
Porcine Reproductive and Respiratory Syndrome Virus Modulates the Switch of Macrophage Polarization from M1 to M2 by Upregulating MoDC-Released sCD83

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Western Blot
2.3. Measurements of TNF-α, IL-10 and sCD83
2.4. RT-qPCR
2.5. Construction of Infectious PRRSV cDNA Clones
2.6. One-Step Viral Growth Curves
2.7. Statistical Analysis
3. Results
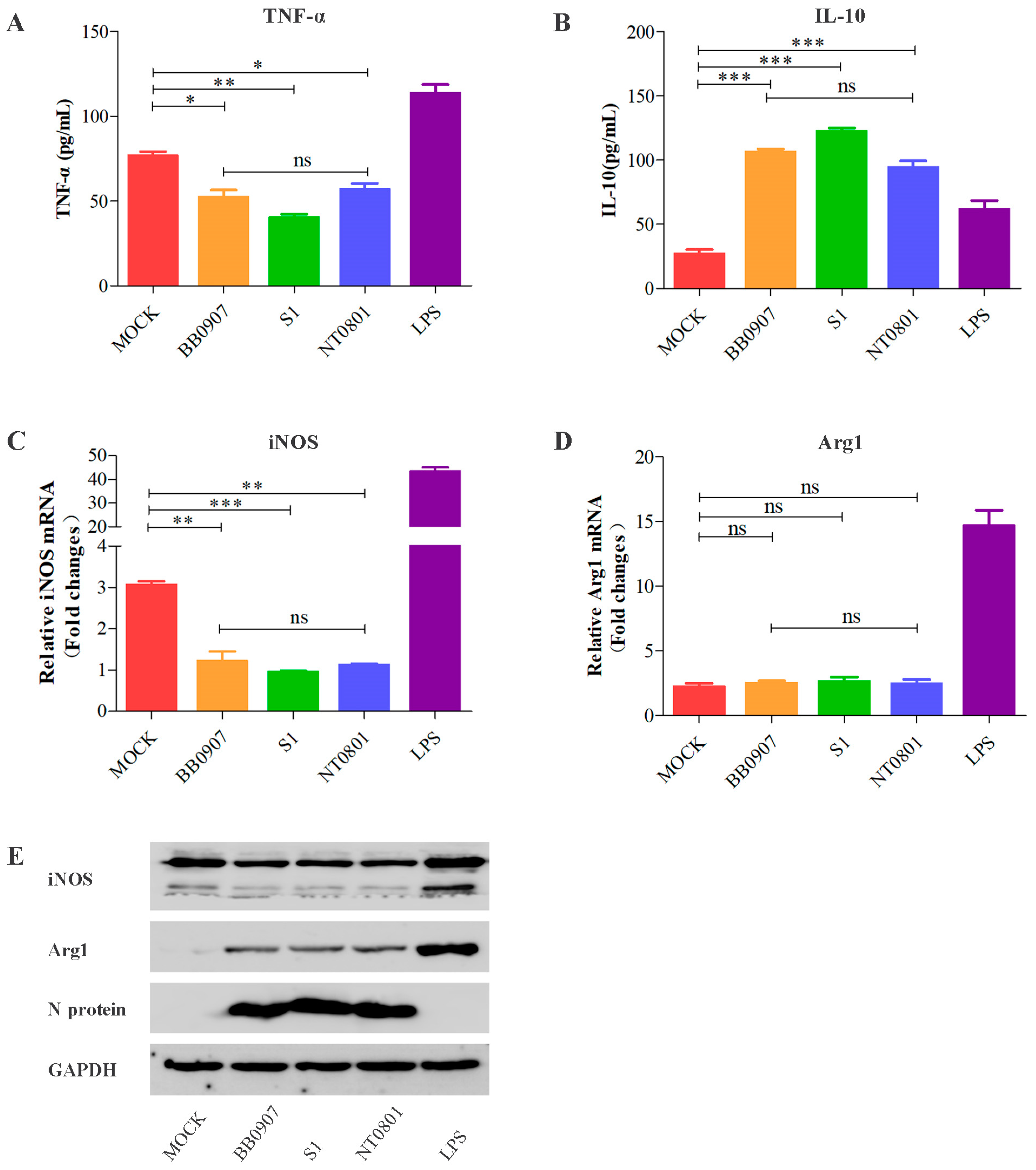
3.1. Alterations in M1 and M2 Macrophage Types after PRRSV Infection In Vitro
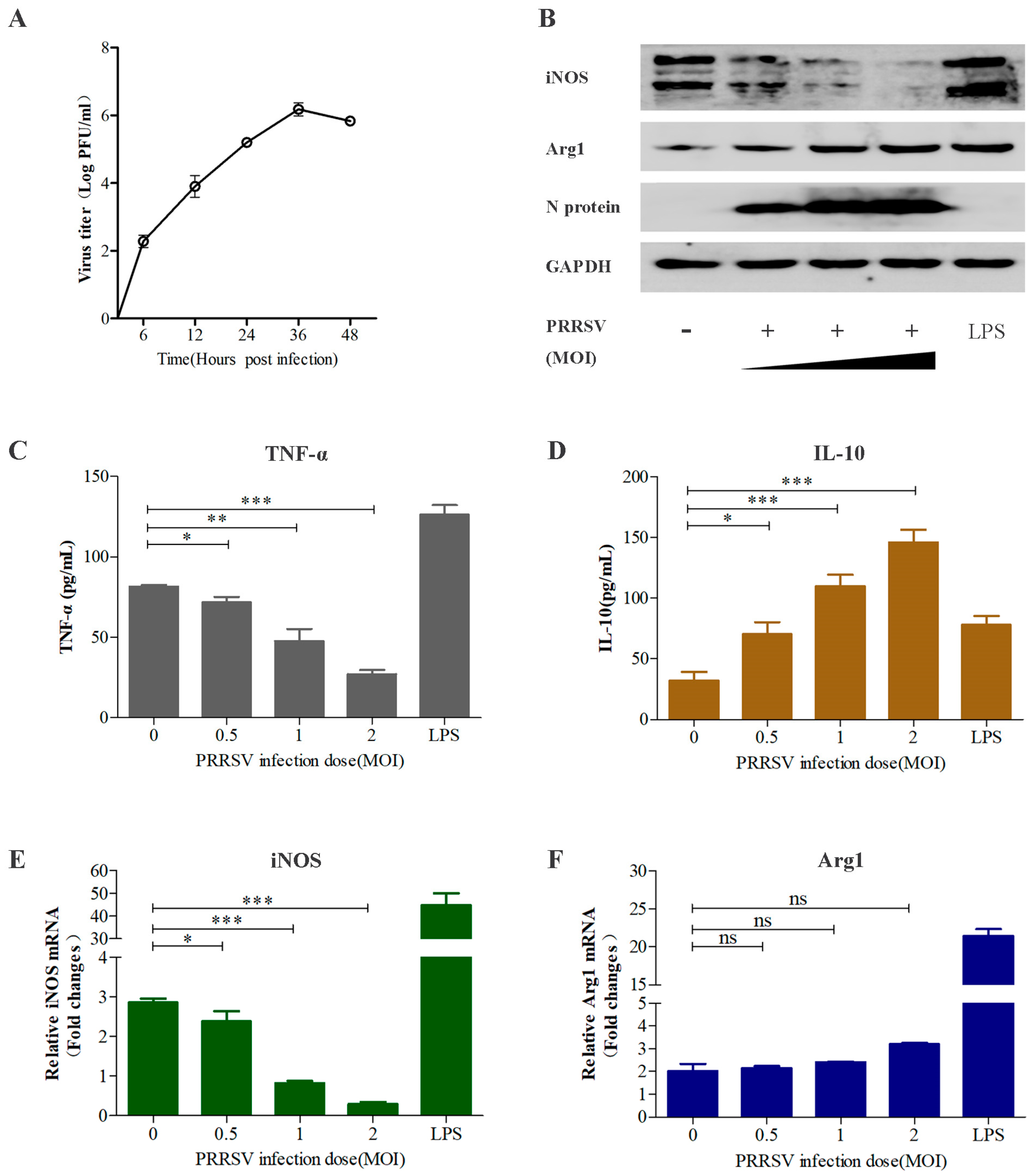
3.2. PRRSV Regulates Macrophage Polarization in a Dose-Dependent Manner
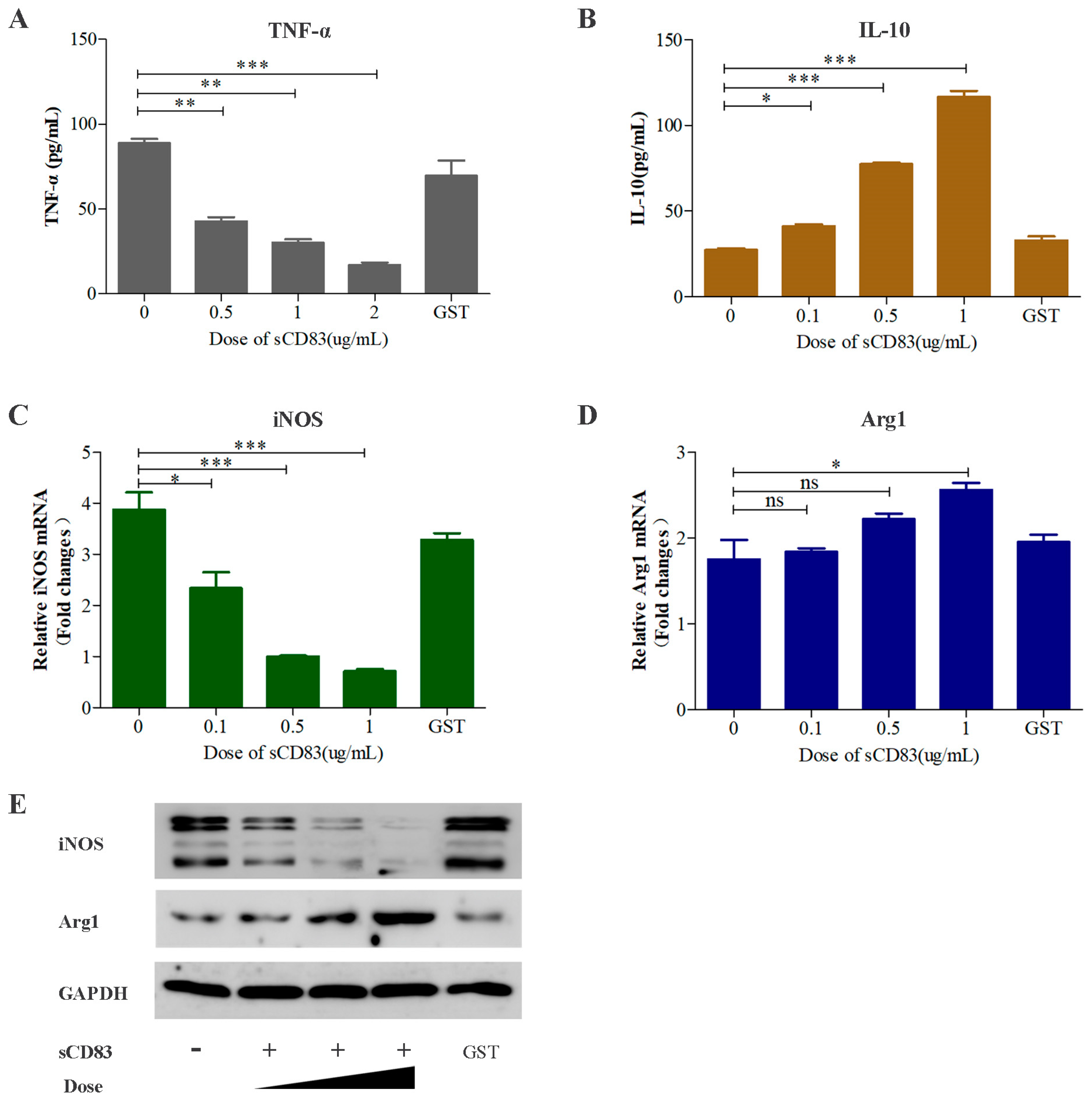
3.3. sCD83 Modulates a Switch in Macrophage Polarization from M1 to M2
3.4. Anti-CD83 Restores a Switch in Alveolar Macrophage Polarization from M1 to M2 in PRRSV Infection
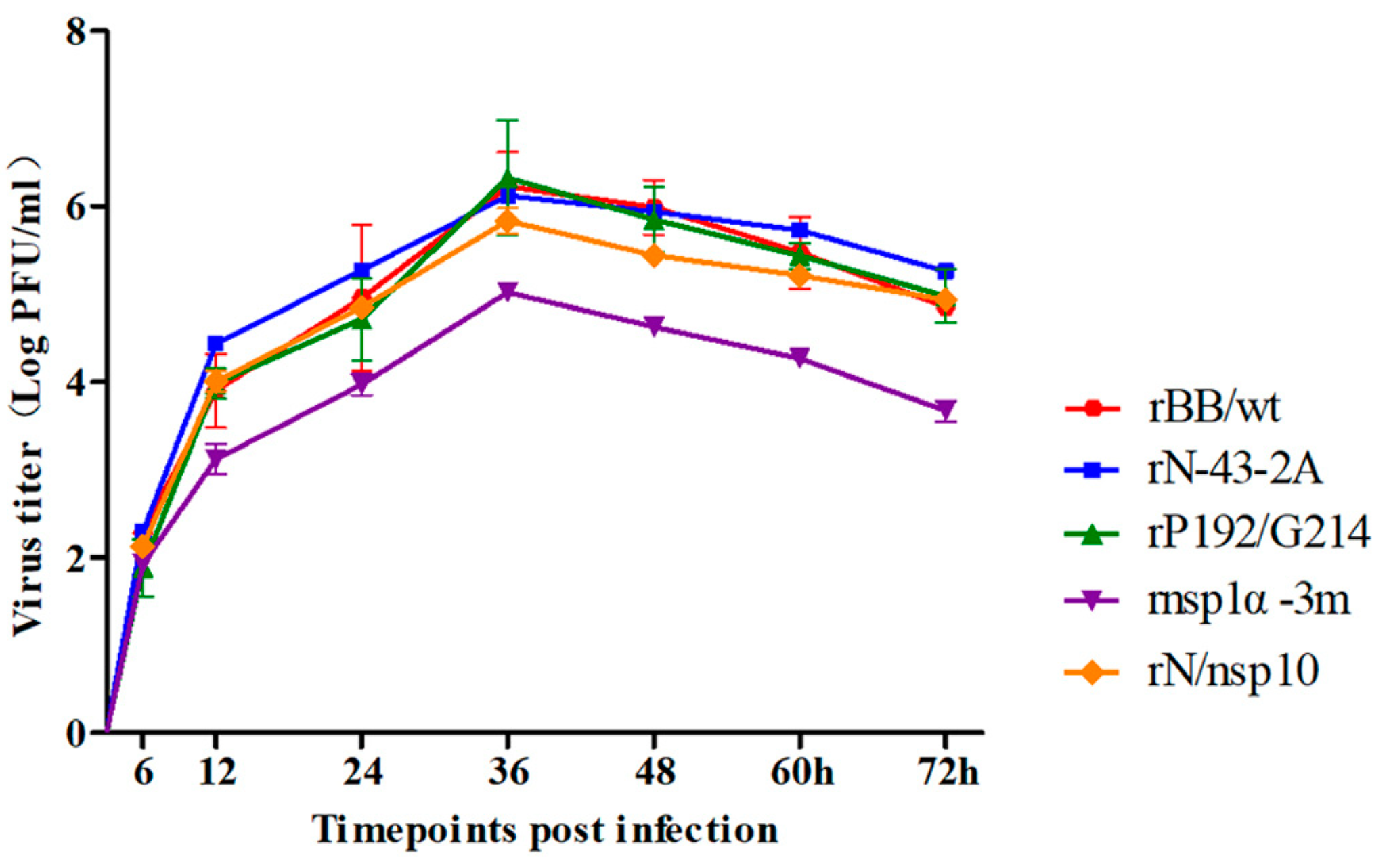
3.5. Construction and Identification of Mutant Recombinant PRRSV
3.6. Effect of Mutant Viruses on Macrophage Polarization from M1 to M2 by sCD83
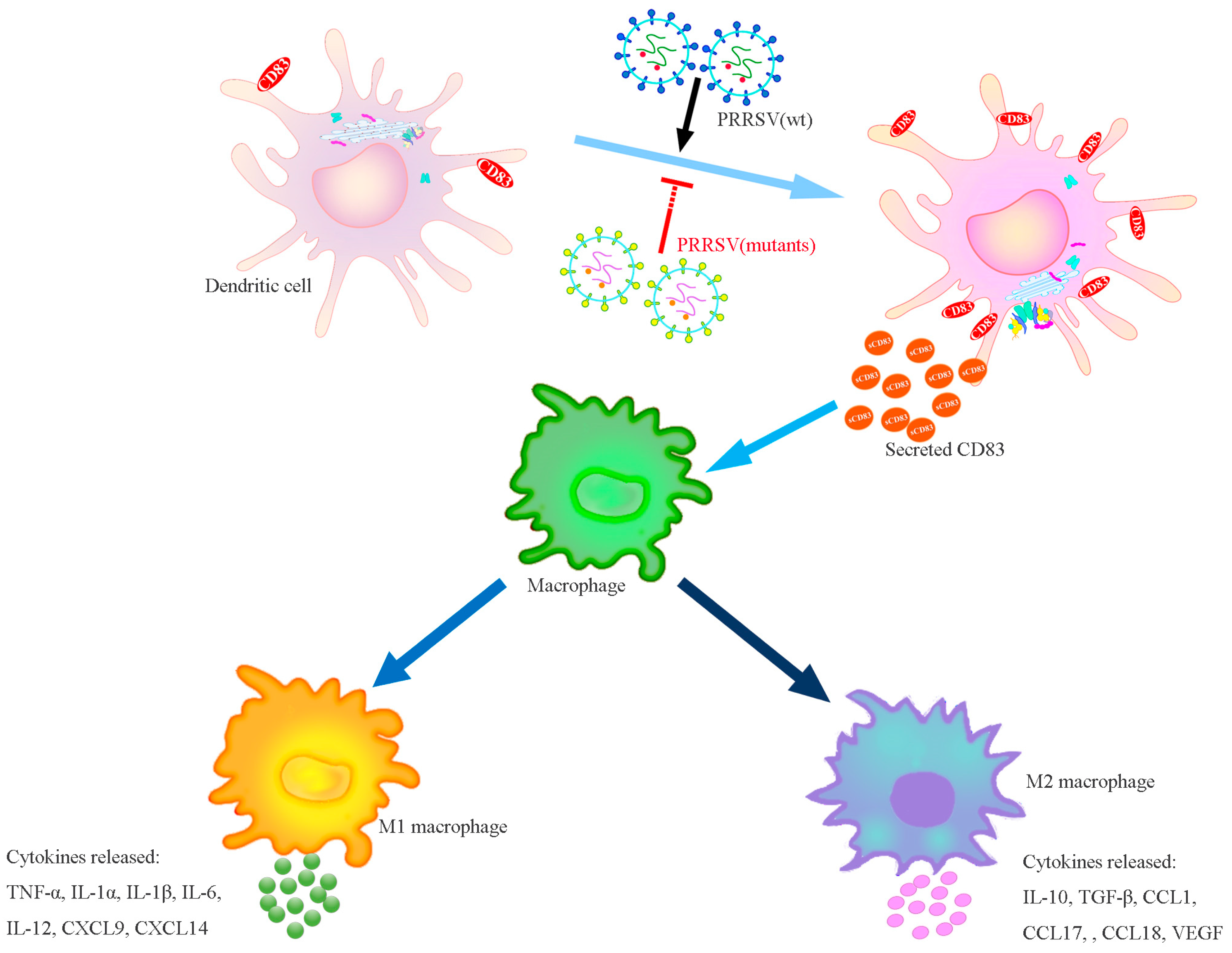
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loving, C.L.; Osorio, F.A.; Murtaugh, M.P.; Zuckermann, F.A. Innate and adaptive immunity against Porcine Reproductive and Respiratory Syndrome Virus. Vet. Immunol. Immunopathol. 2015, 167, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Li, W.; Zhao, J.; Gong, B.; Sun, Q.; Tang, Y.D.; Xiang, L.; Leng, C.; Peng, J.; et al. Novel characteristics of Chinese NADC34-like PRRSV during 2020-2021. Transbound. Emerg. Dis. 2022, 69, e3215–e3224. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zheng, S.; Han, G.; Yang, H.; Deng, Z.; Liu, Z.; He, F. Porcine Reproductive and Respiratory Syndrome Virus Adapts Antiviral Innate Immunity via Manipulating MALT1. Mbio 2022, 13, 22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sha, H.; Qin, L.; Wang, N.; Kong, W.; Huang, L.; Zhao, M. Research Progress in Porcine Reproductive and Respiratory Syndrome Virus-Host Protein Interactions. Animals 2022, 12, 1381. [Google Scholar] [CrossRef]
- Wang, T.Y.; Sun, M.X.; Zhang, H.L.; Wang, G.; Zhan, G.; Tian, Z.J.; Cai, X.H.; Su, C.; Tang, Y.D. Evasion of Antiviral Innate Immunity by Porcine Reproductive and Respiratory Syndrome Virus. Front. Microbiol. 2021, 12, 693799. [Google Scholar] [CrossRef]
