

COVID-19 Salivary Protein Profile: Unravelling Molecular Aspects of SARS-CoV-2 Infection
 , , , , and
, , , , and
Abstract
:1. Introduction
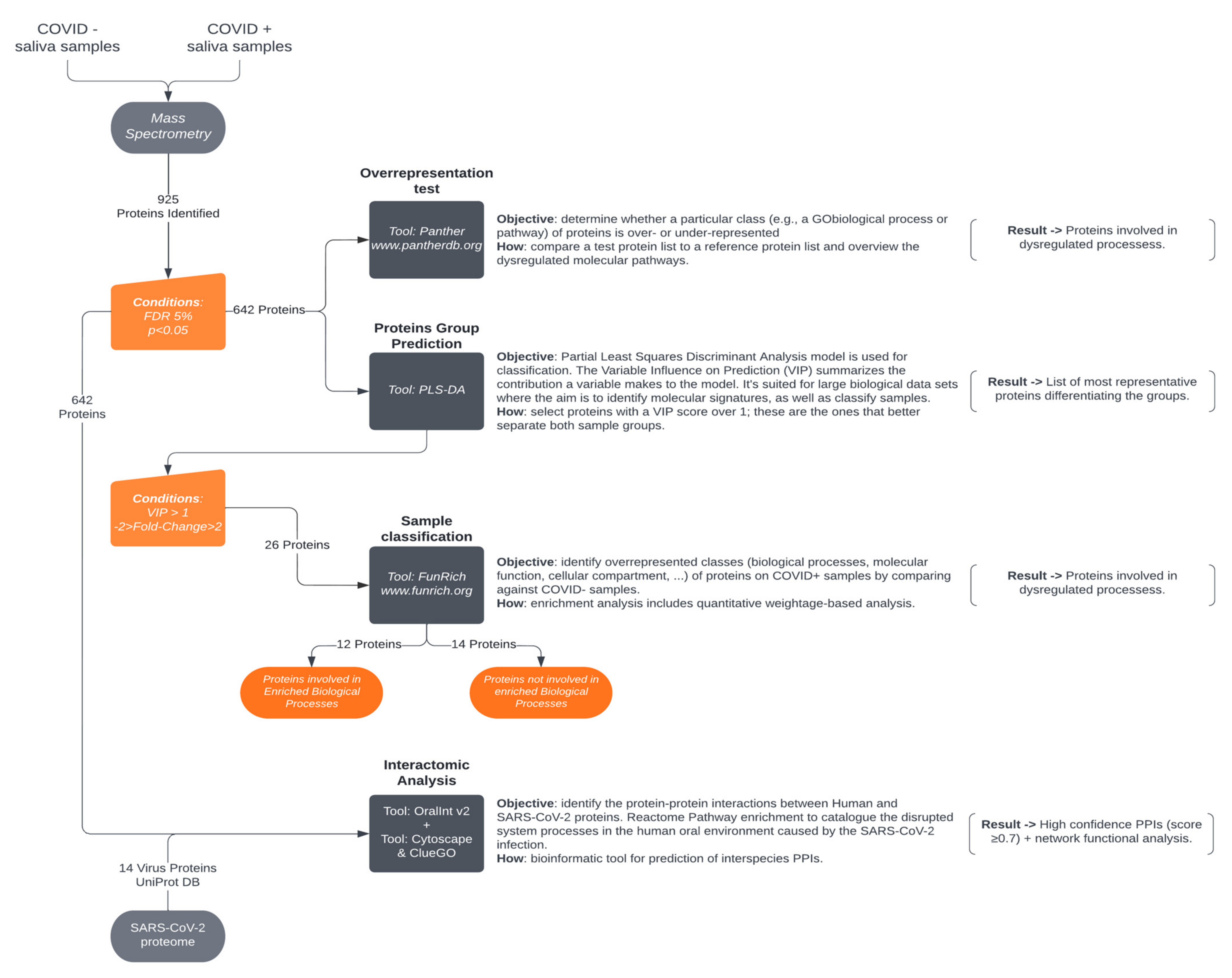
2. Materials and Methods
2.1. Ethical Statement
2.2. Participant Enrolment
2.3. Saliva Collection
2.4. Sample Preparation
2.5. LC-MS Methodology
2.6. Data Analysis
2.6.1. Ion-Library Construction (DDA Information)
2.6.2. Relative Quantification of Proteins (SWATH-MS)
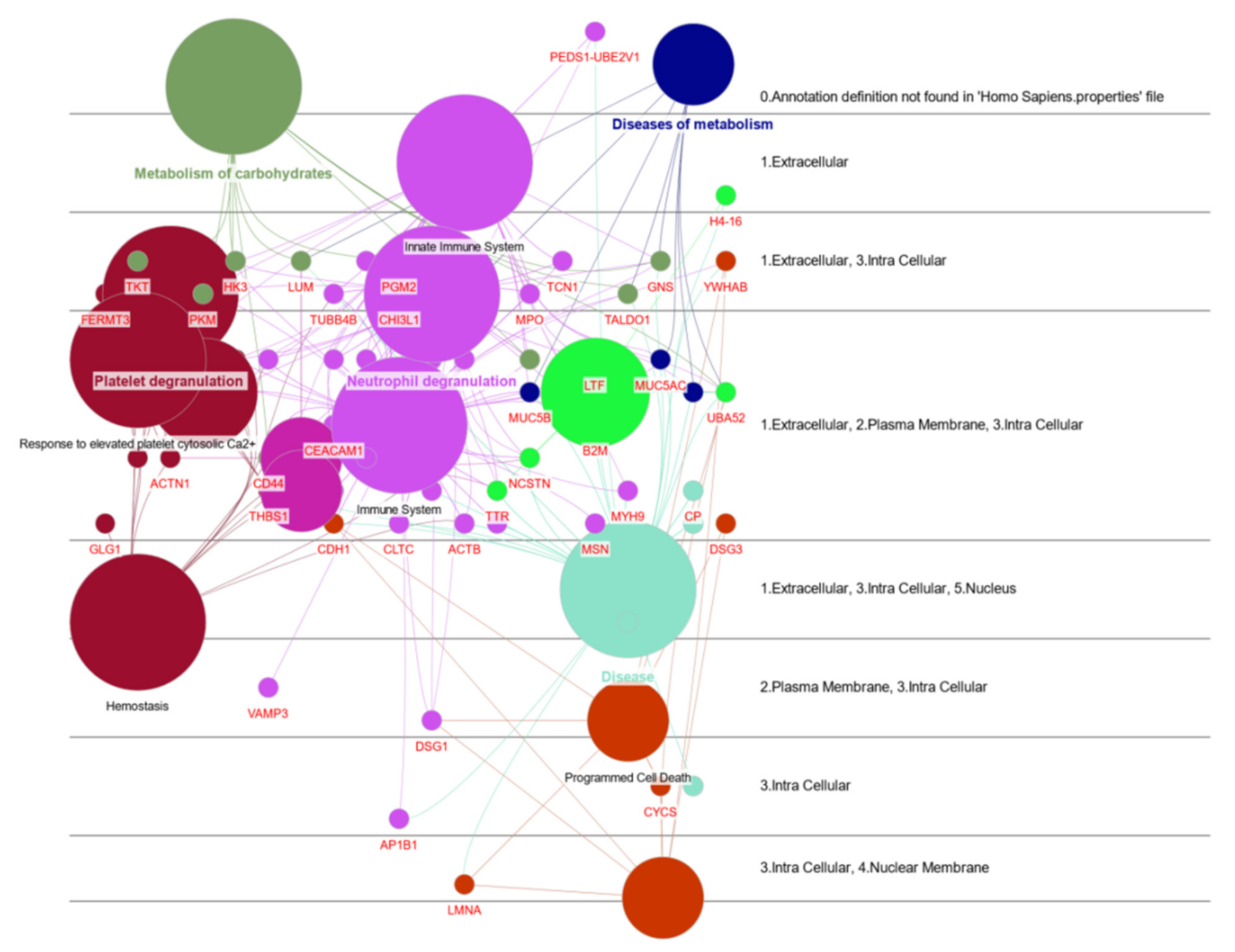
2.7. Protein Functional Analysis
2.8. Human–SARS-CoV-2 In Silico Interactomics Analysis
2.9. Statistical Analysis
3. Results and Discussion
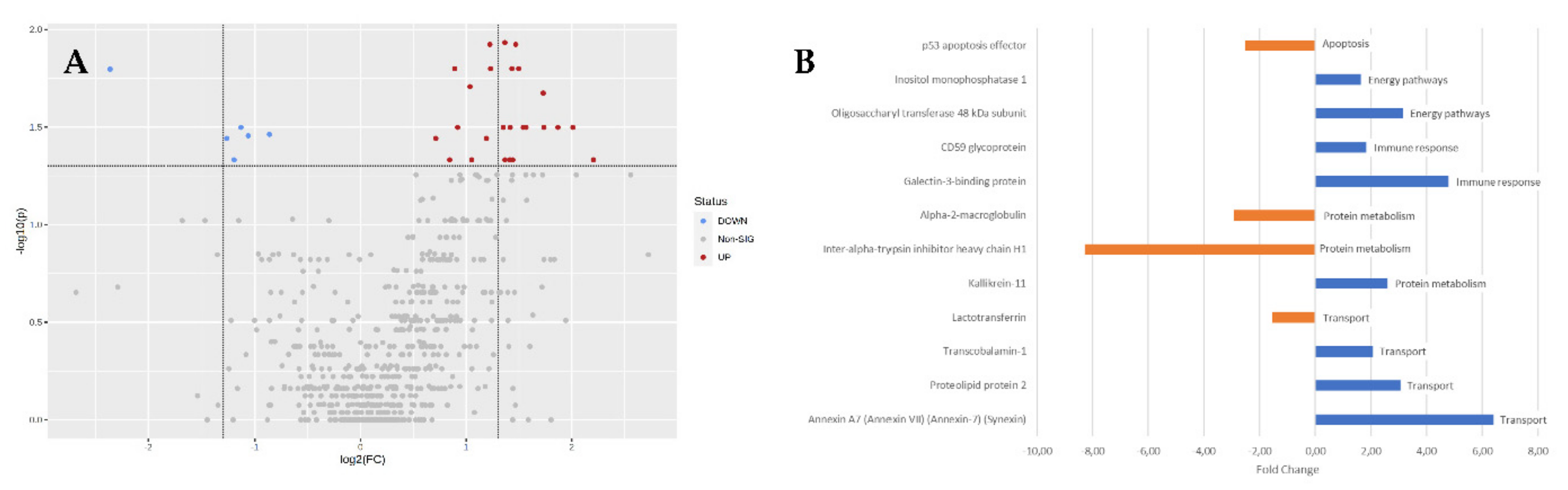
3.1. Protein Functional Analysis
3.1.1. Apoptosis
3.1.2. Energy Pathways
3.1.3. Immune Response
3.1.4. Protein Metabolism
3.1.5. Transport
3.2. Other Proteins
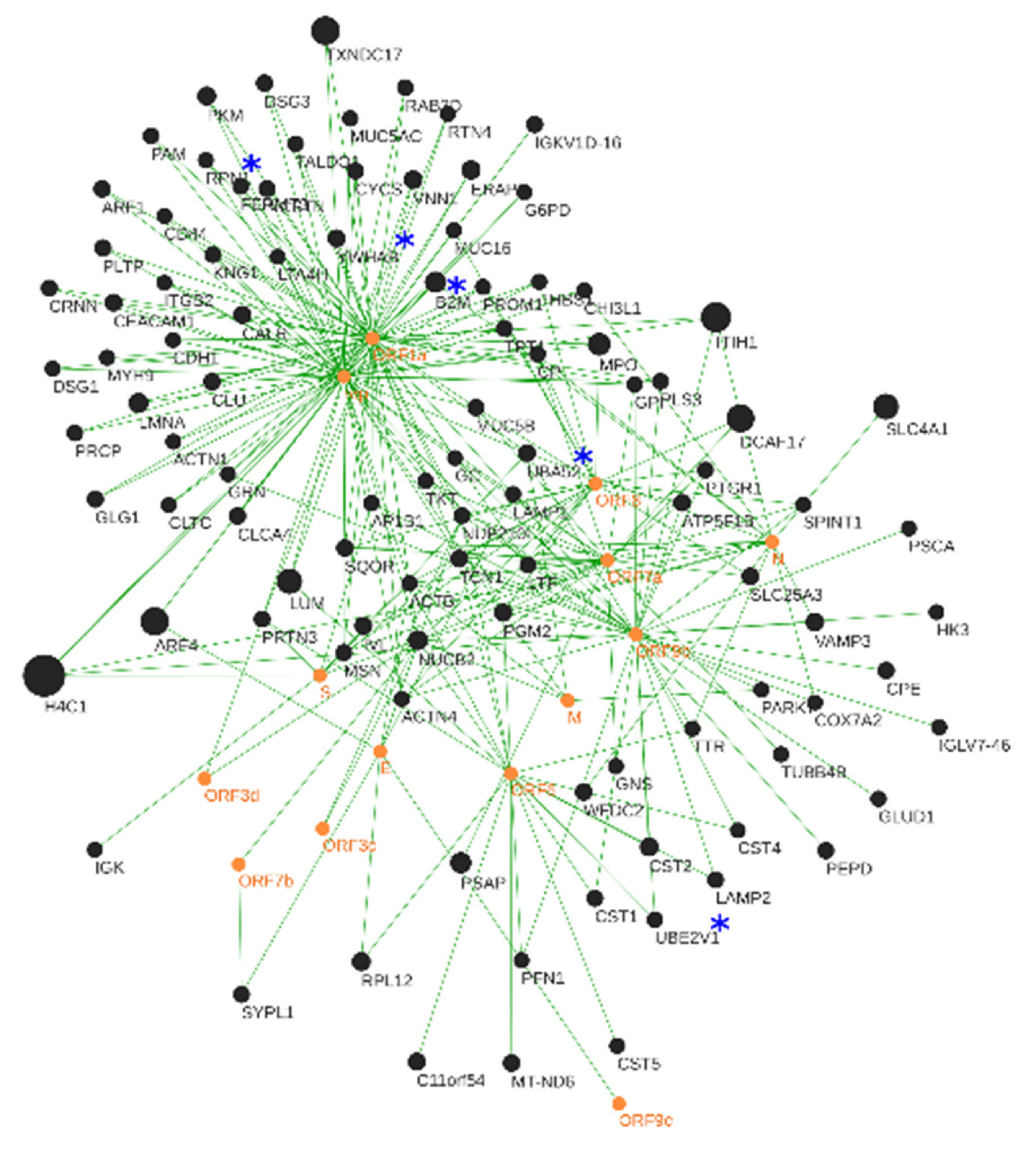
3.3. Human–SARS-CoV-2 In Silico Interactomics Analysis
3.3.1. Viral Hub Proteins
3.3.2. Human Hub Proteins
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costa, M.M.; Martin, H.; Estellon, B.; Dupé, F.-X.; Saby, F.; Benoit, N.; Tissot-Dupont, H.; Million, M.; Pradines, B.; Granjeaud, S.; et al. Exploratory Study on Application of MALDI-TOF-MS to Detect SARS-CoV-2 Infection in Human Saliva. J. Clin. Med. 2022, 11, 295. [Google Scholar] [CrossRef]
- Wandernoth, P.; Kriegsmann, K.; Groh-Mohanu, C.; Daeumer, M.; Gohl, P.; Harzer, O.; Kriegsmann, M.; Kriegsmann, J. Detection of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) by Mass Spectrometry. Viruses 2020, 12, 849. [Google Scholar] [CrossRef]
- Suvarna, K.; Biswas, D.; Pai, M.G.J.; Acharjee, A.; Bankar, R.; Palanivel, V.; Salkar, A.; Verma, A.; Mukherjee, A.; Choudhury, M.; et al. Proteomics and Machine Learning Approaches Reveal a Set of Prognostic Markers for COVID-19 Severity With Drug Repurposing Potential. Front. Physiol. 2021, 12, 652799. [Google Scholar] [CrossRef]
- Costanzo, M.; Caterino, M.; Fedele, R.; Cevenini, A.; Pontillo, M.; Barra, L.; Ruoppolo, M. COVIDomics: The Proteomic and Metabolomic Signatures of COVID-19. Int. J. Mol. Sci. 2022, 23, 2414. [Google Scholar] [CrossRef]
- Völlmy, F.; Toorn, H.V.D.; Chiozzi, R.Z.; Zucchetti, O.; Papi, A.; Volta, C.A.; Marracino, L.; Sega, F.V.D.; Fortini, F.; Demichev, V.; et al. A serum proteome signature to predict mortality in severe COVID-19 patients. Life Sci. Alliance 2021, 4, e202101099. [Google Scholar] [CrossRef]
- Castander-Olarieta, A.; Pereira, C.; Montalbán, I.A.; Mendes, V.M.; Correia, S.; Suárez-Álvarez, S.; Manadas, B.; Canhoto, J.; Moncaleán, P. Proteome-Wide Analysis of Heat-Stress in Pinus Radiata Somatic Embryos Reveals a Combined Response of Sugar Metabolism and Translational Regulation Mechanisms. Front. Plant Sci. 2021, 12, 631239. [Google Scholar] [CrossRef]
- Gisby, J.; Clarke, C.L.; Medjeral-Thomas, N.; Malik, T.H.; Papadaki, A.; Mortimer, P.M.; Buang, N.B.; Lewis, S.; Pereira, M.; Toulza, F.; et al. Longitudinal proteomic profiling of dialysis patients with COVID-19 reveals markers of severity and predictors of death. eLife 2021, 10, e64827. [Google Scholar] [CrossRef]
- Ceron, J.J.; Lamy, E.; Martinez-Subiela, S.; Lopez-Jornet, P.; Silva, F.C.; Eckersall, P.D.; Tvarijonaviciute, A. Use of Saliva for Diagnosis and Monitoring the SARS-CoV-2: A General Perspective. J. Clin. Med. 2020, 9, 1491. [Google Scholar] [CrossRef]
- Esteves, E.; Mendes, A.K.; Barros, M.; Figueiredo, C.; Andrade, J.; Capelo, J.; Novais, A.; Rebelo, C.; Soares, R.; Nunes, A.; et al. Population wide testing pooling strategy for SARS-CoV-2 detection using saliva. PLoS ONE 2022, 17, e0263033. [Google Scholar] [CrossRef]
