
Early Response of CD8+ T Cells in COVID-19 Patients

 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population, Setting, and Data Collection
2.2. Real Time PCR
2.3. Lymphocyte Subpopulation Test
2.4. Immunophenotyping
2.5. Statistical Analysis
3. Results
3.1. Patients’ Baseline Characteristics
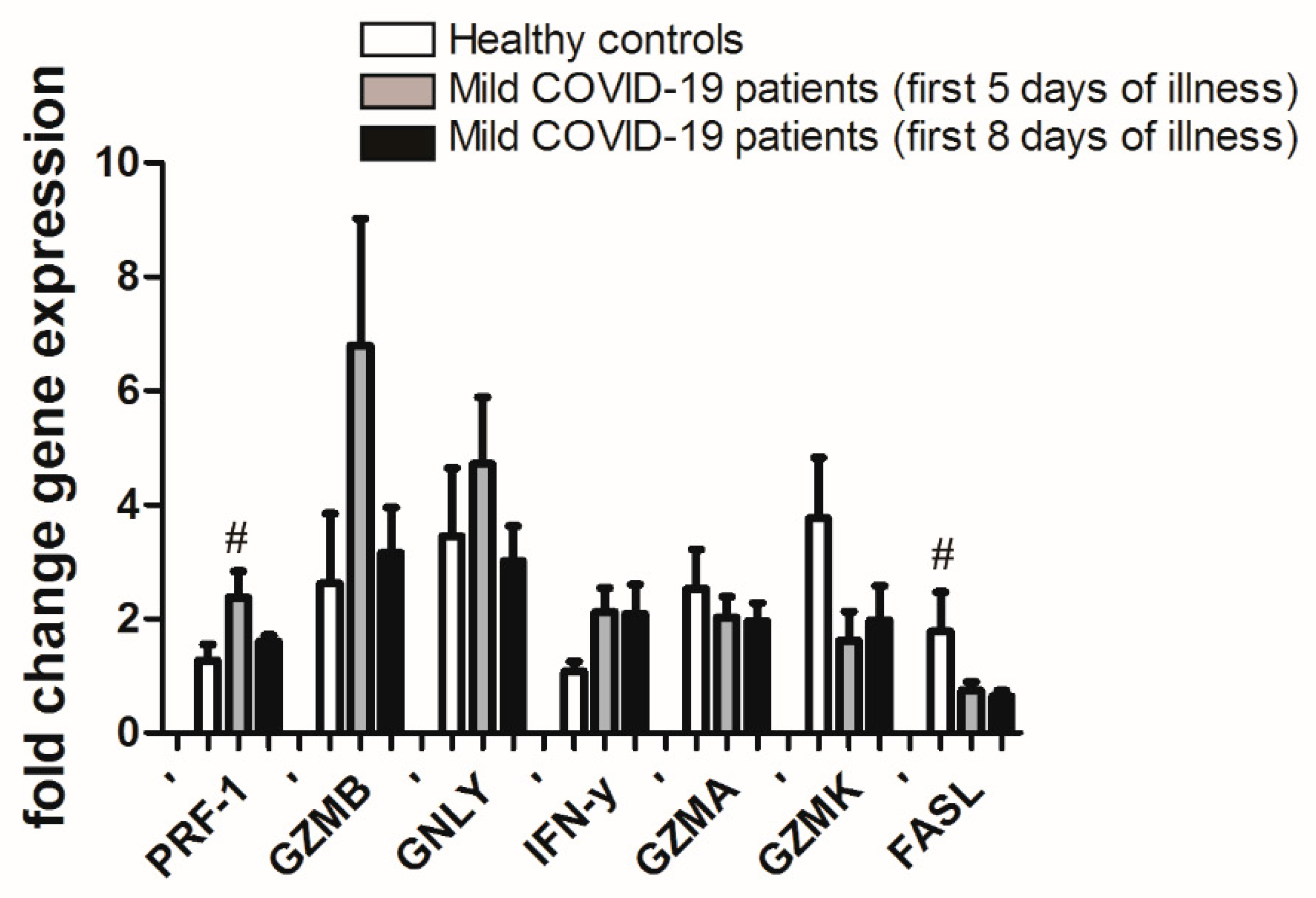
3.2. Gene Expression in CD8+ T Cells from PBL of Mild COVID-19 Patients in the First Week of Illness Compared to Healthy Controls
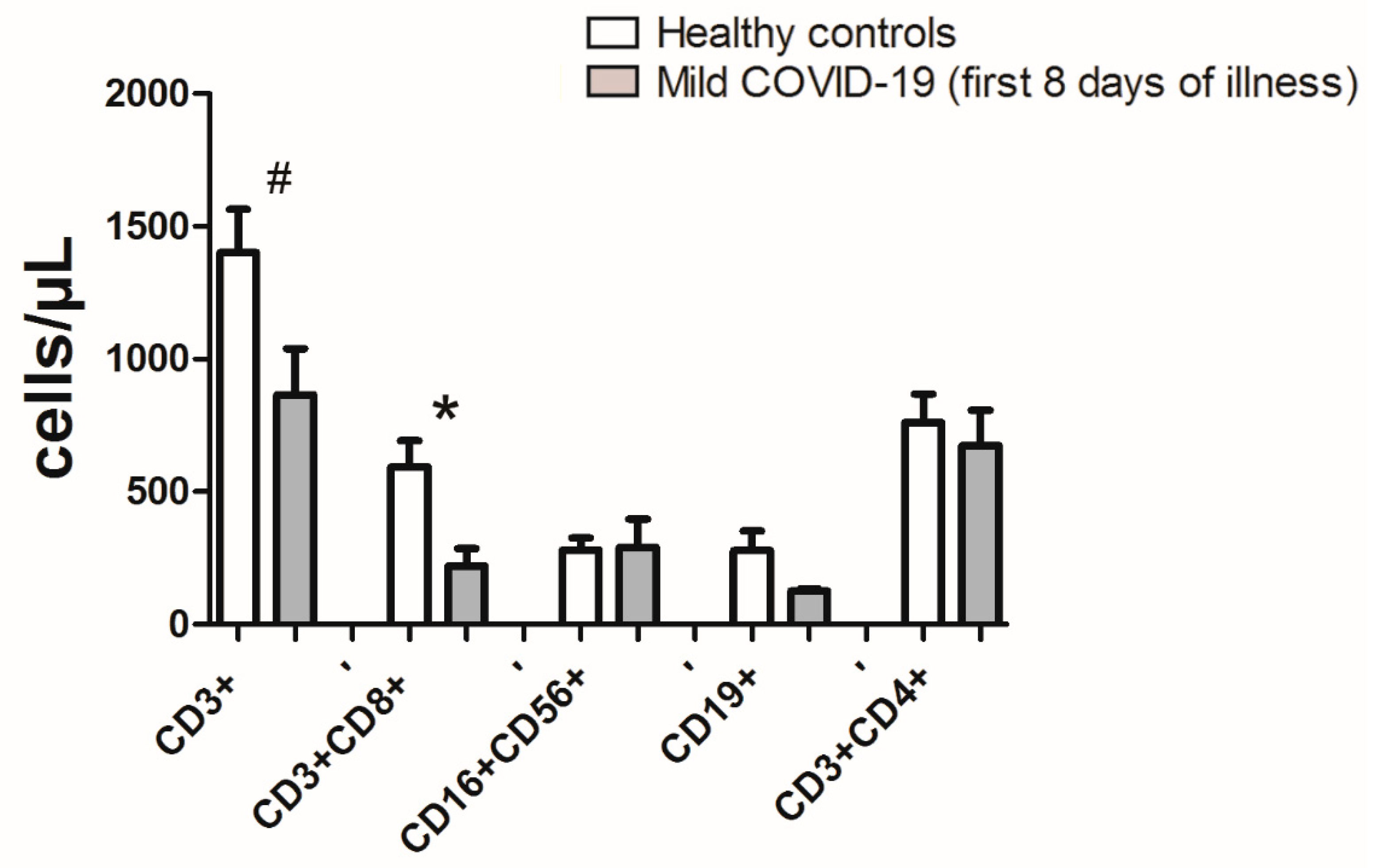
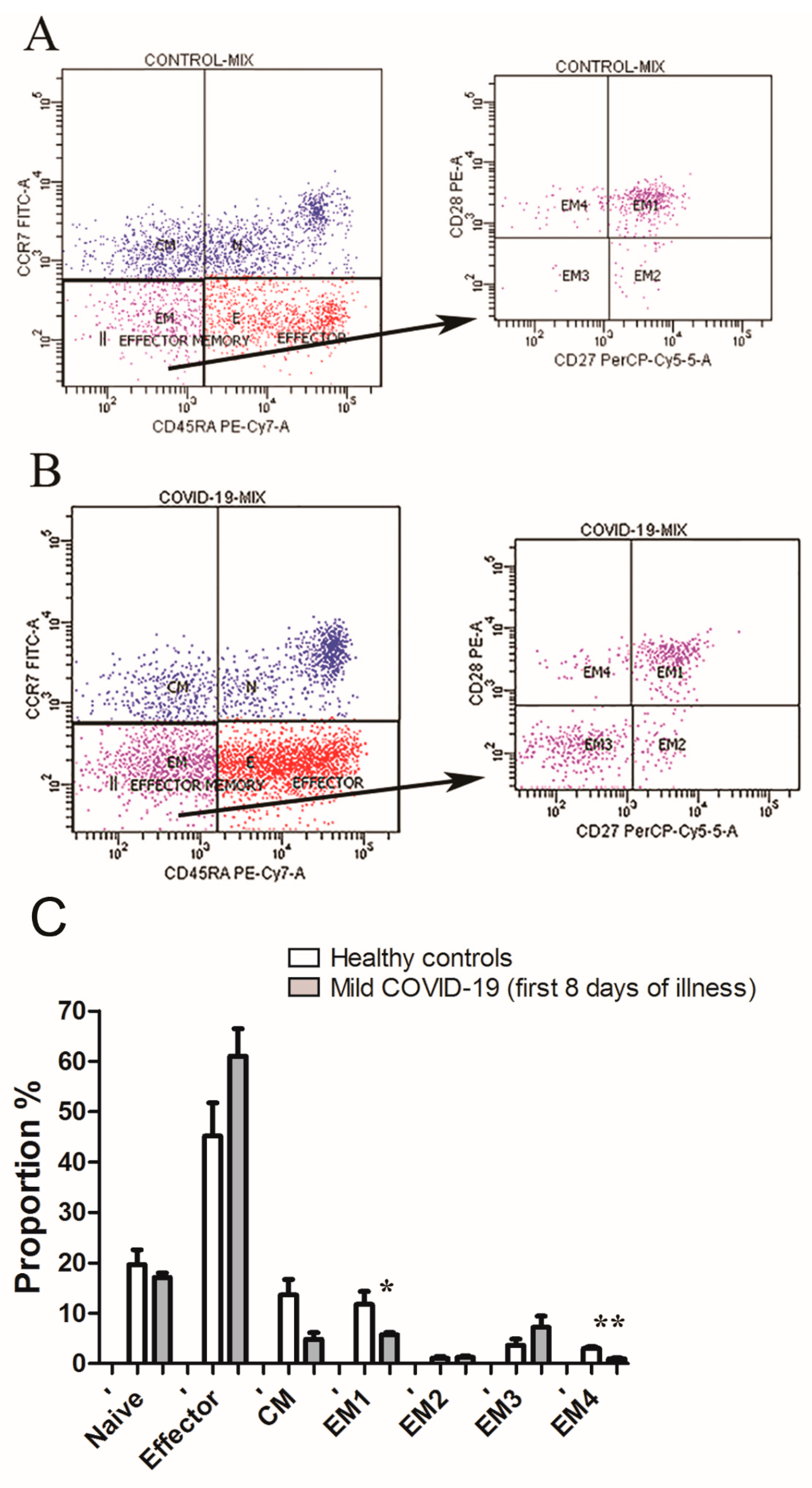
