1. Introduction
The COVID-19 disease is produced by the virus called SARS-COV-2, which emerged in China at the end of 2019. Currently, it has spread worldwide, so the World Health Organization (WHO) has declared a pandemic for which there is no treatment or vaccine [
1].
The process to generate and test a vaccine will take at least a year [
2]. However, by using computational tools such as repositioning, already known drugs (known information on pharmacokinetics, pharmacodynamics and toxicity) and those used for other pathologies can be proposed.
Repositioning drugs shows several advantages; for example, the time and production costs of the drugs are dismissed because the drugs are already available on the market [
3]. In the case of aspirin, this drug was initially used as an analgesic and antipyretic and derived from repositioning strategies; currently, its new indication is as a treatment for colorectal cancer. Another example is Hydroxychloroquine an antiparasitic, which is now used in the treatment of antiarthritic systemic lupus erythematosus [
4]. With regard to the treatment of COVID-19, different molecular targets have been proposed, such as modulation of immune defense, blocking viral cell entry, interfering with the endocytic pathway, targeting the cellular signaling pathway, blocking polyprotein posttranslational processing and various antiviral mechanisms [
5]. SARS-CoV-2 infection can also affect the gastrointestinal tract, liver and pancreatic functions, leading to gastrointestinal symptoms and central and peripheral neurological manifestations, affecting the cardiovascular system, promoting renal dysfunction and, in general, resulting in a multitude of other clinical symptoms [
6].
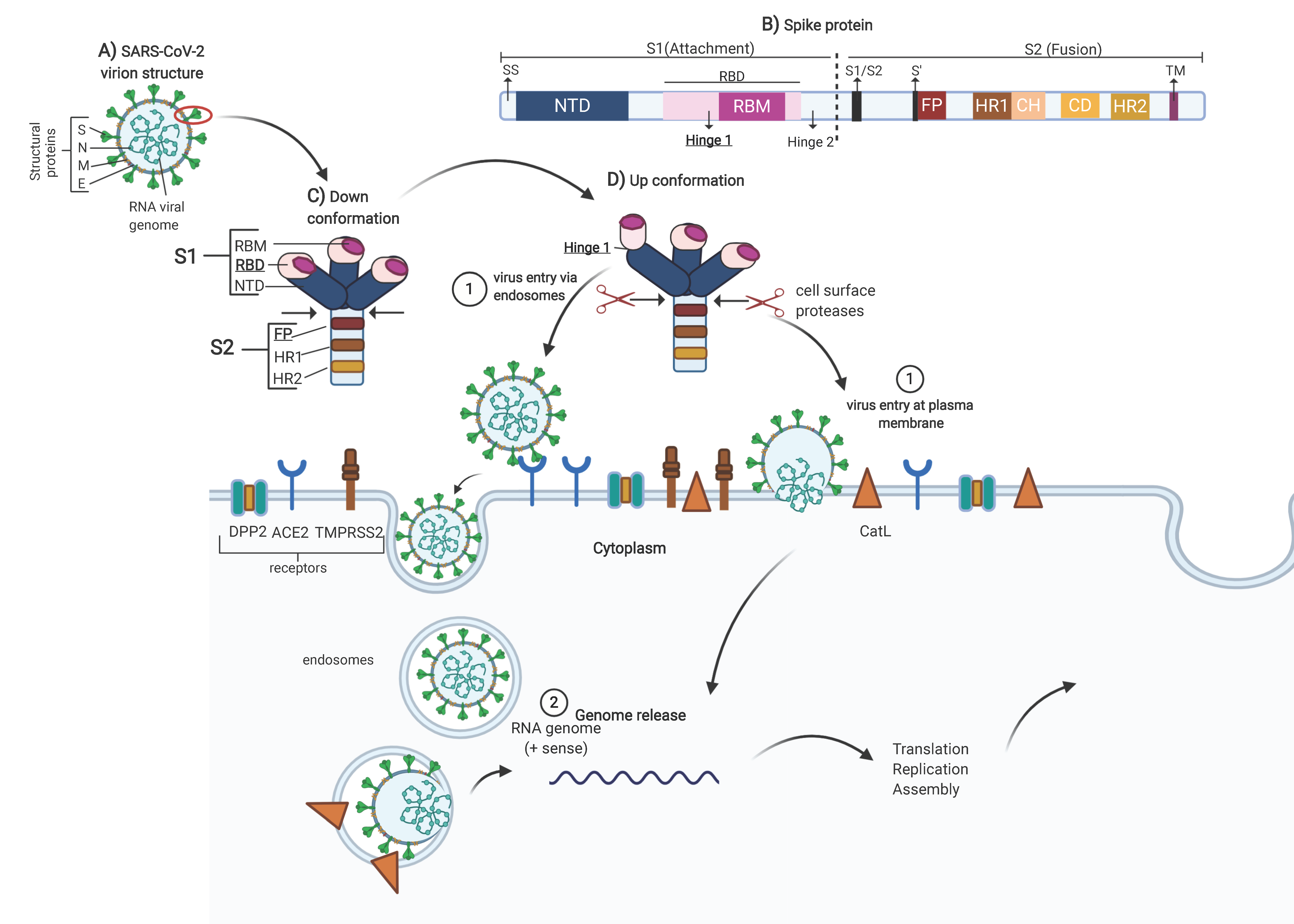
SARS-COV-2 belongs to the coronavirus family, and the genome is +ssRNA, nonsegmented, with a size of 27 to 32 kilobases [
7]. The genome encodes four major structural proteins, spike (S), nucleocapsid (N), membrane (M) and envelope (E), which are required to make complete virus particles (see
Figure 1A) [
7,
8]. The glycoprotein spike allows the penetration of host cells (obligatory step in the virus replication cycle), rendering it a powerful target for drug development.
