV DNA in resting CD4+ T cells. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lécuroux, C.; Girault, I.; Urrutia, A.; Doisne, J.M.; Deveau, C.; Goujard, C.; Meyer, L.; Sinet, M.; Venet, A. Identification of a particular HIV-specific CD8+ T-cell subset with a CD27+ CD45RO/RA+ phenotype and memory characteristics after initiation of HAART during acute primary HIV infection. Blood 2009, 113, 3209–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gren, S.T.; Rasmussen, T.B.; Janciauskiene, S.; Håkansson, K.; Gerwien, J.G.; Grip, O. A Single-Cell Gene-Expression Profile Reveals Inter-Cellular Heterogeneity within Human Monocyte Subsets. PLoS ONE 2015, 10, e0144351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ancuta, P.; Rao, R.; Moses, A.; Mehle, A.; Shaw, S.K.; Luscinskas, F.W.; Gabuzda, D. Fractalkine preferentially mediates arrest and migration of CD16+ monocytes. J. Exp. Med. 2003, 197, 1701–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimen, M.; Yates, C.M.; McGettrick, H.M.; Ward, L.S.C.; Harrison, M.J.; Apta, B.; Dib, L.H.; Imhof, B.A.; Harrison, P.; Nash, G.B.; et al. Monocyte Subsets Coregulate Inflammatory Responses by Integrated Signaling through TNF and IL-6 at the Endothelial Cell Interface. J. Immunol. 2017, 198, 2834–2843. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.L.; Tai, J.J.-Y.; Wong, W.-C.; Han, H.; Sem, X.; Yeap, W.-H.; Kourilsky, P.; Wong, S.-C. Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood 2011, 118, e16–e31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micheletti, A.; Finotti, G.; Calzetti, F.; Lonardi, S.; Zoratti, E.; Bugatti, M.; Stefini, S.; Vermi, W.; Cassatella, M.A. slan/M-DC8+ cells constitute a distinct subset of dendritic cells in human tonsils. Oncotarget 2016, 7, 161–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofer, T.P.; van de Loosdrecht, A.A.; Stahl-Hennig, C.; Cassatella, M.A.; Ziegler-Heitbrock, L. 6-Sulfo LacNAc (Slan) as a Marker for Non-classical Monocytes. Front. Immunol. 2019, 10, 2052. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zou, W.; Du, J.; Zhao, Y. The origins and homeostasis of monocytes and tissue-resident macrophages in physiological situation. J. Cell. Physiol. 2018, 233, 6425–6439. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the full spectrum of macrophage activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Parisi, L.; Gini, E.; Baci, D.; Tremolati, M.; Fanuli, M.; Bassani, B.; Farronato, G.; Bruno, A.; Mortara, L. Macrophage Polarization in Chronic Inflammatory Diseases: Killers or Builders? J. Immunol. Res. 2018, 2018. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, A.C.; Farias, G.B.; Gomes, A.M.C.; Godinho-Santos, A.; Rosmaninho, P.; Conceição, C.M.; Laia, J.; Santos, D.F.; Almeida, A.R.M.; Mota, C.; et al. Severe COVID-19 Recovery Is Associated with Timely Acquisition of a Myeloid Cell Immune-Regulatory Phenotype. Front. Immunol. 2021, 12, 691725. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, H.; Budoff, M.; Haberlen, S.A.; Rezaeian, P.; Ketlogetswe, K.; Tracy, R.; Palella, F.; Witt, M.D.; McConnell, M.V.; Kingsley, L.; et al. Inflammatory markers associated with subclinical coronary artery disease: The multicenter AIDS cohort study. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieblemont, N.; Weiss, L.; Sadeghi, H.M.; Estcourt, C.; Haeffner-Cavaillon, N. CD14lowCD16high: A cytokine-producing monocyte subset which expands during human immunodeficiency virus infection. Eur. J. Immunol. 1995, 25, 3418–3424. [Google Scholar] [CrossRef]