3.3. Lymphopenia with Relative Decreases of CD8+ Effector Memory Subsets Define First Week of COVID-19 Infection
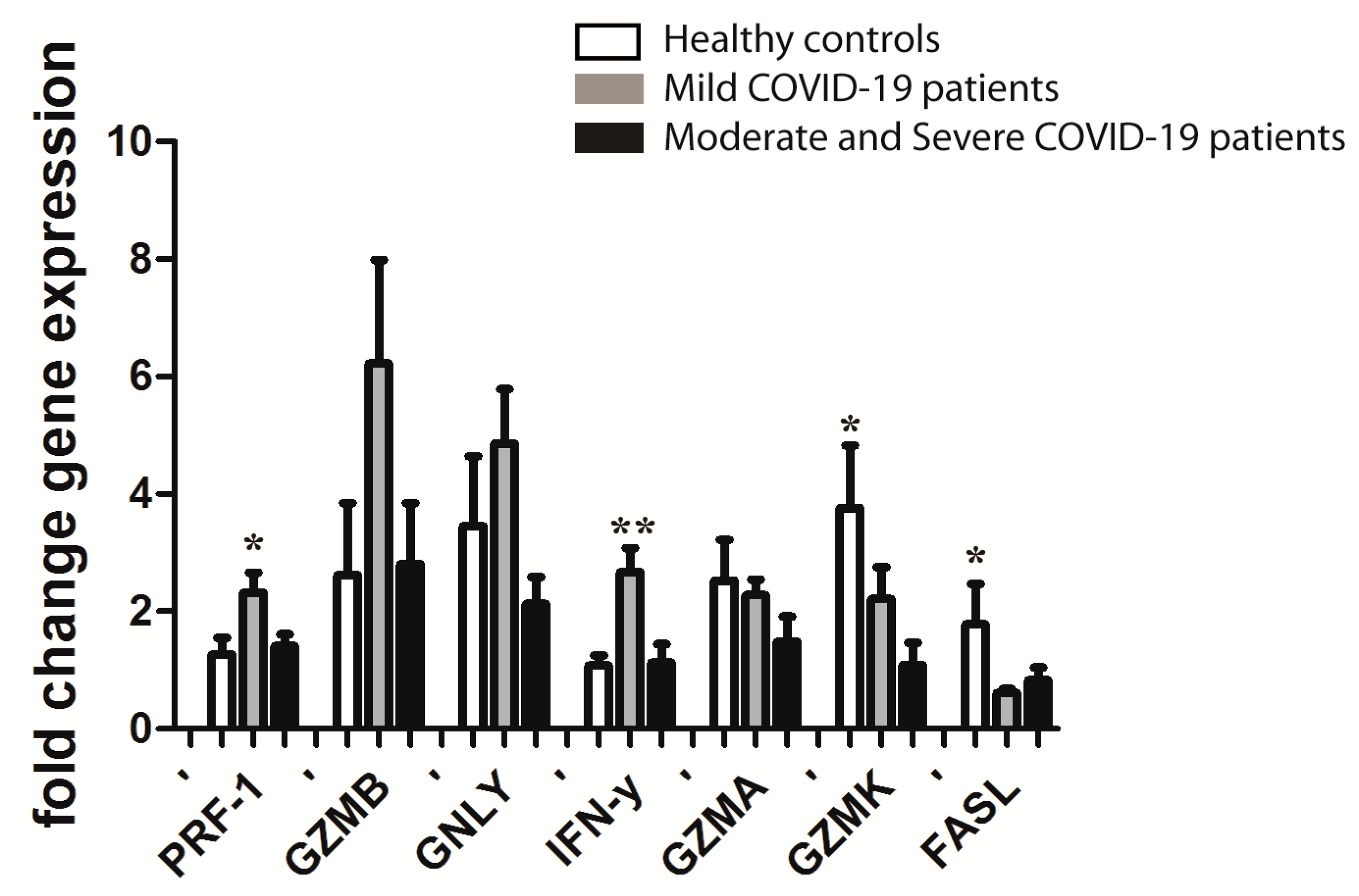
3.4. Gene Expression in CD8+ T Cells from PBL of COVID-19 Patients in the First Eight Days of Disease, Depending on the Severity of Disease after Ten Days (Mild versus Moderate and Severe) Compared to Healthy Controls
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Weekly Epidemiological Update on COVID-19—26 October 2021. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---26-october-2021 (accessed on 1 November 2021).