The spike protein is comprised of the S1 and S2 subunits. The S1 subunit contains a signal peptide (SS), followed by an N-terminal domain (NTD) and receptor-binding domain (RBD). The RBD contains a core structure and a receptor-binding motif (RBM). The S2 subunit contains conserved fusion peptide (FP), heptad repeat 1 (HR1), a central helix (CH), a connector domain (CD), heptad repeat 2 (HR2), a transmembrane domain (TM), and a cytoplasmic tail (CT). At the boundary between S1 and S2 (S1/S2), there is a furin cleavage site at positions 681–684 (see
Figure 1B) [
9,
10].
Conformationally, the spike glycoprotein is organized on the viral surface in homotrimers [
11]. When the RBMs are hidden, the conformation is called down (receptor-inaccessible) (see
Figure 1C). However, the homotrimer is asymmetric because it constantly undergoes structural rearrangement (up conformation) to fuse the viral membrane with the host cell membrane [
12]. When two RBD domains are hidden (receptor-inaccessible), one RBD domain is exposed (receptor-accessible), resulting in the up conformation (see
Figure 1D). This is because the RBD of S1 undergoes hinge-like movements [
9]. In SARS-CoV, two hinge sites were characterized (hinge 1 site (354–361) and hinge 2 site (552–563)), which are responsible for the switch between the up and down conformations [
13]. The RBD in the upper position recognizes the receptor through the receptor-binding motif (RBM), which binds to the outer surface of the claw-like structure of angiotensin-converting enzyme 2 (ACE2). The importance of RBM was demonstrated by comparing the sequences between SARS-CoV and SARS-CoV-2, and the five-amino-acid mutation (Leu455, Phe486, Gln493, Ser494 and Asn501) improves affinity for the receptor. Therefore, it is believed that SARS-CoV-2 is more infectious than SARS-CoV [
10].
After recognition and binding of the RBM by ACE2, the spike glycoprotein needs to be proteolytically activated at the S1/S2 boundary such that S1 dissociates and S2 undergoes a dramatic structural change. These SARS-CoV-2 entry-activating proteases include cell surface type II transmembrane serine proteases (TMPRSS2) and lysosomal protease cathepsins. The structural change that S2 shows when dissociated is the exposure of the fusion peptide (FP) located at amino acid positions 816 to 855. Subsequently, the insertion of the FP into the host membrane is triggered, allowing the fusion of membranes (viral and cellular) to release the viral genome into the cytoplasm [
14].