- Anzinger, J.J.; Butterfield, T.R.; Angelovich, T.A.; Crowe, S.M.; Palmer, C.S. Monocytes as regulators of inflammation and HIV-related comorbidities during cART. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Su, B.; Zhang, T.; Zhu, X.; Xia, W.; Fu, Y.; Zhao, G.; Xia, H.; Dai, L.; Sun, L.; et al. Perturbations of monocyte subsets and their association with T helper cell differentiation in acute and chronic HIV-1-infected patients. Front. Immunol. 2017, 8, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazer, S.W.; Aicher, T.P.; Muema, D.M.; Carroll, S.L.; Ordovas-Montanes, J.; Miao, V.N.; Tu, A.A.; Ziegler, C.G.K.; Nyquist, S.K.; Wong, E.B.; et al. Integrated single-cell analysis of multicellular immune dynamics during hyperacute HIV-1 infection. Nat. Med. 2020, 26, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novelli, S.; Lécuroux, C.; Goujard, C.; Reynes, J.; Villemant, A.; Blum, L.; Essat, A.; Avettand-Fenoël, V.; Launay, O.; Molina, J.M.; et al. Persistence of monocyte activation under treatment in people followed since acute HIV-1 infection relative to participants at high or low risk of HIV infection. EBioMedicine 2020, 62. [Google Scholar] [CrossRef] [PubMed]
- Pastor, L.; Parker, E.; Carrillo, J.; Urrea, V.; Fuente-Soro, L.; Respeito, D.; Jairoce, C.; Mandomando, I.; Blanco, J.; Naniche, D. A cytokine pattern that differentiates preseroconversion from postseroconversion phases of primary HIV infection. JAIDS J. Acquir. Immune Defic. Syndr. 2017, 74, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhems, P.; Lambert, J.; Cooper, D.A.; Perrin, L.; Carr, A.; Hirschel, B.; Vizzard, J.; Kinloch-De Loës, S.; Allard, R. Severity and prognosis of acute human immunodeficiency virus type 1 illness: A dose-response relationship. Clin. Infect. Dis. 1998, 26, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Fiebig, E.W.; Wright, D.J.; Rawal, B.D.; Garrett, P.E.; Schumacher, R.T.; Peddada, L.; Heldebrant, C.; Smith, R.; Conrad, A.; Kleinman, S.H.; et al. Dynamics of HIV viremia and antibody seroconversion in plasma donors. AIDS 2003, 17, 1871–1879. [Google Scholar] [CrossRef]
- Melsen, J.E.; van Ostaijen-ten Dam, M.M.; Lankester, A.C.; Schilham, M.W.; van den Akker, E.B. A Comprehensive Workflow for Applying Single-Cell Clustering and Pseudotime Analysis to Flow Cytometry Data. J. Immunol. 2020, 205, 864–871. [Google Scholar] [CrossRef]
- McInnes, L.; Healy, J.; Melville, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. Arxiv Prepr. 2018, arXiv:1802.03426. [Google Scholar]
- Samusik, N.; Good, Z.; Spitzer, M.H.; Davis, K.L.; Nolan, G.P. Automated mapping of phenotype space with single-cell data. Nat. Methods 2016, 13, 493–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Gassen, S.; Callebaut, B.; Van Helden, M.J.; Lambrecht, B.N.; Demeester, P.; Dhaene, T.; Saeys, Y. FlowSOM: Using Self-Organizing Maps for Visualization and Interpretation of Cytometry Data. Cytom. Part A 2015, 87, 636–645. [Google Scholar] [CrossRef]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Kapellos, T.S.; Bonaguro, L.; Gemünd, I.; Reusch, N.; Saglam, A.; Hinkley, E.R.; Schultze, J.L. Human Monocyte Subsets and Phenotypes in Major Chronic Inflammatory Diseases. Front. Immunol. 2019, 10, 2035. [Google Scholar] [CrossRef] [Green Version]
- Dutertre, C.A.; Amraoui, S.; DeRosa, A.; Jourdain, J.P.; Vimeux, L.; Goguet, M.; Degrelle, S.; Feuillet, V.; Liovat, A.S.; Müller-Trutwin, M.; et al. Pivotal role of M-DC8+ monocytes from viremic HIV-infected patients in TNFα overproduction in response to microbial products. Blood 2012, 120, 2259–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tufa, D.M.; Ahmad, F.; Chatterjee, D.; Ahrenstorf, G.; Schmidt, R.E.; Jacobs, R. Brief Report: HIV-1 Infection Results in Increased Frequency of Active and Inflammatory SlanDCs that Produce High Level of IL-1β. JAIDS J. Acquir. Immune Defic. Syndr. 2016, 73, 34–38. [Google Scholar] [CrossRef]
- Zhou, R.; To, K.K.W.; Wong, Y.C.; Liu, L.; Zhou, B.; Li, X.; Huang, H.; Mo, Y.; Luk, T.Y.; Lau, T.T.K.; et al. Acute SARS-CoV-2 Infection Impairs Dendritic Cell and T Cell Responses. Immunity 2020, 53, 864–877.e5. [Google Scholar] [CrossRef]