- Arrais, J.P.; Rosa, N.; Melo, J.; Coelho, E.D.; Amaral, D.; Correia, M.J.; Barros, M.; Oliveira, J.L. OralCard: A bioinformatic tool for the study of oral proteome. Arch. Oral Biol. 2013, 58, 762–772. [Google Scholar] [CrossRef]
- Rosa, N.; Campos, B.; Esteves, A.C.; Duarte, A.S.; Correia, M.J.; Silva, R.M.; Barros, M. Tracking the functional meaning of the human oral-microbiome protein-protein interactions. Adv. Protein Chem. Struct. Biol. 2020, 121, 199–235. [Google Scholar] [CrossRef]
- Coelho, E.D.; Arrais, J.P.; Matos, S.; Pereira, C.; Rosa, N.; Correia, M.J.; Barros, M.; Oliveira, J.L. Computational prediction of the human-microbial oral interactome. BMC Syst. Biol. 2014, 8, 24. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Anjo, S.I.; Santa, C.; Manadas, B. Short GeLC-SWATH: A fast and reliable quantitative approach for proteomic screenings. Proteomics 2014, 15, 757–762. [Google Scholar] [CrossRef]
- Rosa, N.; Correia, M.J.; Arrais, J.P.; Lopes, P.; Melo, J.; Oliveira, J.L.; Barros, M. From the salivary proteome to the OralOme: Comprehensive molecular oral biology. Arch. Oral Biol. 2012, 57, 853–864. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Paolini, A.; Borella, R.; De Biasi, S.; Neroni, A.; Mattioli, M.; Tartaro, D.L.; Simonini, C.; Franceschini, L.; Cicco, G.; Piparo, A.; et al. Cell Death in Coronavirus Infections: Uncovering Its Role during COVID-19. Cells 2021, 10, 1585. [Google Scholar] [CrossRef]
- Zhang, J.; Ejikemeuwa, A.; Gerzanich, V.; Nasr, M.; Tang, Q.; Simard, J.M.; Zhao, R.Y. Understanding the Role of SARS-CoV-2 ORF3a in Viral Pathogenesis and COVID-19. Front. Microbiol. 2022, 13, 854567. [Google Scholar] [CrossRef]
- Ma-Lauer, Y.; Carbajo-Lozoya, J.; Hein, M.Y.; Müller, M.A.; Deng, W.; Lei, J.; Meyer, B.; Kusov, Y.; von Brunn, B.; Bairad, D.R.; et al. p53 down-regulates SARS coronavirus replication and is targeted by the SARS-unique domain and PLpro via E3 ubiquitin ligase RCHY1. Proc. Natl. Acad. Sci. USA 2016, 113, E5192–E5201. [Google Scholar] [CrossRef]
- Milani, D.; Caruso, L.; Zauli, E.; Al Owaifeer, A.M.; Secchiero, P.; Zauli, G.; Gemmati, D.; Tisato, V. p53/NF-kB Balance in SARS-CoV-2 Infection: From OMICs, Genomics and Pharmacogenomics Insights to Tailored Therapeutic Perspectives (COVIDomics). Front. Pharmacol. 2022, 13, 1583. [Google Scholar] [CrossRef]
- Cui, D.; Qu, R.; Liu, D.; Xiong, X.; Liang, T.; Zhao, Y. The Cross Talk Between p53 and mTOR Pathways in Response to Physiological and Genotoxic Stresses. Front. Cell Dev. Biol. 2021, 9, 3320. [Google Scholar] [CrossRef]
- Ayres, J.S. A metabolic handbook for the COVID-19 pandemic. Nat. Metab. 2020, 2, 572–585. [Google Scholar] [CrossRef]
- Wee, A.K.H. COVID-19’s toll on the elderly and those with diabetes mellitus—Is vitamin B12 deficiency an accomplice? Med. Hypotheses 2020, 146, 110374. [Google Scholar] [CrossRef]
- Davies, J.P.; Almasy, K.M.; McDonald, E.F.; Plate, L. Comparative Multiplexed Interactomics of SARS-CoV-2 and Homologous Coronavirus Nonstructural Proteins Identifies Unique and Shared Host-Cell Dependencies. ACS Infect. Dis. 2020, 6, 3174–3189. [Google Scholar] [CrossRef]
- Fabregat, A.; Sidiropoulos, K.; Viteri, G.; Marin-Garcia, P.; Ping, P.; Stein, L.; D’Eustachio, P.; Hermjakob, H. Reactome diagram viewer: Data structures and strategies to boost performance. Bioinformatics 2017, 34, 1208–1214. [Google Scholar] [CrossRef]
- Duart, G.; García-Murria, M.J.; Grau, B.; Acosta-Cáceres, J.M.; Martínez-Gil, L.; Mingarro, I. SARS-CoV-2 envelope protein topology in eukaryotic membranes. Open Biol. 2020, 10, 200209. [Google Scholar] [CrossRef]