As mentioned above, the spike glycoprotein is in first contact with the host cell and is crucial for various processes, such as attachment, receptor binding, membrane fusion via conformational changes, internalization of the virus, and host tissue tropism [
8,
10]. Therefore, it is one of the main targets for the development of vaccines and antivirals.
In the search for potential compounds that could be used to treat COVID-19, in this work, we considered employing repositioning techniques, since they represent a new way of approaching drug compounds and targets that have been “derisked” during the development stages. In this work, we focused on drugs approved by the FDA that could target the spike glycoprotein during three specific events (conformational change from down to up, receptor recognition and membrane fusion).
3. Discussion
Spike glycoprotein is the structural protein of the SARS-CoV-2 virus that allows adhesion and binding to the receptor, making it crucial in the first step of infection [
15]. Currently it is considered one of the main targets for vaccine design. However, this protein evades the immune response due to its dynamic structure, for example, when it is found in the down conformation (receptor-inaccessible) where it hides RBD (hidden RBD) and prevents the binding of neutralizing antibodies. Additionally, when the spike glycoprotein is found in the up conformation (accessible receptor), the RBD has high affinity for the receptor (higher affinity than SARS-CoV). In addition, the virus can enter through two routes, receptor-mediated endocytosis and direct fusion to the cell membrane, because the fusion peptide can be exposed by cell surface proteases (depending on the cell type). Therefore, vaccines targeting the membrane fusion S2 subunit can be developed; however, the S2 subunit is less immunogenic than the S1 subunit [
16,
17].