- Titanji, K.; De Milito, A.; Cagigi, A.; Thorstensson, R.; Grützmeier, S.; Atlas, A.; Hejdeman, B.; Kroon, F.P.; Lopalco, L.; Nilsson, A.; et al. Loss of memory B cells impairs maintenance of long-term serologic memory during HIV-1 infection. Blood 2006, 108, 1580–1587. [Google Scholar] [CrossRef] [Green Version]
- Samji, H.; Cescon, A.; Hogg, R.S.; Modur, S.P.; Althoff, K.N.; Buchacz, K.; Burchell, A.N.; Cohen, M.; Gebo, K.A.; Gill, M.J.; et al. Closing the gap: Increases in life expectancy among treated HIV-positive individuals in the United States and Canada. PLoS ONE 2013, 8, e81355. [Google Scholar] [CrossRef]
- Biancotto, A.; Iglehart, S.J.; Vanpouille, C.; Condack, C.E.; Lisco, A.; Ruecker, E.; Hirsch, I.; Margolis, L.B.; Grivel, J.C. HIV-1-induced activation of CD4+ T cells creates new targets for HIV-1 infection in human lymphoid tissue ex vivo. Blood 2008, 111, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Dentener, M.A.; Bazil, V.; Von Asmuth, E.J.; Ceska, M.; Buurman, W.A. Involvement of CD14 in lipopolysaccharide-induced tumor necrosis factor-alpha, IL-6 and IL-8 release by human monocytes and alveolar macrophages. J. Immunol. 1993, 150, 2885–2891. [Google Scholar] [PubMed]
- Sousa, A.E.; Carneiro, J.; Meier-Schellersheim, M.; Grossman, Z.; Victorino, R.M.M. CD4 T Cell Depletion Is Linked Directly to Immune Activation in the Pathogenesis of HIV-1 and HIV-2 but Only Indirectly to the Viral Load. J. Immunol. 2002, 169, 3400–3406. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440. [Google Scholar] [CrossRef]
- Zingaropoli, M.A.; Nijhawan, P.; Carraro, A.; Pasculli, P.; Zuccalà, P.; Perri, V.; Marocco, R.; Kertusha, B.; Siccardi, G.; Del Borgo, C.; et al. Increased sCD163 and sCD14 Plasmatic Levels and Depletion of Peripheral Blood Pro-Inflammatory Monocytes, Myeloid and Plasmacytoid Dendritic Cells in Patients With Severe COVID-19 Pneumonia. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Anbazhagan, K.; Duroux-Richard, I.; Jorgensen, C.; Apparailly, F. Transcriptomic Network Support Distinct Roles of Classical and Non-Classical Monocytes in Human. Int. Rev. Immunol. 2014, 33, 470–489. [Google Scholar] [CrossRef] [PubMed]
- Schäkel, K.; von Kietzell, M.; Hänsel, A.; Ebling, A.; Schulze, L.; Haase, M.; Semmler, C.; Sarfati, M.; Barclay, A.N.; Randolph, G.J.; et al. Human 6-Sulfo LacNAc-Expressing Dendritic Cells Are Principal Producers of Early Interleukin-12 and Are Controlled by Erythrocytes. Immunity 2006, 24, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäkel, K.; Kannagi, R.; Kniep, B.; Goto, Y.; Mitsuoka, C.; Zwirner, J.; Soruri, A.; von Kietzell, M.; Rieber, E.P. 6-Sulfo LacNAc, a Novel Carbohydrate Modification of PSGL-1, Defines an Inflammatory Type of Human Dendritic Cells. Immunity 2002, 17, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Hänsel, A.; Günther, C.; Ingwersen, J.; Starke, J.; Schmitz, M.; Bachmann, M.; Meurer, M.; Rieber, E.P.; Schäkel, K. Human slan (6-sulfo LacNAc) dendritic cells are inflammatory dermal dendritic cells in psoriasis and drive strong T 17/T 1 T-cell responses. J. Allergy Clin. Immunol. 2011, 127, 787–794. [Google Scholar] [CrossRef]
- Mager, K.; Wehner, R.; Bahr, F.; Oelschlägel, U.; Platzbecker, U.; Wermke, M.; Shayegi, N.; Middeke, J.M.; Radke, J.; Röllig, C.; et al. Reconstitution of 6-sulfo LacNAc dendritic cells after allogeneic stem-cell transplantation. Transplantation 2012, 93, 1270–1275. [Google Scholar] [CrossRef]