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Vabret, N.; Britton, G.J.; Gruber, C.; Hegde, S.; Kim, J.; Kuksin, M.; Levantovsky, R.; Malle, L.; Moreira, A.; Park, M.D.; et al. Immunology of COVID-19: Current State of the Science. Immunity 2020, 52, 910–941. [Google Scholar] [CrossRef]
- Fathi, N.; Rezaei, N. Lymphopenia in COVID-19: Therapeutic opportunities. Cell Biol. Int. 2020, 44, 1792–1797. [Google Scholar] [CrossRef]
- Ganji, A.; Farahani, I.; Khansarinejad, B.; Ghazavi, A.; Mosayebi, G. Increased expression of CD8 marker on T-cells in COVID-19 patients. Blood Cells Mol. Dis. 2020, 83, 102437. [Google Scholar] [CrossRef] [PubMed]
- DiPiazza, A.T.; Graham, B.S.; Ruckwardt, T.J. T cell immunity to SARS-CoV-2 following natural infection and vaccination. Biochem. Biophys. Res. Commun. 2021, 538, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Long, W.; Tu, M.; Chen, S.; Huang, Y.; Wang, S.; Zhou, W.; Chen, D.; Zhou, L.; Wang, M.; et al. Lymphocyte subset (CD4+, CD8+) counts reflect the severity of infection and predict the clinical outcomes in patients with COVID-19. J. Infect. 2020, 81, 318–356. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.; Moulana, Z.; Sepidarkish, M.; Bagherzadeh, M.; Rezanejad, M.; Mirzakhani, M.; Jafari, M.; Mohammadnia-Afrouzi, M. Pronounce expression of Tim-3 and CD39 but not PD1 defines CD8 T cells in critical COVID-19 patients. Microb. Pathog. 2021, 153, 104779. [Google Scholar] [CrossRef]
- Belaid, B.; Mahammad, L.L.; Mihi, B.; Rahali, S.Y.; Djidjeli, A.; Larab, Z.; Berkani, L.; Berkane, I.; Sayah, W.; Merah, F.; et al. T cell counts and IL-6 concentration in blood of North African COVID-19 patients are two independent prognostic factors for severe disease and death. J. Leukoc. Biol. 2021. online ahead of print. [Google Scholar] [CrossRef]
- Peng, X.; Ouyang, J.; Isnard, S.; Lin, J.; Fombuena, B.; Zhu, B.; Routy, J.-P. Sharing CD4+ T Cell Loss: When COVID-19 and HIV Collide on Immune System. Front. Immunol. 2020, 11, 596631. [Google Scholar] [CrossRef]
- Hamann, D.; Roos, M.T.; van Lier, R.A. Faces and phases of human CD8 T-cell development. Immunol. Today 1999, 20, 177–180. [Google Scholar] [CrossRef]
- Pennock, N.D.; White, J.T.; Cross, E.W.; Cheney, E.E.; Tamburini, B.A.; Kedl, R.M. T cell responses: Naïve to memory and everything in between. Adv. Physiol. Educ. 2013, 37, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef]
- Basic Immunology—6th Edition. Available online: https://www.elsevier.com/books/basic-immunology/abbas/978-0-323-54943-1 (accessed on 26 September 2021).
- Nagasawa, M.; Ogawa, K.; Nagata, K.; Shimizu, N. Serum granulysin as a possible biomarker of natural killer cell neoplasms. Br. J. Haematol. 2010, 148, 812–814. [Google Scholar] [CrossRef]
- Bade, B.; Boettcher, H.E.; Lohrmann, J.; Hink-Schauer, C.; Bratke, K.; Jenne, D.E.; Virchow, J.C., Jr.; Luttmann, W. Differential expression of the granzymes A, K and M and perforin in human peripheral blood lymphocytes. Int. Immunol. 2005, 17, 1419–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, M.; Tian, L.; Luo, G.; Yu, X. Interferon-Gamma-Mediated Osteoimmunology. Front. Immunol. 2018, 9, 1508. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, A.; Dixit, V.M. Death receptors: Signaling and modulation. Science 1998, 281, 1305–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hersperger, A.R.; Pereyra, F.; Nason, M.; Demers, K.; Sheth, P.; Shin, L.Y.; Kovacs, C.M.; Rodriguez, B.; Sieg, S.F.; Teixeira-Johnson, L.; et al. Perforin Expression Directly Ex Vivo by HIV-Specific CD8+ T-Cells Is a Correlate of HIV Elite Control. PLoS Pathog. 2010, 6, e1000917. [Google Scholar] [CrossRef]
- Almeida, J.R.; Price, D.A.; Papagno, L.; Arkoub, Z.A.; Sauce, D.; Bornstein, E.; Asher, T.E.; Samri, A.; Schnuriger, A.; Theodorou, I.; et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J. Exp. Med. 2007, 204, 2473–2485. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Zhang, M.; Zhu, T.; Zhili, N.; Liu, Z.; Xiang, R.; Zhang, W.; Xu, Y. Dynamic changes in peripheral blood lymphocyte subsets in adult patients with COVID-19. Int. J. Infect. Dis. 2020, 98, 353–358. [Google Scholar] [CrossRef]
- Information on COVID-19 Treatment, Prevention and Research. COVID-19 Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov/ (accessed on 26 September 2021).