In this work, we employed an in silico strategy called drug repositioning, in which we searched for available drugs and targets of the spike glycoprotein (hinge sites, RBM and FP of interest). From this analysis, it was possible to identify 43 drugs, of which 11 are included in reported clinical trials [
18].
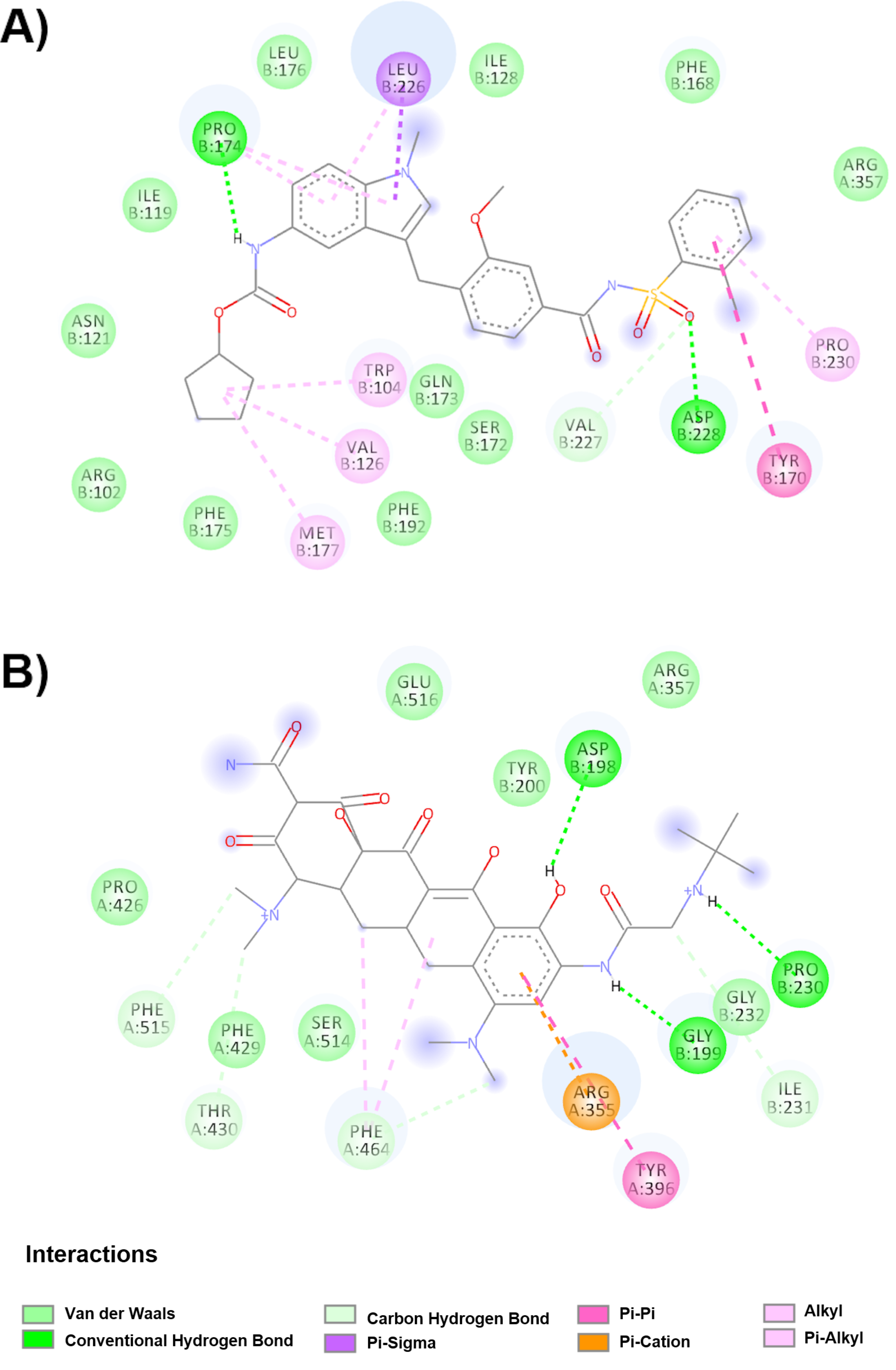
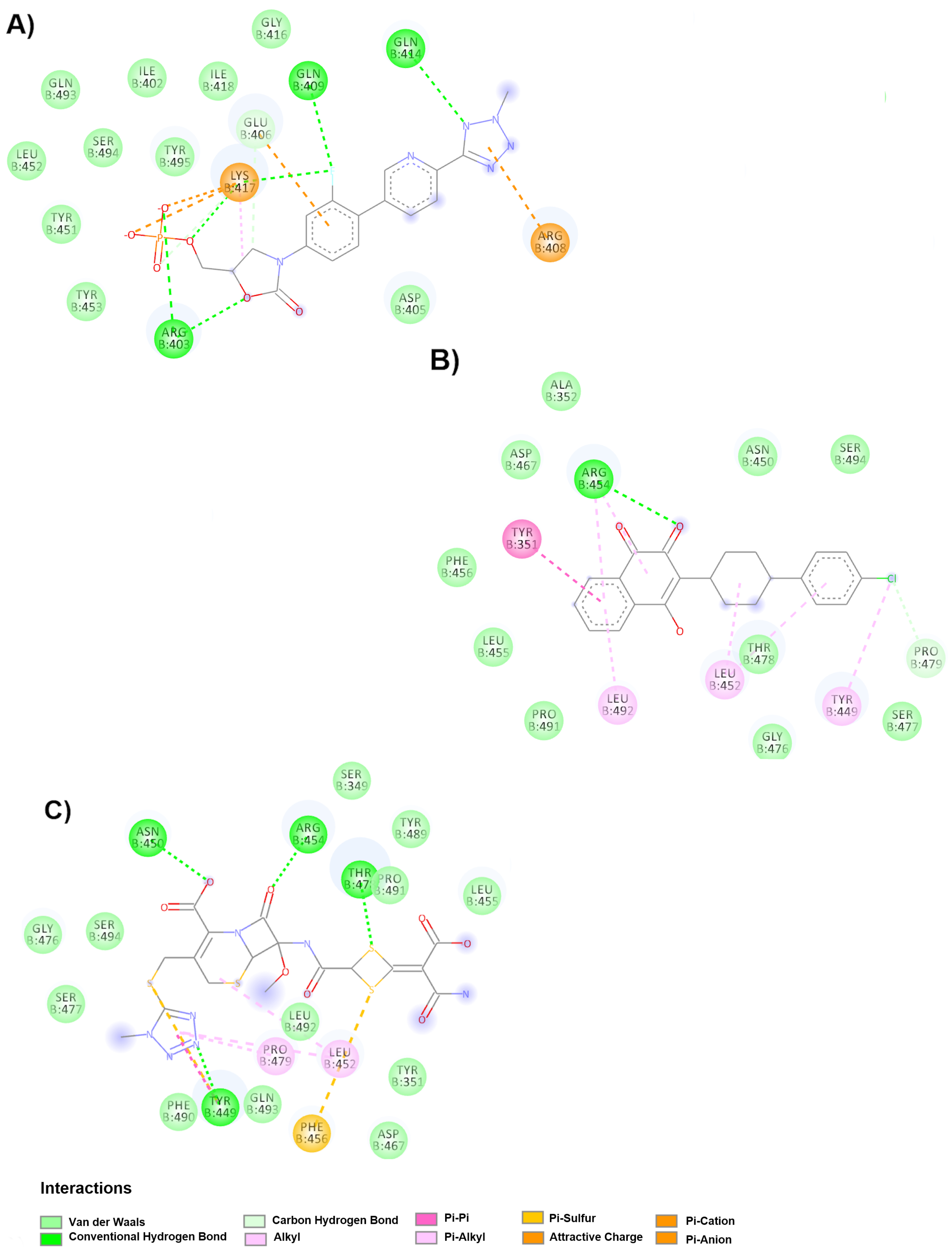
Regarding the hinge region in the down conformation, 3 drugs were found to undergo possible binding and prevent the change to the up conformation, therefore avoiding exposure of the receptor binding site. The drug with the highest affinity is varenicline (−7.6 kcal/mol); however, it is not found in any clinical study, unlike docosahexaenoic acid (DHA) and sulbactam, which are being studied as complementary components of primary treatments. In the DHA clinical trial, the main treatment was fenretinide (LAU-7b), a synthetic retinoid derivative that has been used in the treatment of some types of cancer and cystic fibrosis. DHA is an omega-3 fatty acid and can be included in the diet. Another fatty acid that has been reported in six clinical trials is icosapent ethyl (Vascepa™), although not as part of a dietary food supplement. Its effect is observed on inflammatory biomarkers in individuals with COVID-19. Arachidonic acid (AA) and other unsaturated fatty acids (especially eicosapentaenoic acid, EPA and DHA) are known to inactivate enveloped viruses and inhibit proliferation, so they can serve as endogenous antivirals [
19]. Furthermore, sulbactam is a semisynthetic beta lactamase inhibitor that has been commonly used in conjunction with intravenous ampicillin [
20] to avoid secondary infection in patients with cytokine adsorption in severe COVID-19 pneumonia requiring extracorporeal membrane oxygenation (CYCOV).