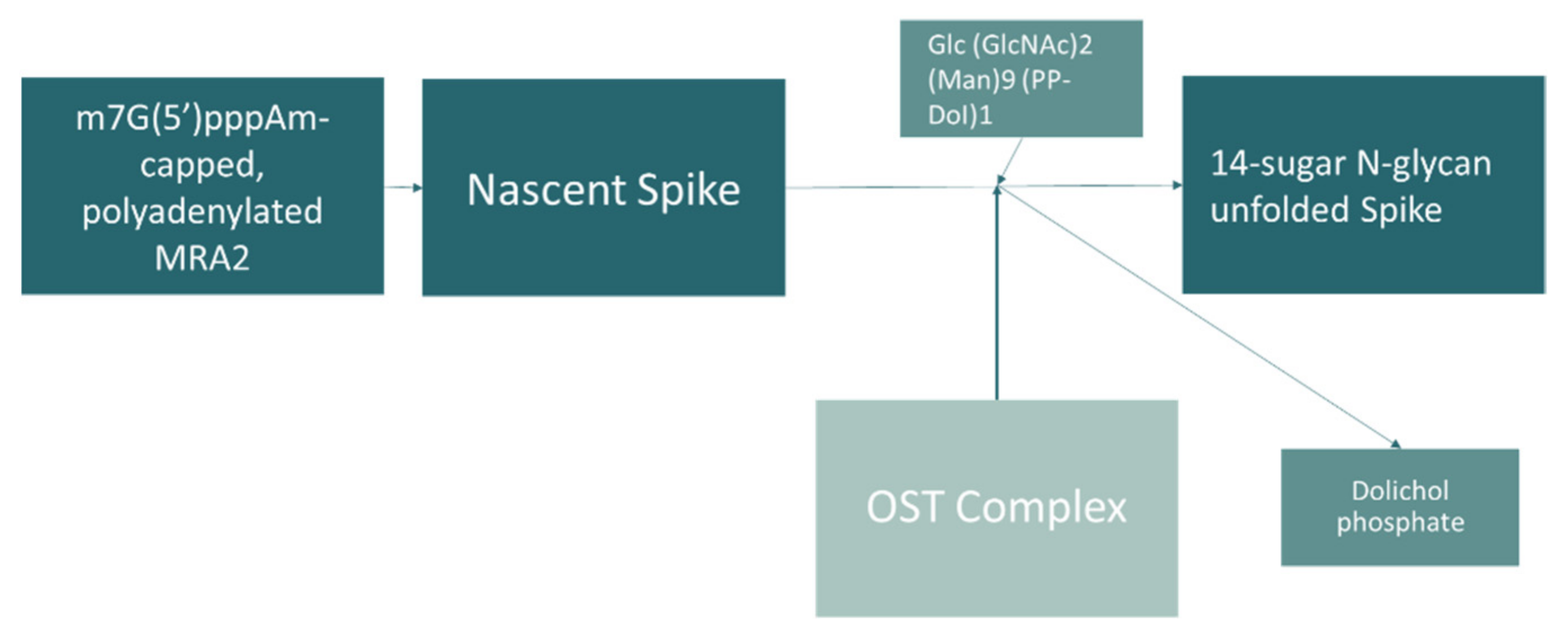
- Williams, S.J.; Goddard-Borger, E.D. α-glucosidase inhibitors as host-directed antiviral agents with potential for the treatment of COVID-19. Biochem. Soc. Trans. 2020, 48, 1287–1295. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Zhao, H.; Huang, X.; Deng, Y.-Q.; Li, X.-F.; Ye, Q.; Li, R.-T.; Xu, Y.-P.; Cao, T.-S.; Qin, C.-F. Identification of oligosaccharyltransferase as a host target for inhibition of SARS-CoV-2 and its variants. Cell Discov. 2021, 7, 116. [Google Scholar] [CrossRef]
- Rafikov, R.; McBride, M.L.; Zemskova, M.; Kurdyukov, S.; McClain, N.; Niihori, M.; Langlais, P.R.; Rafikova, O. Inositol monophosphatase 1 as a novel interacting partner of RAGE in pulmonary hypertension. Am. J. Physiol. Cell. Mol. Physiol. 2019, 316, L428–L444. [Google Scholar] [CrossRef]
- Menichini, D.; Facchinetti, F. Myo-Inositol in the Treatment of Airways Diseases: A Minireview. Int. J. Med. Device Adjuv. Treat 2021, 4, e296. [Google Scholar]
- Farahani, M.; Niknam, Z.; Amirabad, L.M.; Amiri-Dashatan, N.; Koushki, M.; Nemati, M.; Pouya, F.D.; Rezaei-Tavirani, M.; Rasmi, Y.; Tayebi, L. Molecular pathways involved in COVID-19 and potential pathway-based therapeutic targets. Biomed. Pharmacother. 2021, 145, 112420. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, Q. Cellular Immune Response to COVID-19 and Potential Immune Modulators. Front. Immunol. 2021, 12, 646333. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Sun, Q.; Knopf, J.; Herrmann, M.; Lin, L.; Jiang, J.; Shao, C.; Li, P.; He, X.; et al. Immune response in COVID-19: What is next? Cell Death Differ. 2022, 29, 1107–1122. [Google Scholar] [CrossRef]
- Garcia-Revilla, J.; Deierborg, T.; Venero, J.L.; Boza-Serrano, A. Hyperinflammation and Fibrosis in Severe COVID-19 Patients: Galectin-3, a Target Molecule to Consider. Front. Immunol. 2020, 11, 2069. [Google Scholar] [CrossRef]
- Caniglia, J.L.; Asuthkar, S.; Tsung, A.J.; Guda, M.R.; Velpula, K.K. Immunopathology of galectin-3: An increasingly promising target in COVID-19. F1000Research 2020, 9, 1078. [Google Scholar] [CrossRef]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Tartaro, D.L.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef]
- Cervantes-Alvarez, E.; la Rosa, N.L.-D.; la Mora, M.S.-D.; Valdez-Sandoval, P.; Palacios-Jimenez, M.; Rodriguez-Alvarez, F.; Vera-Maldonado, B.I.; Aguirre-Aguilar, E.; Escobar-Valderrama, J.M.; Alanis-Mendizabal, J.; et al. Galectin-3 as a potential prognostic biomarker of severe COVID-19 in SARS-CoV-2 infected patients. Sci. Rep. 2022, 12, 1856. [Google Scholar] [CrossRef]
- Kalfaoglu, B.; Almeida-Santos, J.; Tye, C.A.; Satou, Y.; Ono, M. T-Cell Hyperactivation and Paralysis in Severe COVID-19 Infection Revealed by Single-Cell Analysis. Front. Immunol. 2020, 11, 589380. [Google Scholar] [CrossRef]
- Wei, Y.; Ji, Y.; Guo, H.; Zhi, X.; Han, S.; Zhang, Y.; Gao, Y.; Chang, Y.; Yanquan, W.; Li, K.; et al. CD59 association with infectious bronchitis virus particles protects against antibody-dependent complement-mediated lysis. J. Gen. Virol. 2017, 98, 2725–2730. [Google Scholar] [CrossRef]
- Biswas, C. Chapter 18—Inflammation in Systemic Immune Diseases: Role of TLR9 Signaling and the Resultant Oxidative Stress in Pathology of Lupus. In Immunity and Inflammation in Health and Disease; Chatterjee, S., Jungraithmayr, W., Bagchi, D., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 223–237. ISBN 9780128054178. [Google Scholar]