- Li, Y.; Wei, Y.; Hao, W.; Zhao, W.; Zhou, Y.; Wang, D.; Xiao, S.; Fang, L. Porcine reproductive and respiratory syndrome virus infection promotes C1QBP secretion to enhance inflammatory responses. Vet. Microbiol. 2020, 241, 108563. [Google Scholar] [CrossRef]
- Chaudhari, J.; Liew, C.S.; Riethoven, J.M.; Sillman, S.; Vu, H. Porcine Reproductive and Respiratory Syndrome Virus Infection Upregulates Negative Immune Regulators and T-Cell Exhaustion Markers. J. Virol. 2021, 95, e105221. [Google Scholar] [CrossRef]
- Huang, J.; Liu, H.; Wang, M.; Bai, X.; Cao, J.; Zhang, Z.; Wang, Q. Mannosylated gelatin nanoparticles enhanced inactivated PRRSV targeting dendritic cells and increased T cell immunity. Vet. Immunol. Immunopathol. 2021, 235, 110237. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.X.; Liu, X.; Wu, H.; Wen, Y.J. Transcriptome sequencing analysis of porcine alveolar macrophages infected with PRRSV strains to elucidate virus pathogenicity and immune evasion strategies. Virusdisease 2021, 32, 559–567. [Google Scholar] [CrossRef]
- Razzuoli, E.; Armando, F.; De Paolis, L.; Ciurkiewicz, M.; Amadori, M. The Swine IFN System in Viral Infections: Major Advances and Translational Prospects. Pathogens 2022, 11, 175. [Google Scholar] [CrossRef]
- Amadori, M.; Listorti, V.; Razzuoli, E. Reappraisal of PRRS Immune Control Strategies: The Way Forward. Pathogens 2021, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Amadori, M.; Razzuoli, E. Immune Control of PRRS: Lessons to be Learned and Possible Ways Forward. Front. Vet. Sci. 2014, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bai, J.; Liu, X.; Song, Z.; Zhang, Q.; Wang, X.; Jiang, P. Nsp1alpha of Porcine Reproductive and Respiratory Syndrome Virus Strain BB0907 Impairs the Function of Monocyte-Derived Dendritic Cells via the Release of Soluble CD83. J. Virol. 2018, 92, 18. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Q.; Bai, J.; Zhao, Y.; Wang, X.; Wang, H.; Jiang, P. The Nucleocapsid Protein and Nonstructural Protein 10 of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Enhance CD83 Production via NF-kappaB and Sp1 Signaling Pathways. J. Virol. 2017, 91, 17. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Wu, F.; Zhang, J.; Wang, X.; Li, W.; Cui, D.; Zuo, Y.; Hu, M.; Zhong, F. Porcine soluble CD83 alleviates LPS-induced abortion in mice by promoting Th2 cytokine production, Treg cell generation and trophoblast invasion. Theriogenology 2020, 157, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Li, L.X.; Xia, Y.T.; Sun, X.Y.; Li, L.R.; Yao, L.; Ali, M.I.; Gu, W.; Zhang, J.P.; Liu, J.; Huang, S.G.; et al. CXCL-10/CXCR3 in macrophages regulates tissue repair by controlling the expression of Arg1, VEGFa and TNFalpha. J. Biol. Regul. Homeost. Agents 2020, 34, 987–999. [Google Scholar] [CrossRef]