- El Zein, S.; Chehab, O.; Kanj, A.; Akrawe, S.; Alkassis, S.; Mishra, T.; Shatta, M.; El-Hor, N.; Salimnia, H.; Chandrasekar, P. SARS-CoV-2 infection: Initial viral load (iVL) predicts severity of illness/outcome, and declining trend of iVL in hospitalized patients corresponds with slowing of the pandemic. PLoS ONE 2021, 16, e0255981. [Google Scholar] [CrossRef] [PubMed]
- García, L.F. Immune Response, Inflammation, and the Clinical Spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef] [PubMed]
- Ponti, G.; Maccaferri, M.; Ruini, C.; Tomasi, A.; Ozben, T. Biomarkers associated with COVID-19 disease progression. Crit. Rev. Clin. Lab. Sci. 2020, 57, 389–399. [Google Scholar] [CrossRef]
- Samprathi, M.; Jayashree, M. Biomarkers in COVID-19: An Up-To-Date Review. Front. Pediatr. 2021, 8, 607647. [Google Scholar] [CrossRef] [PubMed]
- Velavan, T.P.; Meyer, C.G. Mild versus severe COVID-19: Laboratory markers. Int. J. Infect. Dis. 2020, 95, 304–307. [Google Scholar] [CrossRef]
- Ahmadi, P.; Hartjen, P.; Kohsar, M.; Kummer, S.; Schmiedel, S.; Bockmann, J.-H.; Fathi, A.; Huber, S.; Haag, F.; Wiesch, J.S.Z. Defining the CD39/CD73 Axis in SARS-CoV-2 Infection: The CD73- Phenotype Identifies Polyfunctional Cytotoxic Lymphocytes. Cells 2020, 9, 1750. [Google Scholar] [CrossRef]
- Mazzoni, A.; Salvati, L.; Maggi, L.; Capone, M.; Vanni, A.; Spinicci, M.; Mencarini, J.; Caporale, R.; Peruzzi, B.; Antonelli, A.; et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J. Clin. Investig. 2020, 130, 4694–4703. [Google Scholar] [CrossRef]
- Bellesi, S.; Metafuni, E.; Hohaus, S.; Maiolo, E.; Marchionni, F.; D’Innocenzo, S.; la Sorda, M.; Ferraironi, M.; Ramundo, F.; Fantoni, M.; et al. Increased CD95 (Fas) and PD-1 expression in peripheral blood T lymphocytes in COVID-19 patients. Br. J. Haematol. 2020, 191, 207–211. [Google Scholar] [CrossRef]
- Li, M.; Guo, W.; Dong, Y.; Wang, X.; Dai, D.; Liu, X.; Wu, Y.; Li, M.; Zhang, W.; Zhou, H.; et al. Elevated Exhaustion Levels of NK and CD8+ T Cells as Indicators for Progression and Prognosis of COVID-19 Disease. Front. Immunol. 2020, 11, 580237. [Google Scholar] [CrossRef]
- Kang, C.K.; Han, G.C.; Kim, M.; Kim, G.; Shin, H.M.; Song, K.-H.; Choe, P.G.; Beom Park, W.; Kim, E.S.; Kim, H.B.; et al. Aberrant hyperactivation of cytotoxic T-cell as a potential determinant of COVID-19 severity. Int. J. Infect. Dis. 2020, 97, 313–321. [Google Scholar] [CrossRef]