- Barnum, S.R.; Schein, T.N. Chapter 2—The Complement System. In The Complement FactsBook, 2nd ed.; Barnum, S., Schein, T., Eds.; Factsbook; Academic Press: Cambridge, MA, USA, 2018; pp. 7–20. ISBN 9780128104200. [Google Scholar]
- Ramlall, V.; Thangaraj, P.M.; Meydan, C.; Foox, J.; Butler, D.; Kim, J.; May, B.; De Freitas, J.K.; Glicksberg, B.S.; Mason, C.E.; et al. Immune complement and coagulation dysfunction in adverse outcomes of SARS-CoV-2 infection. Nat. Med. 2020, 26, 1609–1615. [Google Scholar] [CrossRef]
- Santiesteban-Lores, L.E.; Amamura, T.A.; da Silva, T.F.; Midon, L.M.; Carneiro, M.C.; Isaac, L.; Bavia, L. A double edged-sword—The Complement System during SARS-CoV-2 infection. Life Sci. 2021, 272, 119245. [Google Scholar] [CrossRef]
- Stachowska, E.; Folwarski, M.; Jamioł-Milc, D.; Maciejewska, D.; Skonieczna-Żydecka, K. Nutritional Support in Coronavirus 2019 Disease. Medicina 2020, 56, 289. [Google Scholar] [CrossRef]
- de Carvalho, P.R.; Sirois, P.; Fernandes, P.D. The role of kallikrein-kinin and renin-angiotensin systems in COVID-19 infection. Peptides 2020, 135, 170428. [Google Scholar] [CrossRef]
- Lipcsey, M.; Persson, B.; Eriksson, O.; Blom, A.M.; Fromell, K.; Hultström, M.; Huber-Lang, M.; Ekdahl, K.N.; Frithiof, R.; Nilsson, B. The Outcome of Critically Ill COVID-19 Patients Is Linked to Thromboinflammation Dominated by the Kallikrein/Kinin System. Front. Immunol. 2021, 12, 627579. [Google Scholar] [CrossRef]
- Słomka, A.; Kowalewski, M.; Żekanowska, E. Coronavirus Disease 2019 (COVID–19): A Short Review on Hematological Manifestations. Pathogens 2020, 9, 493. [Google Scholar] [CrossRef]
- Demichev, V.; Tober-Lau, P.; Lemke, O.; Nazarenko, T.; Thibeault, C.; Whitwell, H.; Röhl, A.; Freiwald, A.; Szyrwiel, L.; Ludwig, D.; et al. A time-resolved proteomic and prognostic map of COVID-19. Cell Syst. 2021, 12, 780–794. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, X.; Ge, W.; Wang, D.; Zhu, G.; Qian, L.; Xiang, N.; Yue, L.; Liang, S.; Zhang, F.; et al. Potential Use of Serum Proteomics for Monitoring COVID-19 Progression to Complement RT-PCR Detection. J. Proteome Res. 2021, 21, 90–100. [Google Scholar] [CrossRef]
- Meier, U.-C.; Billich, A.; Mann, K.; Schramm, H.J.; Schramm, W. α2-Macroglobulin is Cleaved by HIV-1 Protease in the Bait Region but not in the C-Terminal Inter-Domain Region. Biol. Chem. Hoppe-Seyler 1991, 372, 1051–1056. [Google Scholar] [CrossRef]
- Okada, Y. Proteinases and Matrix Degradation. In Kelley and Firestein’s Textbook of Rheumatology; Elsevier: Philadelphia, PA, USA, 2017; pp. 106–125. ISBN 978-0-323-31696-5. [Google Scholar]
- Seitz, R.; Gürtler, L.; Schramm, W. Thromboinflammation in COVID-19: Can α2-macroglobulin help to control the fire? J. Thromb. Haemost. 2020, 19, 351–354. [Google Scholar] [CrossRef]
- Schramm, W.; Seitz, R.; Gürtler, L. COVID-19-associated Coagulopathy—Hypothesis: Are Children Protected Due to Enhanced Thrombin Inhibition by Higher A2-Macroglobulin (A2-M)? J. Thromb. Haemost. 2020, 18, 2416. [Google Scholar] [CrossRef]
- Vandooren, J.; Itoh, Y. Alpha-2-Macroglobulin in Inflammation, Immunity and Infections. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Yi, K.; Jung, S.; Cho, G.; Seol, H.; Hong, S.; Oh, M.; Kim, H. Effects of sFlt-1 and alpha 2-macroglobulin on vascular endothelial growth factor-induced endothelin-1 upregulation in human microvascular endothelial cells. Placenta 2014, 35, 64–69. [Google Scholar] [CrossRef]
- Oguntuyo, K.Y.; Stevens, C.S.; Siddiquey, M.N.; Schilke, R.M.; Woolard, M.D.; Zhang, H.; Acklin, J.A.; Ikegame, S.; Hung, C.-T.; Lim, J.K.; et al. In Plain Sight: The Role of Alpha-1-Antitrypsin in COVID-19 Pathogenesis and Therapeutics. bioRxiv 2020. [Google Scholar] [CrossRef]