- Liu, J.; Wei, S.; Liu, L.; Shan, F.; Zhao, Y.; Shen, G. The role of porcine reproductive and respiratory syndrome virus infection in immune phenotype and Th1/Th2 balance of dendritic cells. Dev. Comp. Immunol. 2016, 65, 245–252. [Google Scholar] [CrossRef]
- Wahyuningtyas, R.; Lai, Y.; Wu, M.; Chen, H.; Chung, W.; Chaung, H.; Chang, K. Recombinant Antigen of Type 2 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV-2) Promotes M1 Repolarization of Porcine Alveolar Macrophages and Th1 Type Response. Vaccines 2021, 9, 1009. [Google Scholar] [CrossRef]
- Boutilier, A.J.; Elsawa, S.F. Macrophage Polarization States in the Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 6995. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, H.; Hao, Y.; Ren, Z.; Qu, S.; Liu, L.; Wei, D.; Tang, Z.; Zhang, J.; Jiang, Z. Macrophage polarization in atherosclerosis. Clin. Chim. Acta 2020, 501, 142–146. [Google Scholar] [CrossRef]
- Orliaguet, L.; Dalmas, E.; Drareni, K.; Venteclef, N.; Alzaid, F. Mechanisms of Macrophage Polarization in Insulin Signaling and Sensitivity. Front. Endocrinol. 2020, 11, 62. [Google Scholar] [CrossRef]
- Chen, X.; Tang, J.; Shuai, W.; Meng, J.; Feng, J.; Han, Z. Macrophage polarization and its role in the pathogenesis of acute lung injury/acute respiratory distress syndrome. Inflamm. Res. 2020, 69, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J. Macrophage Polarization. Annu. Rev. Physiol. 2017, 79, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Isidro, R.A.; Appleyard, C.B. Colonic macrophage polarization in homeostasis, inflammation, and cancer. Am. J. Physiol.-Gastroint. Liver Physiol. 2016, 311, G59–G73. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zou, X.; Chai, Y.; Yao, Y. Macrophage Polarization in Inflammatory Diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef]
- Franzoni, G.; Mura, L.; Razzuoli, E.; De Ciucis, C.G.; Fruscione, F.; Dell’Anno, F.; Zinellu, S.; Carta, T.; Anfossi, A.G.; Dei, G.S.; et al. Heterogeneity of Phenotypic and Functional Changes to Porcine Monocyte-Derived Macrophages Triggered by Diverse Polarizing Factors In Vitro. Int. J. Mol. Sci. 2023, 24, 4671. [Google Scholar] [CrossRef]