- Jiang, Y.; Wei, X.; Guan, J.; Qin, S.; Wang, Z.; Lu, H.; Qian, J.; Wu, L.; Chen, Y.; Chen, Y.; et al. COVID-19 pneumonia: CD8+ T and NK cells are decreased in number but compensatory increased in cytotoxic potential. Clin. Immunol. 2020, 218, 108516. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhang, C.; Huang, F.; Yang, Y.; Wang, F.; Yuan, J.; Zhang, Z.; Qin, Y.; Li, X.; Zhao, D.; et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Natl. Sci. Rev. 2020, 7, 1003–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westmeier, J.; Paniskaki, K.; Karaköse, Z.; Werner, T.; Sutter, K.; Dolff, S.; Overbeck, M.; Limmer, A.; Liu, J.; Zheng, X.; et al. Impaired Cytotoxic CD8+ T Cell Response in Elderly COVID-19 Patients. mBio 2020, 11, e02243-20. [Google Scholar] [CrossRef]
- Tavukcuoglu, E.; Horzum, U.; Inkaya, A.C.; Unal, S.; Esendagli, G. Functional responsiveness of memory T cells from COVID-19 patients. Cell Immunol. 2021, 365, 104363. [Google Scholar] [CrossRef]
- Kratzer, B.; Trapin, D.; Ettel, P.; Körmöczi, U.; Rottal, A.; Tuppy, F.; Feichter, M.; Gattinger, P.; Borochova, K.; Dorofeeva, Y.; et al. Immunological imprint of COVID-19 on human peripheral blood leukocyte populations. Allergy 2021, 76, 751–765. [Google Scholar] [CrossRef]
- Kusnadi, A.; Ramírez-Suástegui, C.; Fajardo, V.; Chee, S.J.; Meckiff, B.J.; Simon, H.; Pelosi, E.; Seumois, G.; Ay, F.; Vijayanand, P.; et al. Severely ill COVID-19 patients display impaired exhaustion features in SARS-CoV-2-reactive CD8+ T cells. Sci. Immunol. 2021, 6, eabe4782. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 n = 16 | Healthy Control n = 9 | p Value | |
|---|---|---|---|
| Age (years) (mean ± SD) | 51 ± 13 | 45 ± 10 | 0.678 |
| Gender (M/F) (n, %) | 7 (44%)/9 (56%) | 4 (44%)/5 (56%) | 0.563 |
| Comorbidities (n, %) | 7 (44%) | 2 (22%) | 0.040 |
| Thrombocytes(×109/L) (mean ± SD) | 191 ± 71 | 255 ± 62 | 0.062 |
| Leukocytes (×109/L) (mean ± SD) | 4.3 (3.8−4.9) | 5.9 ± 1.7 | 0.076 |
| Neutrophils (×109/L) (mean ± SD) | 2.4 (1.6−3.3) | 3.32 ± 1.16 | 0.285 |
| Lymphocytes (×109/L) (mean± SD) | 1.3 ± 0.5 | 1.9 ± 0.5 | 0.007 |
| Neutrophils/Lymphocytes (median; IQR) | 1.7 (1.1−3.1) | 1.7 (1.5–1.9) | 0.664 |
| Leukocytes/Lymphocytes (mean ± SD) | 3.2 (2.6−4.8) | 3.1 ± 0.6 | 0.462 |
| Urea (mmol/L) (median; IQR) | 4.2 (3.7−4.6) | 5.0 (4.8–6.0) | 0.015 |
| AST (U/L) (mean ± SD) | 29 (26−39) | 20 ± 7 | 0.016 |
| ALT (U/L) (mean ± SD) | 25 (19−46) | 22 ± 10 | 0.151 |
| LDH (U/L) (median; IQR) | 226 ± 73 | 151 (144–168) | 0.005 |
| CRP (mg/L) (median; IQR) | 9.9 (3.7−45.2) | 1.1 (0.6–1.3) | <0.001 |
| IL6 (pg/mL) (median; IQR) | 5.9 (2.1−35.9) | 1.5 (1.5–1.5) | <0.001 |
| Ferritin (µg/L) (mean ± SD) | 129.0 (86.0−245.0) | 122.5 ± 176.6 | 0.204 |
| d-dimer (mg/L) (mean ± SD) | 0.52 ± 0.27 | 0.47 ± 0.16 | 0.626 |
| PT (ng/mL) (median; IQR) | 0.04 (0.02−0.06) | 0.01 (0.01–0.02) | 0.003 |
| Post Hoc Tests p Value | |||||||