- Schneider, W.M.; Luna, J.M.; Hoffmann, H.-H.; Sánchez-Rivera, F.J.; Leal, A.A.; Ashbrook, A.W.; Le Pen, J.; Ricardo-Lax, I.; Michailidis, E.; Peace, A.; et al. Genome-Scale Identification of SARS-CoV-2 and Pan-coronavirus Host Factor Networks. Cell 2021, 184, 120–132.e14. [Google Scholar] [CrossRef]
- Chaudhry, F.; Jageka, C.; Levy, P.D.; Cerghet, M.; Lisak, R.P. Review of the COVID-19 Risk in Multiple Sclerosis. J. Cell. Immunol. 2021, 3, 68–77. [Google Scholar] [CrossRef]
- Venkatakrishnan, A.J.; Kayal, N.; Anand, P.; Badley, A.D.; Church, G.M.; Soundararajan, V. Benchmarking evolutionary tinkering underlying human–viral molecular mimicry shows multiple host pulmonary–arterial peptides mimicked by SARS-CoV-2. Cell Death Discov. 2020, 6, 1–14. [Google Scholar] [CrossRef]
- Zuniga, M.; Gomes, C.; Carsons, S.E.; Bender, M.T.; Cotzia, P.; Miao, Q.R.; Lee, D.C.; Rodriguez, A. Autoimmunity to annexin A2 predicts mortality among hospitalised COVID-19 patients. Eur. Respir. J. 2021, 58, 2100918. [Google Scholar] [CrossRef]
- PLP2 Membrane Protein Introduction—Creative Biolabs. Available online: https://www.creative-biolabs.com/plp2-membrane-protein-introduction.html (accessed on 22 July 2022).
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodríguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef]
- Schimke, L.F.; Marques, A.H.C.; Baiocchi, G.C.; Prado, C.A.d.S.; Fonseca, D.L.M.; Freire, P.P.; Plaça, D.R.; Filgueiras, I.S.; Salgado, R.C.; Jansen-Marques, G.; et al. Severe COVID-19 Shares a Common Neutrophil Activation Signature with Other Acute Inflammatory States. Cells 2022, 11, 847. [Google Scholar] [CrossRef]
- Akgun, E.; Tuzuner, M.B.; Sahin, B.; Kilercik, M.; Kulah, C.; Cakiroglu, H.N.; Serteser, M.; Unsal, I.; Baykal, A.T. Proteins associated with neutrophil degranulation are upregulated in nasopharyngeal swabs from SARS-CoV-2 patients. PLoS ONE 2020, 15, e0240012. [Google Scholar] [CrossRef]
- Itelman, E.; Wasserstrum, Y.; Segev, A.; Avaky, C.; Negru, L.; Cohen, D.; Turpashvili, N.; Anani, S.; Zilber, E.; Lasman, N.; et al. Clinical Characterization of 162 COVID-19 patients in Israel: Preliminary Report from a Large Tertiary Center. Isr. Med. Assoc. J. 2020, 22, 271–274. [Google Scholar]
- Zhong, W.; Altay, O.; Arif, M.; Edfors, F.; Doganay, L.; Mardinoglu, A.; Uhlen, M.; Fagerberg, L. Next generation plasma proteome profiling of COVID-19 patients with mild to moderate symptoms. eBioMedicine 2021, 74, 103723. [Google Scholar] [CrossRef]
- Salaris, C.; Scarpa, M.; Elli, M.; Bertolini, A.; Guglielmetti, S.; Pregliasco, F.; Blandizzi, C.; Brun, P.; Castagliuolo, I. Protective Effects of Lactoferrin against SARS-CoV-2 Infection In Vitro. Nutrients 2021, 13, 328. [Google Scholar] [CrossRef]
- Campione, E.; Lanna, C.; Cosio, T.; Rosa, L.; Conte, M.P.; Iacovelli, F.; Romeo, A.; Falconi, M.; Del Vecchio, C.; Franchin, E.; et al. Lactoferrin as Antiviral Treatment in COVID-19 Management: Preliminary Evidence. Int. J. Environ. Res. Public Health 2021, 18, 10985. [Google Scholar] [CrossRef]
- Hu, Y.; Meng, X.; Zhang, F.; Xiang, Y.; Wang, J. The in vitro antiviral activity of lactoferrin against common human coronaviruses and SARS-CoV-2 is mediated by targeting the heparan sulfate co-receptor. Emerg. Microbes Infect. 2021, 10, 317–330. [Google Scholar] [CrossRef]
- Tian, W.; Zhang, N.; Jin, R.; Feng, Y.; Wang, S.; Gao, S.; Gao, R.; Wu, G.; Tian, D.; Tan, W.; et al. Immune suppression in the early stage of COVID-19 disease. Nat. Commun. 2020, 11, 5859. [Google Scholar] [CrossRef]