As mentioned before, the spike glycoprotein has been shown to be very dynamic due to its hinge sites, and preventing the switch from up to down conformation can help avoid hide the receptor binding site, while combination therapy (with neutralizing antibodies) can prevent viral attachment. There were 5 drugs that were found to bind best at the hinge site in the upper position (zafirlukast, tigecycline, betamethasone, triamcinolone acetonide, atazanavir), three of which are in clinical trials (betamethasone and triamcinolone acetonide, both corticosteroids; and atazanavir, an antiviral). Betamethasone is a steroid from a group of corticosteroids that is used in pregnancies complicated by SARS-CoV-2 due to its immunosuppressive and anti-inflammatory properties. However, a potential diabetogenic effect was found, in addition to the glycemic effects of SARS-CoV-2 and other coronaviruses [
21].
Therefore, the use of betamethasone in clinical trials is not recommended and has not been reported. In contrast, the short-term use of dexamethasone in severe, intubated COVID-19 patients has recently been recommended due to the anti-inflammatory effect that would limit the production and damaging effect of cytokines, but dexamethasone will also inhibit the protective function of T cells and block B cells from making antibodies [
22]. The use of betamethasone and dexamethasone, given the recognition of hinge site blockage, has not been explored, and the two corticosteroids have the same molecular weight and high structural similarity (the only difference is the orientation of the methyl groups on carbon 16), which could have similar effects. The use of inhaled corticosteroids has been associated with lower expression of ACE2 and TMPRSS2, but treatment with triamcinolone acetonide did not decrease the expression of either gene [
23]. On the other hand, the use of atazanavir (protease inhibitor) has been reported in computer studies of inhibitory potency activity with a Kd of 94.94 nM against the SARS-CoV-2 3C-like protease inhibitor [
24]. Clinical trials were beginning recruiting in October.