|---|---|---|---|---|---|---|---|
| Control (n = 9) | Mild COVID-19 (n = 9) | MS COVID-19 (n = 7) | p Value | Mild vs. MS | Mild vs. Control | MS vs. Control | |
| Age (years) (mean ± SD) | 45 ± 10 | 46 ± 12 | 58 ± 12 | 0.073 | |||
| Gender (M/F) (n, %) | 5 (56%)/4 (44%) | 3 (33%)/6 (67%) | 4(57%)/3 (43%) | 0.558 | |||
| Comorbidities (n, %) | 2 (22%) | 3 (33%) | 4 (57%) | 0.628 | |||
| Thrombocytes (×109/L) (mean ± SD) | 255 ± 62 | 209 ± 56 | 167 ± 84 | 0.063 | |||
| Leukocytes (×109/L) (mean ± SD) | 5.9 ± 1.7 | 4.1 ± 0.9 | 5.3 ± 2.5 | 0.108 | |||
| Neutrophils (×109/L) (mean ± SD) | 3.32 ± 1.16 | 2.12 ± 0.98 | 3.56 ± 2.21 | 0.133 | |||
| Lymphocytes (×109/L) (mean ± SD) | 1.9 ± 0.5 | 1.4 ± 0.4 | 1.2 ± 0.5 | 0.024 | 0.765 | 0.087 | 0.049 |
| Neutrophils/Lymphocytes (median; IQR) | 1.7 (1.5–1.9) | 1.8 (1.4–2.5) | 3.1 (2.4–3.9) | 0.231 | |||
| Leukocytes/Lymphocytes (mean ± SD) | 3.1 ± 0.6 | 3.2 ± 1.4 | 4.5 ± 1.8 | 0.127 | |||
| Urea (mmol/L) (median; IQR) | 5.0 (4.8–6.0) | 4.2 (4.0–4.4) | 3.7 (3.3–5.7) | 0.066 | |||
| AST (U/L) (mean ± SD) | 20 ± 7 | 26 ± 7 | 44 ± 17 | <0.001 | 0.061 | 0.183 | 0.018 |
| ALT (U/L) (mean ± SD) | 22 ± 10 | 27 ± 13 | 42 ± 27 | 0.083 | |||
| LDH (U/L) (median; IQR) | 151 (144–168) | 156 (145–207) | 299 (284–330) | 0.001 | 0.017 | 1.000 | 0.001 |
| CRP (mg/L) (median; IQR) | 1.1 (0.6−1.3) | 3.8 (3.1−6.0) | 53.7 (16.8–109.0) | <0.001 | 0.126 | 0.049 | <0.001 |
| Ferritin (µg/L) (mean ± SD) | 122.5 ± 176.6 | 122.4 ± 89.3 | 361.7 ± 337.4 | 0.067 | |||
| d-dimer (mg/L) (mean ± SD) | 0.47 ± 0.16 | 0.38 ± 0.22 | 0.71 ± 0.21 | 0.011 | 0.021 | 0.604 | 0.076 |
| PT (ng/mL) (median; IQR) | 0.01 (0.01–0.02 | 0.03 (0.02–0.04) | 0.08 (0.04−0.12) | 0.003 | 0.315 | 0.137 | 0.002 |
| IL6 (pg/mL) (median; IQR) | 1.5 (1.5−1.5) | 2.1 (1.7−4.5) | 50.2 (23.4−56.1) | <0.001 | 0.046 | 0.141 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramljak, D.; Vukoja, M.; Curlin, M.; Vukojevic, K.; Barbaric, M.; Glamoclija, U.; Purisevic, B.; Peric, O.; Soljic, V. Early Response of CD8+ T Cells in COVID-19 Patients. J. Pers. Med. 2021, 11, 1291. https://doi.org/10.3390/jpm11121291
Ramljak D, Vukoja M, Curlin M, Vukojevic K, Barbaric M, Glamoclija U, Purisevic B, Peric O, Soljic V. Early Response of CD8+ T Cells in COVID-19 Patients. Journal of Personalized Medicine. 2021; 11(12):1291. https://doi.org/10.3390/jpm11121291
Chicago/Turabian StyleRamljak, Deni, Martina Vukoja, Marina Curlin, Katarina Vukojevic, Maja Barbaric, Una Glamoclija, Bejana Purisevic, Olivera Peric, and Violeta Soljic. 2021. "Early Response of CD8+ T Cells in COVID-19 Patients" Journal of Personalized Medicine 11, no. 12: 1291. https://doi.org/10.3390/jpm11121291