- Grassi, F.; Hosmalin, A.; McIlroy, D.; Calvez, V.; Debré, P.; Autran, B. Depletion in blood CD11c-positive dendritic cells from HIV-infected patients. AIDS 1999, 13, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Moir, S.; Fauci, A.S. B cells in HIV infection and disease. Nat. Rev. Immunol. 2009, 9, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Muir, R.; Metcalf, T.; Tardif, V.; Takata, H.; Phanuphak, N.; Kroon, E.; Colby, D.J.; Trichavaroj, R.; Valcour, V.; Robb, M.L.; et al. Altered Memory Circulating T Follicular Helper-B Cell Interaction in Early Acute HIV Infection. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef]
- Sun, Y.; Fu, Y.; Zhang, Z.; Tang, T.; Liu, J.; Ding, H.; Han, X.; Xu, J.; Chu, Z.; Shang, H.; et al. The investigation of CD4 + T-cell functions in primary HIV infection with antiretroviral therapy. Medicine 2017, 96. [Google Scholar] [CrossRef]
- Potter, S.J.; Lacabaratz, C.; Lambotte, O.; Perez-Patrigeon, S.; Vingert, B.; Sinet, M.; Colle, J.-H.; Urrutia, A.; Scott-Algara, D.; Boufassa, F.; et al. Preserved Central Memory and Activated Effector Memory CD4+ T-Cell Subsets in Human Immunodeficiency Virus Controllers: An ANRS EP36 Study. J. Virol. 2007, 81, 13904–13915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipp, H.; Zemlin, A.E.; Erasmus, R.T.; Glashoff, R.H. Role of inflammation in HIV-1 disease progression and prognosis. Crit. Rev. Clin. Lab. Sci. 2014, 51, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Cirión, A.; Hamimi, C.; Bergamaschi, A.; David, A.; Versmisse, P.; Mélard, A.; Boufassa, F.; Barré-Sinoussi, F.; Lambotte, O.; Rouzioux, C.; et al. Restriction of HIV-1 replication in macrophages and CD4+ T cells from HIV controllers. Blood 2011, 118, 955–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| HIV-1 Acute | HIV-1 Chronic | COVID-19 * | Healthy Controls | |
|---|---|---|---|---|
| Number (males) | 1 (0) | 1 (1) | 20 (17) | 11 (9) |
| Age (years) | 22 | 49 | 55.5 (39–65) | 58 (39–65) |
| Time from symptoms start (days) | 13 | NA | 8.5 (5–11) | NA |
| CRP (mg/dL) | 2.60 | 0.37 | 8.8 (4.85–25.5) | ND |
| PCT (ng/mL) | 0.48 | ULoD | 0.16 (0.11–0.38) | ND |
| Ferritin (ng/mL) | 412 | 262 | 939 (402–1906) | ND |
| Interleukin 6 (pg/mL) | 1.5 | 2.8 | 18 (4.5–36) | 0.85 (0.24–1.6) |
| Lymphocytes/μL | 1620 | 3130 | 920 (845–1662) | 1940 (1423–2200) |
| CD4+ T cells/μL | 364 | 210 | 247 (133–392) | 768 (544–998) |
| CD8+ T cells/μL | 503 | 863 | 145 (81.2–262) | 414 (158–577) |
| CD4/CD8 ratio | 0.72 | 0.24 | 1.78 (0.93–2.50) | 1.95 (1.53–4.17) |
| Neutrophils/mL | 1830 | 2290 | 4251 (2413–6917) | 3228 (2521–6390) |
| Lymphocytes/neutrophils ratio | 0.89 | 1.37 | 0.23 (0.15–0.5) | 0.51 (0.47–0.61) |
| Monocytes/mL | 810 | 720 | 349 (223–537) | 398 (275–733) |
| Basophils/mL | 10 | 50 | 20 (9.7–35) | 32 (16–63) |
| Eosinophils/mL | 30 | 200 | 13 (6.7–56) | 115 (96–297) |
| SARS-CoV-2 Plasma Viral Load # (RNA cps/mL) | NA | NA | 112 (24–498) | NA |
| HIV-1 Plasma Viral Load (RNA cps/mL) | 1,320,000 | 57,200 | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farias, G.B.; Badura, R.; Conceição, C.M.; Gomes, A.M.C.; Godinho-Santos, A.; Laia, J.; Rosmaninho, P.; Santos, D.F.; Mota, C.; Almeida, A.R.M.; et al. Acute HIV-1 and SARS-CoV-2 Infections Share Slan+ Monocyte Depletion—Evidence from an Hyperacute HIV-1 Case Report. Viruses 2021, 13, 1805. https://doi.org/10.3390/v13091805
Farias GB, Badura R, Conceição CM, Gomes AMC, Godinho-Santos A, Laia J, Rosmaninho P, Santos DF, Mota C, Almeida ARM, et al. Acute HIV-1 and SARS-CoV-2 Infections Share Slan+ Monocyte Depletion—Evidence from an Hyperacute HIV-1 Case Report. Viruses. 2021; 13(9):1805. https://doi.org/10.3390/v13091805
Chicago/Turabian StyleFarias, Guilherme B., Robert Badura, Carolina M. Conceição, André M. C. Gomes, Ana Godinho-Santos, Joel Laia, Pedro Rosmaninho, Diana F. Santos, Catarina Mota, Afonso R. M. Almeida, and et al. 2021. "Acute HIV-1 and SARS-CoV-2 Infections Share Slan+ Monocyte Depletion—Evidence from an Hyperacute HIV-1 Case Report" Viruses 13, no. 9: 1805. https://doi.org/10.3390/v13091805