On the other hand, we found 10 better evaluated drugs from docking studies at the RBM site in the down conformation, which, although it would not prevent the change to the up conformation, would prevent receptor binding. Nine of the drugs show interaction with two amino acids (Leu455 and Gln493) out of the five reported (Leu455, Phe486, Gln493, Ser494 and Asn501) as crucial for the RBM site in the formation of a stable complex between spike viral glycoprotein and human ACE2 [
10,
25]. Only saquinavir interacts with the key amino acid Leu455. Despite these interactions, the affinity for the glycoprotein is different, with the highest affinity being (−12.7 kcal/mol) and the lowest affinity being suvorexant, riciguan, glibenclamide and candesartan (−10.6 kcal/mol). Candesartan was included in four clinical studies due to the selective antagonist function of angiotensin II AT1 receptors. In another study, pancuronium bromide was included as a muscle relaxant in pregnant and postpartum women hospitalized with flu syndrome and COVID-19 (clinical trial NCT04462367). Interestingly, there are some scientific reports available about its possible potential as a drug to treat COVID-19 [
26].
When the structural conformation of the spike glycoprotein in the S1 domain changes to the upper position, it shows high affinity for the receptor, which can be blocked by drugs that bind to the RBM in up position. Six drugs were found to show affinity for the RBM site in the up position. However, the affinity values are lower than the affinity shown in the down position (−9.7 kcal/mol in the up position vs. −12.7 kcal/mol in the down position). The drugs with the highest affinity were tedizolid phosphate and atovaquone (−9.7 kcal/mol). Atovaquone is in a clinical trial (NCT04339426, in combination with azithromycin) to treat COVID-19. Losartan has been reported in 16 clinical trials and has been tested for the treatment of SARS-CoV-2 pneumonia in noncritically ill subjects. On the other hand, the drug with the lowest affinity was rosiglitazone (−8.3 kcal/mol), which was used to regulate glucose in type 2 diabetes; however, its use is discontinued [
27].
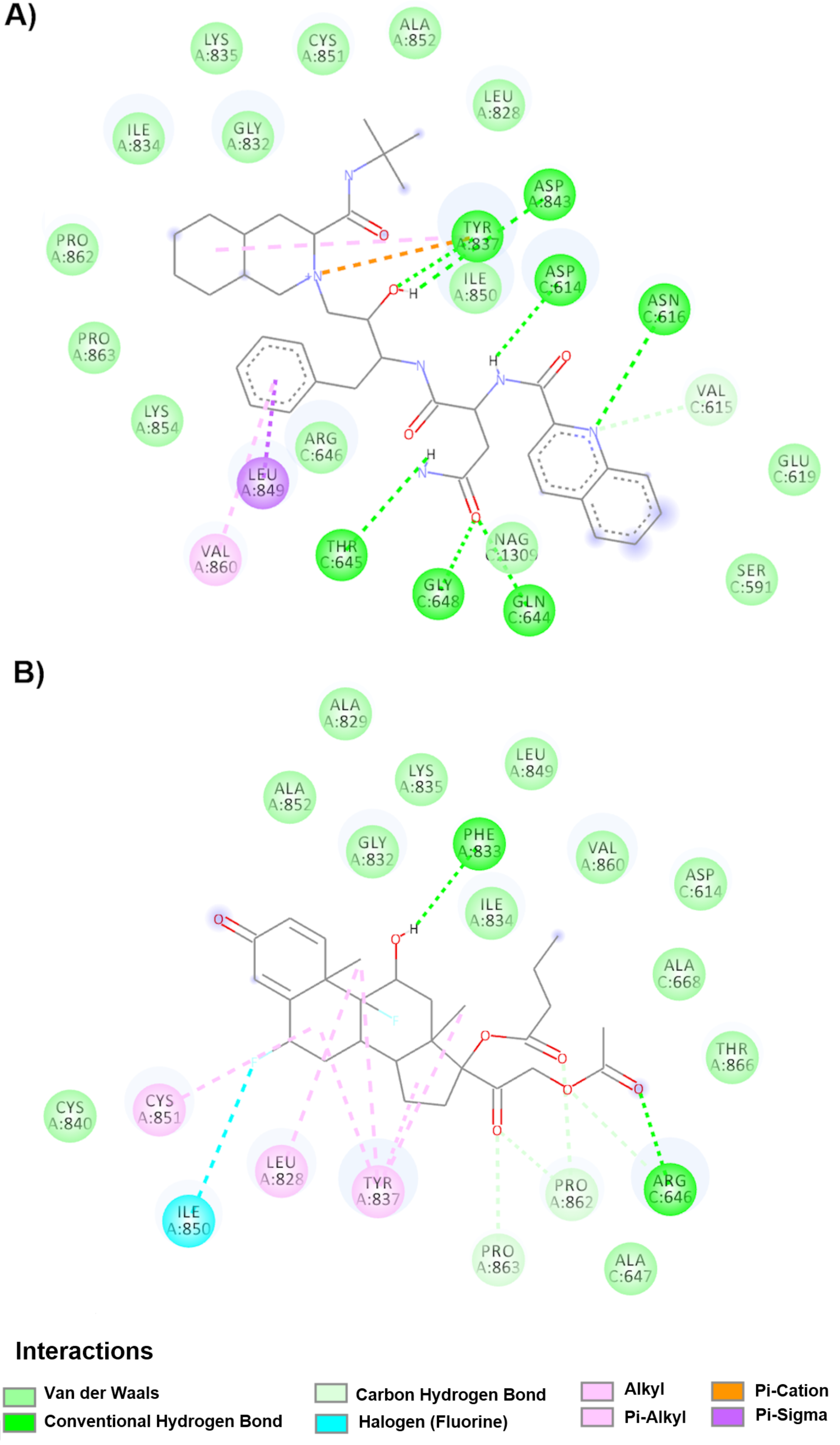