- He, B.; Garmire, L. Prediction of repurposed drugs for treating lung injury in COVID-19. F1000Research 2020, 9, 609. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, D.; Du, G.; Du, R.; Zhao, J.; Jin, Y.; Fu, S.; Gao, L.; Cheng, Z.; Lu, Q.; et al. Remdesivir in adults with severe COVID-19: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet 2020, 395, 1569–1578. [Google Scholar] [CrossRef]
- Li, W.; Yue, H. Thymidine Phosphorylase Is Increased in COVID-19 Patients in an Acuity-Dependent Manner. Front. Med. 2021, 8, 653773. [Google Scholar] [CrossRef] [PubMed]
- Schuurman, A.R.; Reijnders, T.D.; Saris, A.; Moral, I.R.; Schinkel, M.; de Brabander, J.; van Linge, C.; Vermeulen, L.; Scicluna, B.P.; Wiersinga, W.J.; et al. Integrated single-cell analysis unveils diverging immune features of COVID-19, influenza, and other community-acquired pneumonia. eLife 2021, 10, e69661. [Google Scholar] [CrossRef]
- Meng, Y.; Cai, K.; Zhao, J.; Huang, K.; Ma, X.; Song, J.; Liu, Y. Transcriptional Profiling Reveals Kidney Neutrophil Heterogeneity in Both Healthy People and ccRCC Patients. J. Immunol. Res. 2021, 2021, 5598627. [Google Scholar] [CrossRef]
- Patterson, B.K.; Francisco, E.B.; Yogendra, R.; Long, E.; Pise, A.; Rodrigues, H.; Hall, E.; Herrera, M.; Parikh, P.; Guevara-Coto, J.; et al. Persistence of SARS CoV-2 S1 Protein in CD16+ Monocytes in Post-Acute Sequelae of COVID-19 (PASC) up to 15 Months Post-Infection. Front. Immunol. 2022, 12, 746021. [Google Scholar] [CrossRef]
- Lee, Y.-B.; Jung, M.; Kim, J.; Kang, M.-G.; Kwak, C.; Kim, J.-S.; Mun, J.-Y.; Rhee, H.-W. Endomembrane Systems Are Reorganized by ORF3a and Membrane (M) of SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Fraser, D.D.; Cepinskas, G.; Patterson, E.K.; Slessarev, M.; Martin, C.; Daley, M.; Patel, M.A.; Miller, M.R.; O’Gorman, D.B.; Gill, S.E.; et al. Novel Outcome Biomarkers Identified With Targeted Proteomic Analyses of Plasma From Critically Ill Coronavirus Disease 2019 Patients. Crit. Care Explor. 2020, 2, e0189. [Google Scholar] [CrossRef]
- Schmidt, N.; Lareau, C.A.; Keshishian, H.; Ganskih, S.; Schneider, C.; Hennig, T.; Melanson, R.; Werner, S.; Wei, Y.; Zimmer, M.; et al. The SARS-CoV-2 RNA–protein interactome in infected human cells. Nat. Microbiol. 2020, 6, 339–353. [Google Scholar] [CrossRef]
- Pereira, G.J.D.S.; Leão, A.H.F.F.; Erustes, A.G.; Morais, I.B.D.M.; Vrechi, T.A.D.M.; Zamarioli, L.D.S.; Pereira, C.A.S.; Marchioro, L.D.O.; Sperandio, L.P.; Lins, Í.V.F.; et al. Pharmacological Modulators of Autophagy as a Potential Strategy for the Treatment of COVID-19. Int. J. Mol. Sci. 2021, 22, 4067. [Google Scholar] [CrossRef]
- Yan, Y.-Y.; Zhou, W.-M.; Wang, Y.-Q.; Guo, Q.-R.; Zhao, F.-X.; Zhu, Z.-Y.; Xing, Y.-X.; Zhang, H.-Y.; Aljofan, M.; Jarrahi, A.M.; et al. The Potential Role of Extracellular Vesicles in COVID-19 Treatment: Opportunity and Challenge. Front. Mol. Biosci. 2021, 8, 695. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, C.; Feng, X.; Nie, L.; Tang, M.; Zhang, H.; Xiong, Y.; Swisher, S.K.; Srivastava, M.; Chen, J. Interactomes of SARS-CoV-2 and human coronaviruses reveal host factors potentially affecting pathogenesis. EMBO J. 2021, 40, e107776. [Google Scholar] [CrossRef]
- Tate, W.; Walker, M.; Sweetman, E.; Helliwell, A.; Peppercorn, K.; Edgar, C.; Blair, A.; Chatterjee, A. Molecular Mechanisms of Neuroinflammation in ME/CFS and Long COVID to Sustain Disease and Promote Relapses. Front. Neurol. 2022, 13, 936. [Google Scholar] [CrossRef]