- Carta, T.; Razzuoli, E.; Fruscione, F.; Zinellu, S.; Meloni, D.; Anfossi, A.; Chessa, B.; Dei, G.S.; Graham, S.P.; Oggiano, A.; et al. Comparative Phenotypic and Functional Analyses of the Effects of IL-10 or TGF-beta on Porcine Macrophages. Animals 2021, 11, 1098. [Google Scholar] [CrossRef]
- Sebastian, R.; Sravanthi, M.; Umapathi, V.; Krishnaswamy, N.; Priyanka, M.; Dechamma, H.J.; Ganesh, K.; Basagoudanavar, S.H.; Sanyal, A.; Reddy, G.R. Foot and mouth disease virus undergoes non-progressive replication in mice peritoneal macrophages and induces M1 polarization. Virus Res. 2020, 281, 197906. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Liu, D.; Chen, J.; Gan, F.; Huang, K. Low-Level Aflatoxin B1 Promotes Influenza Infection and Modulates a Switch in Macrophage Polarization from M1 to M2. Cell. Physiol. Biochem. 2018, 49, 1151–1167. [Google Scholar] [CrossRef]
- Xia, C.; Xu, W.; Ai, X.; Zhu, Y.; Geng, P.; Niu, Y.; Zhu, H.; Zhou, W.; Huang, H.; Shi, X. Autophagy and Exosome Coordinately Enhance Macrophage M1 Polarization and Recruitment in Influenza A Virus Infection. Front. Immunol. 2022, 13, 2053. [Google Scholar] [CrossRef]
- Xiang, X.; Wu, Y.; Lv, X.; Xu, R.; Liu, Y.; Pan, S.; He, M.; Lai, G. Hepatitis B Virus Infection Promotes M2 Polarization of Macrophages by Upregulating the Expression of B7x In Vivo and In Vitro. Viral Immunol. 2022, 35, 597–608. [Google Scholar] [CrossRef]
- Moyano, A.; Ferressini Gerpe, N.M.; De Matteo, E.; Preciado, M.V.; Chabay, P.; Jung, J.U.; Jung, J.U. M1 Macrophage Polarization Prevails in Epstein-Barr Virus-Infected Children in an Immunoregulatory Environment. J. Virol. 2022, 96, e143421. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Yu, Z.; Chen, Z.; Zhang, R.; Ding, X.; Yang, X.; Niu, X.; Li, M.; Zhang, L.; Billiar, T.R.; et al. The HIV protease inhibitor Saquinavir attenuates sepsis-induced acute lung injury and promotes M2 macrophage polarization via targeting matrix metalloproteinase-9. Cell Death Dis. 2021, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Jhan, M.; Chen, C.; Shen, T.; Tseng, P.; Wang, Y.; Satria, R.D.; Yu, C.; Lin, C. Polarization of Type 1 Macrophages Is Associated with the Severity of Viral Encephalitis Caused by Japanese Encephalitis Virus and Dengue Virus. Cells 2021, 10, 3181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, Y.; Zhai, N.; Song, H.; Li, H.; Yang, Y.; Li, T.; Guo, X.; Chi, B.; Niu, J.; et al. HCV core protein inhibits polarization and activity of both M1 and M2 macrophages through the TLR2 signaling pathway. Sci. Rep. 2016, 6, 6160. [Google Scholar] [CrossRef]
- Grosche, L.; Knippertz, I.; Konig, C.; Royzman, D.; Wild, A.B.; Zinser, E.; Sticht, H.; Muller, Y.A.; Steinkasserer, A.; Lechmann, M. The CD83 Molecule-An Important Immune Checkpoint. Front. Immunol. 2020, 11, 721. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, Q.R.; Mi, Y.; Liu, J.Q.; Liu, Z.Q.; Wang, S.; Liu, Z.G.; Yang, P.C.; Zheng, P.Y. Intestinal Epithelial Cell-Derived CD83 Contributes to Regulatory T-Cell Generation and Inhibition of Food Allergy. J. Innate Immun. 2021, 13, 295–305. [Google Scholar] [CrossRef]
- Xiong, L.; Wang, D.; Lin, S.; Wang, Y.; Luo, M.; Gao, L. Soluble CD83 inhibits acute rejection by up regulating TGF-beta and IDO secretion in rat liver transplantation. Transpl. Immunol. 2021, 64, 101351. [Google Scholar] [CrossRef]
- Huo, S.; Zhang, J.; Liang, S.; Wu, F.; Zuo, Y.; Cui, D.; Zhang, Y.; Zhong, Z.; Zhong, F. Membrane-bound and soluble porcine CD83 functions antithetically in T cell activation and dendritic cell differentiation in vitro. Dev. Comp. Immunol. 2019, 99, 103398. [Google Scholar] [CrossRef]
- Doebbeler, M.; Koenig, C.; Krzyzak, L.; Seitz, C.; Wild, A.; Ulas, T.; Bassler, K.; Kopelyanskiy, D.; Butterhof, A.; Kuhnt, C.; et al. CD83 expression is essential for Treg cell differentiation and stability. JCI Insight 2018, 3, 9712. [Google Scholar] [CrossRef]
- Wu, Y.J.; Song, Y.N.; Geng, X.R.; Ma, F.; Mo, L.H.; Zhang, X.W.; Liu, D.B.; Liu, Z.G.; Yang, P.C. Soluble CD83 alleviates experimental allergic rhinitis through modulating antigen-specific Th2 cell property. Int. J. Biol. Sci. 2020, 16, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Liedtke, K.; Alter, C.; Gunther, A.; Hovelmeyer, N.; Klopfleisch, R.; Naumann, R.; Wunderlich, F.T.; Buer, J.; Westendorf, A.M.; Hansen, W. Endogenous CD83 Expression in CD4(+) Conventional T Cells Controls Inflammatory Immune Responses. J. Immunol. 2020, 204, 3217–3226. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, B.; Walton, K.; Reff, J.; Sagatys, E.M.; Tu, N.; Boucher, J.; Li, G.; Ghafoor, T.; Felices, M.; Miller, J.S.; et al. Human CD83-targeted chimeric antigen receptor T cells prevent and treat graft-versus-host disease. J. Clin. Investig. 2020, 130, 4652–4662. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Mizuguchi, M.; Takahashi, Y.; Fujii, H.; Tanaka, R.; Fukushima, T.; Tomoyose, T.; Ansari, A.A.; Nakamura, M. Human T-cell leukemia virus type-I Tax induces the expression of CD83 on T cells. Retrovirology 2015, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Fan, B.; Bai, J.; Wang, H.; Li, Y.; Jiang, P. The N-N non-covalent domain of the nucleocapsid protein of type 2 porcine reproductive and respiratory syndrome virus enhances induction of IL-10 expression. J. Gen. Virol. 2015, 96, 1276–1286. [Google Scholar] [CrossRef]
- Wang, H.; Du, L.; Liu, F.; Wei, Z.; Gao, L.; Feng, W.H. Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Induces Interleukin-17 Production via Activation of the IRAK1-PI3K-p38MAPK-C/EBPbeta/CREB Pathways. J. Virol. 2019, 93, 19. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, A.; Nemati, M.; Jafarzadeh, S. Contribution of STAT3 to the pathogenesis of COVID-19. Microb. Pathog. 2021, 154, 104836. [Google Scholar] [CrossRef] [PubMed]
- Lugo-Villarino, G.; Verollet, C.; Maridonneau-Parini, I.; Neyrolles, O. Macrophage polarization: Convergence point targeted by mycobacterium tuberculosis and HIV. Front. Immunol. 2011, 2, 43. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.F.; Thomas, P.; Lopez, O.A.; Errasti, A.E.; Charo, N.; Romanowski, V.; Gorgojo, J.; Rodriguez, M.E.; Carrera, S.E.; Gomez, R.M. Junin Virus Triggers Macrophage Activation and Modulates Polarization According to Viral Strain Pathogenicity. Front. Immunol. 2019, 10, 2499. [Google Scholar] [CrossRef]
- Ji, Q.; Qu, G.; Liu, B.; Bai, Y.; Wang, G.; Chen, R.; Zheng, X.; Zhang, Z.; Yang, Y.; Wu, C. Evaluation of porcine GM-CSF during PRRSV infection in vitro and in vivo indicating a protective role of GM-CSF related with M1 biased activation in alveolar macrophage during PRRSV infection. Front. Immunol. 2022, 13, 7338. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, S.; Cui, Z.; Li, W.; Xu, P.; Liu, H.; Miao, X.; Chen, Y.; Han, F.; Zhang, H.; et al. MicroRNA-376b-3p Promotes Porcine Reproductive and Respiratory Syndrome Virus Replication by Targeting Viral Restriction Factor TRIM22. J. Virol. 2022, 96, e159721. [Google Scholar] [CrossRef]
- De Baere, M.I.; Van Gorp, H.; Delputte, P.L.; Nauwynck, H.J. Interaction of the European genotype porcine reproductive and respiratory syndrome virus (PRRSV) with sialoadhesin (CD169/Siglec-1) inhibits alveolar macrophage phagocytosis. Vet. Res. 2012, 43, 47. [Google Scholar] [CrossRef]