- Mariano, G.; Farthing, R.J.; Lale-Farjat, S.L.M.; Bergeron, J.R.C. Structural Characterization of SARS-CoV-2: Where We Are, and Where We Need to Be. Front. Mol. Biosci. 2020, 7, 344. [Google Scholar] [CrossRef]
- Cornillez-Ty, C.T.; Liao, L.; Yates, J.R.; Kuhn, P.; Buchmeier, M.J. Severe Acute Respiratory Syndrome Coronavirus Nonstructural Protein 2 Interacts with a Host Protein Complex Involved in Mitochondrial Biogenesis and Intracellular Signaling. J. Virol. 2009, 83, 10314–10318. [Google Scholar] [CrossRef]
- Perrin-Cocon, L.; Diaz, O.; Jacquemin, C.; Barthel, V.; Ogire, E.; Ramière, C.; André, P.; Lotteau, V.; Vidalain, P.-O. The current landscape of coronavirus-host protein–protein interactions. J. Transl. Med. 2020, 18, 319. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Blanco, M.R.; Bruce, E.A.; Honson, D.D.; Chen, L.M.; Chow, A.; Bhat, P.; Ollikainen, N.; Quinodoz, S.A.; Loney, C.; et al. SARS-CoV-2 Disrupts Splicing, Translation, and Protein Trafficking to Suppress Host Defenses. Cell 2020, 183, 1325–1339.e21. [Google Scholar] [CrossRef]
- Thoms, M.; Buschauer, R.; Ameismeier, M.; Koepke, L.; Denk, T.; Hirschenberger, M.; Kratzat, H.; Hayn, M.; Mackens-Kiani, T.; Cheng, J.; et al. Structural basis for translational shutdown and immune evasion by the Nsp1 protein of SARS-CoV-2. Science 2020, 369, 1249–1255. [Google Scholar] [CrossRef]
- Xia, H.; Cao, Z.; Xie, X.; Zhang, X.; Chen, J.Y.-C.; Wang, H.; Menachery, V.D.; Rajsbaum, R.; Shi, P.-Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef]
- Hsieh, I.-N.; White, M.; Hoeksema, M.; Deluna, X.; Hartshorn, K. Histone H4 potentiates neutrophil inflammatory responses to influenza A virus: Down-modulation by H4 binding to C-reactive protein and Surfactant protein D. PLoS ONE 2021, 16, e0247605. [Google Scholar] [CrossRef]
- Nainu, F.; Shiratsuchi, A.; Nakanishi, Y. Induction of Apoptosis and Subsequent Phagocytosis of Virus-Infected Cells As an Antiviral Mechanism. Front. Immunol. 2017, 8, 1220. [Google Scholar] [CrossRef]
- Lohr, K.; Sardana, H.; Lee, S.; Wu, F.; Huso, D.L.; Hamad, A.R.; Chakravarti, S. Extracellular matrix protein lumican regulates inflammation in a mouse model of colitis. Inflamm. Bowel Dis. 2012, 18, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Furtmüller, P.G.; Burner, U.; Obinger, C. Reaction of Myeloperoxidase Compound I with Chloride, Bromide, Iodide, and Thiocyanate. Biochemistry 1998, 37, 17923–17930. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, L.; Horgan, C.P.; McCaffrey, M.W. Knockdown of β2-microglobulin perturbs the subcellular distribution of HFE and hepcidin. Biochem. Biophys. Res. Commun. 2009, 378, 727–731. [Google Scholar] [CrossRef]
- Girelli, D.; Marchi, G.; Busti, F.; Vianello, A. Iron metabolism in infections: Focus on COVID-19. Semin. Hematol. 2021, 58, 182–187. [Google Scholar] [CrossRef]
- Drakesmith, H.; Pasricha, S.-R.; Cabantchik, I.; Hershko, C.; Weiss, G.; Girelli, D.; Stoffel, N.; Muckenthaler, M.U.; Nemeth, E.; Camaschella, C.; et al. Vaccine efficacy and iron deficiency: An intertwined pair? Lancet Haematol. 2021, 8, e666–e669. [Google Scholar] [CrossRef]
- Conca, W.; Alabdely, M.; Albaiz, F.; Foster, M.W.; Alamri, M.; Alkaff, M.; Al-Mohanna, F.; Nagelkerke, N.; Almaghrabi, R.S. Serum β2-microglobulin levels in Coronavirus disease 2019 (COVID-19): Another prognosticator of disease severity? PLoS ONE 2021, 16, e0247758. [Google Scholar] [CrossRef]
- Zhang, N.; Ke, Y.; Zhang, L. Interplay between hepatitis C virus and ARF4. Virol. Sin. 2017, 32, 533–536. [Google Scholar] [CrossRef]
- Jeong, W.; Chang, T.-S.; Boja, E.S.; Fales, H.M.; Rhee, S.G. Roles of TRP14, a Thioredoxin-related Protein in Tumor Necrosis Factor-α Signaling Pathways. J. Biol. Chem. 2004, 279, 3151–3159. [Google Scholar] [CrossRef]
- Islam, A.; Adamik, B.; Hawari, F.I.; Ma, G.; Rouhani, F.N.; Zhang, J.; Levine, S.J. Extracellular TNFR1 Release Requires the Calcium-dependent Formation of a Nucleobindin 2-ARTS-1 Complex. J. Biol. Chem. 2006, 281, 6860–6873. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2021, 50, D543–D552. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniprotKB AC | Protein Name | Gene Name | Fold Change | Biological Process |
|---|---|---|---|---|
| P19827 | Inter-alpha-trypsin inhibitor heavy chain H1 | ITIH1 | −8.28 | * Protein metabolism |
| P01023 | Alpha-2-macroglobulin | A2M | −2.91 | * Protein metabolism |
| Q96FX8 | p53 apoptosis effector | PERP | −2.52 | Apoptosis |
| P02788 | Lactotransferrin | LTF | −1.53 | Transport |
| Q99623 | Prohibitin-2 | PHB2 | 1.61 | Mitochondrion organization |
| P29218 | Inositol monophosphatase 1 | IMPA1 | 1.64 | * Energy pathways |
| Q9Y376 | Calcium-binding protein 39 | CAB39 | 1.77 | Protein serine/threonine kinase activity |
| P13987 | CD59 glycoprotein | CD59 | 1.84 | * Immune response |
| P15153 | Ras-related C3 botulinum toxin substrate 2 | RAC2 | 1.88 | Regulation of respiratory burst |
| Q86VR7 | V-set and immunoglobulin domain-containing protein 10-like | VSIG10L | 1.99 | Cell adhesion molecule |
| P20061 | Transcobalamin-1 | TCN1 | 2.08 | * Transport |
| P01706 | Immunoglobulin lambda variable 2–11 | IGLV2–11 | 2.17 | Response to bacterium |
| P20340 | Ras-related protein Rab-6A | RAB6A | 2.18 | Antigen receptor-mediated signaling pathway |
| P58499 | Protein FAM3B | FAM3B | 2.25 | Antimicrobial response protein |
| Q9HC07 | Transmembrane protein 165 | TMEM165 | 2.42 | Humoral immune response |
| Q9UBX7 | Kallikrein-11 | KLK11 | 2.61 | * Protein metabolism |
| Q6UXB3 | Ly6/PLAUR domain-containing protein 2 | LYPD2 | 2.62 | Mitotic cell cycle |
| Q04941 | Proteolipid protein 2 | PLP2 | 3.05 | * Transport |
| P63000 | Ras-related C3 botulinum toxin substrate 1 | RAC1 | 3.06 | Regulation of cell shape |
| A0A0C4DH68 | Immunoglobulin kappa variable 2–24 | IGKV2–24 | 3.09 | Immune response |
| P39656 | Oligosaccharyl transferase 48 kDa subunit | DDOST | 3.15 | * Energy pathways |
| P0DP04 | Immunoglobulin heavy variable 3–43D | IGHV3–43D | 3.37 | Defense response to bacterium |
| Q08380 | Galectin-3 binding protein | LGALS3BP | 4.80 | * Immune response |
| P0DOX2 | Immunoglobulin alpha-2 heavy chain | 4.86 | Immune response | |
| P19971 | Thymidine phosphorylase | TYMP | 5.89 | Mitochondrial genome maintenance |
| P20073 | Annexin A7 | ANXA7 | 6.40 | * Transport |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esteves, E.; Mendes, V.M.; Manadas, B.; Lopes, R.; Bernardino, L.; Correia, M.J.; Barros, M.; Esteves, A.C.; Rosa, N. COVID-19 Salivary Protein Profile: Unravelling Molecular Aspects of SARS-CoV-2 Infection. J. Clin. Med. 2022, 11, 5571. https://doi.org/10.3390/jcm11195571
Esteves E, Mendes VM, Manadas B, Lopes R, Bernardino L, Correia MJ, Barros M, Esteves AC, Rosa N. COVID-19 Salivary Protein Profile: Unravelling Molecular Aspects of SARS-CoV-2 Infection. Journal of Clinical Medicine. 2022; 11(19):5571. https://doi.org/10.3390/jcm11195571
Chicago/Turabian StyleEsteves, Eduardo, Vera M. Mendes, Bruno Manadas, Rafaela Lopes, Liliana Bernardino, Maria José Correia, Marlene Barros, Ana Cristina Esteves, and Nuno Rosa. 2022. "COVID-19 Salivary Protein Profile: Unravelling Molecular Aspects of SARS-CoV-2 Infection" Journal of Clinical Medicine 11, no. 19: 5571. https://doi.org/10.3390/jcm11195571