- Mo, L.H.; Luo, X.Q.; Yang, G.; Liu, J.Q.; Yang, L.T.; Liu, Z.Q.; Wang, S.; Liu, D.B.; Liu, Z.G.; Yang, P.C. Epithelial cell-derived CD83 restores immune tolerance in the airway mucosa by inducing regulatory T-cell differentiation. Immunology 2021, 163, 310–322. [Google Scholar] [CrossRef]
- Horvatinovich, J.M.; Grogan, E.W.; Norris, M.; Steinkasserer, A.; Lemos, H.; Mellor, A.L.; Tcherepanova, I.Y.; Nicolette, C.A.; Debenedette, M.A. Soluble CD83 Inhibits T Cell Activation by Binding to the TLR4/MD-2 Complex on CD14(+) Monocytes. J. Immunol. 2017, 198, 2286–2301. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Liang, S.; Zhong, Z.; Wen, J.; Li, W.; Wang, L.; Xu, J.; Zhong, F.; Li, X. Soluble CD83 inhibits human monocyte differentiation into dendritic cells in vitro. Cell. Immunol. 2014, 292, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Peckert-Maier, K.; Schonberg, A.; Wild, A.B.; Royzman, D.; Braun, G.; Stich, L.; Hadrian, K.; Tripal, P.; Cursiefen, C.; Steinkasserer, A.; et al. Pre-incubation of corneal donor tissue with sCD83 improves graft survival via the induction of alternatively activated macrophages and tolerogenic dendritic cells. Am. J. Transplant. 2022, 22, 438–454. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) |
|---|---|
| GAPDH-F | CCTTCCGTGTCCCTACTGCCAAC |
| GAPDH-R | GACGCCTGCTTCACCACCTTCT |
| TNF-α-F | ACTCGGAACCTCATGGACAG |
| TNF-α-R | AGGGGTGAGTCAGTGTGACC |
| IL-10-F | ACTTTAAGGGTTACCTGGGTTT |
| IL-10-R | TGCTTCACTTTTGCATCTTC |
| iNOS-F | GCACCTGCGTTATGCCACCAAC |
| iNOS-R | TGAGCTGAGCGTTCCAGACCC |
| Arg-1-F | AGCCCGTGTCAACATGACTTCC |
| Arg-1-R | TTGTGTTGGCATCTTTACTGA |
| sCD83-F | CGCTCTCCAGAATGGCTCTT |
| sCD83-R | ACTCTGCTGTCGTGCAAACT |
| Primer | Sequence (5′-3′) a |
|---|---|
| A-BB0907-Fwd | GCGTTAATTAAACCGTCATGACGTATAGGTGTTG |
| A-BB0907-Rev | TGTCTCGAGAATCATCTTTGGGAGAAACC |
| B-BB0907-Fwd | TTCTTAATTAAATGATTCTCGAGACACCGCC |
| B-BB0907-Rev | GTGCTTAAGTTCATTACCACCTGTAACGGAT |
| C1-BB0907-Fwd | GCGTTAATTAAAATGAACTTAAGCACCTATGCC |
| C1-BB0907-Rev | TTGACACAGAGGTAATCGGGTCGCCAGAC |
| C2-BB0907-Fwd | GTCTGGCGACCCGATTACCTCTGTGTCAA |
| C2-BB0907-Rev | CGGGGGAAAATGAAACCTCATGCTGGT |
| D1-BB0907-Fwd | TCGTTAATTAAGTTTCGGGCGCGCCAGAAAGGG |
| D-BB0907-Rev(SwaI) | TTCGGCTTGGGATTTAAATATGCATTTTTTTTTTTTTTTTTTTTT |
| D-BB0907-Rev(SpeI) | CTCACTAGTAACGGCCGCCAGTGTGCTGGAATTCGGCTTGGGATTT |
| XhoI-BB0907-Fwd | GGTACCATGGCCAAACTCGAGGCTTTTGCCGATACC |
| XhoI-BB0907-Rev | GGTATCGGCAAAAGCCTCGAGTTTGGCCATGGTACC |
| Primer | Sequence (5′-3′) a |
|---|---|
| 43ASF | GGCAAGGGACCGGGGGCGAAAAATAGGAAGACA |
| 43ASR | TGTCTTCCTATTTTTCGCCCCCGGTCCCTTGCC |
| 44ASF | AAGGGACCGGGGAAGGCAAATAGGAAGACAAAA |
| 44ASR | TTTTGTCTTCCTATTTGCCTTCCCCGGTCCCTT |
| P192-5ASF | TGCCGGTTTAACGTTGCAGCAGCTGCAGCGCTGCAATTCCCTGCC |
| P192-5ASR | GGCAGGGAATTGCAGCGCTGCAGCTGCTGCAACGTTAAACCGGCA |
| L212-5ASF | CCATGGGTTCGCATCGCGGCCGCCGCTGCGTGTCCTGGCAAGAAC |
| L212-5ASR | GTTCTTGCCAGGACACGCAGCGGCGGCCGCGATGCGAACCCATGG |
| L5ASF | GATAAGTCTGGGATAGCTGATCGGTGCACGTGT |
| L5ASR | ACACGTGCACCGATCAGCTATCCCAGACTTATC |
| D6ASF | AAGTCTGGGATACTTGCTCGGTGCACGTGTACC |
| D6ASR | GGTACACGTGCACCGAGCAAGTATCCCAGACTT |
| G45ASF | CAAGTTCCTGAGCTTGCGGTGCTGGGTCTATT |
| G45ASR | AAATAGACCCAGCACCGCAGCTCAGGAACTTG |
| G48ASF | GAGCTTGGGGTGCTGGCTCTATTTTATAGGCCC |
| G48ASR | GGGCCTATAAAATAGAGCCAGCACCCCAAGCTC |
| L61ASF | CCACTCCGGTGGACGGCGCCACGTGCATTCCCC |
| L61ASR | GGGGAATGCACGTGGCGCCGTCCACCGGAGTGG |
| P62ASF | CTCCGGTGGACGTTGGCACGTGCATTCCCCACT |
| P62ASR | AGTGGGGAATGCACGTGCCAACGTCCACCGGAG |
| R63ASF | CGGTGGACGTTGCCAGCTGCATTCCCCACTGTC |
| R63ASR | GACAGTGGGGAATGCAGCTGGCAACGTCCACCG |
| F65ASF | ACGTTGCCACGTGCAGCCCCCACTGTCGAGTGC |
| F65ASR | GCACTCGACAGTGGGGGCTGCACGTGGCAACGT |
| P66ASF | TTGCCACGTGCATTCGCCACTGTCGAGTGCTCC |
| P66ASR | GGAGCACTCGACAGTGGCGAATGCACGTGGCAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Ma, T.; Zhang, Q.; Wang, Y.; Song, C.; Lai, M.; Zhang, C.; Fang, X.; Chen, X. Porcine Reproductive and Respiratory Syndrome Virus Modulates the Switch of Macrophage Polarization from M1 to M2 by Upregulating MoDC-Released sCD83. Viruses 2023, 15, 773. https://doi.org/10.3390/v15030773
Gong X, Ma T, Zhang Q, Wang Y, Song C, Lai M, Zhang C, Fang X, Chen X. Porcine Reproductive and Respiratory Syndrome Virus Modulates the Switch of Macrophage Polarization from M1 to M2 by Upregulating MoDC-Released sCD83. Viruses. 2023; 15(3):773. https://doi.org/10.3390/v15030773
Chicago/Turabian StyleGong, Xingyu, Tianyi Ma, Qiaoya Zhang, Yanhong Wang, Chengchuang Song, Min Lai, Chunlei Zhang, Xingtang Fang, and Xi Chen. 2023. "Porcine Reproductive and Respiratory Syndrome Virus Modulates the Switch of Macrophage Polarization from M1 to M2 by Upregulating MoDC-Released sCD83" Viruses 15, no. 3: 773. https://doi.org/10.3390/v15030773