Considering another point in the viral replication cycle, we looked for drugs capable of binding to FP, taking into account that it is a conserved sequence, and SARS-CoV shows two FP sites. FP1 (amino acids 798 to 818) and FP2 (amino acids 816 to 835) have the structural characteristics of an active fusion region; furthermore, they postulate that the regions function cooperatively as an extended FP (FP1-2) [
28,
29]. In the case of the SARS-CoV-2 spike glycoprotein, Tang and collaborators recently determined that FP is found in the region located in amino acids 816 to 855, which corresponds to the region 798–835 for the SARS-CoV spike glycoprotein. The amino acids Leu803, Leu804, Phe823, Cys822, Cys833, Asp830 and Leu831 in SARS-CoV (corresponding to the amino acids Leu821, Leu822, Phe823, Cys840, Asp848, Leu849, and Cys851 in SARS-CoV-2) have been shown to be essential for membrane fusion processes through biophysical studies and site directed mutagenesis studies. [
28]. In addition, these amino acids are conserved in SARS-CoV, MERS-CoV and SARS-CoV-2 [
14].
Notably, nine drugs were bound in the down conformation within the FP and interact mainly with two highly conserved amino acids (Cys851A and Leu849A), which are crucial for the process of membrane fusion in the family of coronaviruses [
14]. However, only nebivolol is currently in an observational clinical trial to compare SARS-CoV-2-positive outpatients and compare all-cause hospitalization and mortality rates between doses of ACEI/ARB- vs. non-ACEI/ARB-based regimens. In this computational study, the drug with the highest affinity was saquinavir (−11.1 kcal/mol), and the lowest affinity was difluprednate (−8.9 kcal/mol). However difluprednate interacts with an additional amino acid to modulate the membrane fusion process (Cys840A) among a total of three critical amino acids (Cys840A, Cys851A and Leu849A). To be able to use these drugs in future clinical studies (in addition to the interactions found), commercial presentation must be considered. Such is the case for terconazole, which is administered as a cream and suppository vaginal presentation (
https://www.accessdata.fda.gov), so it cannot be used for the treatment of COVID-19 with that presentation.
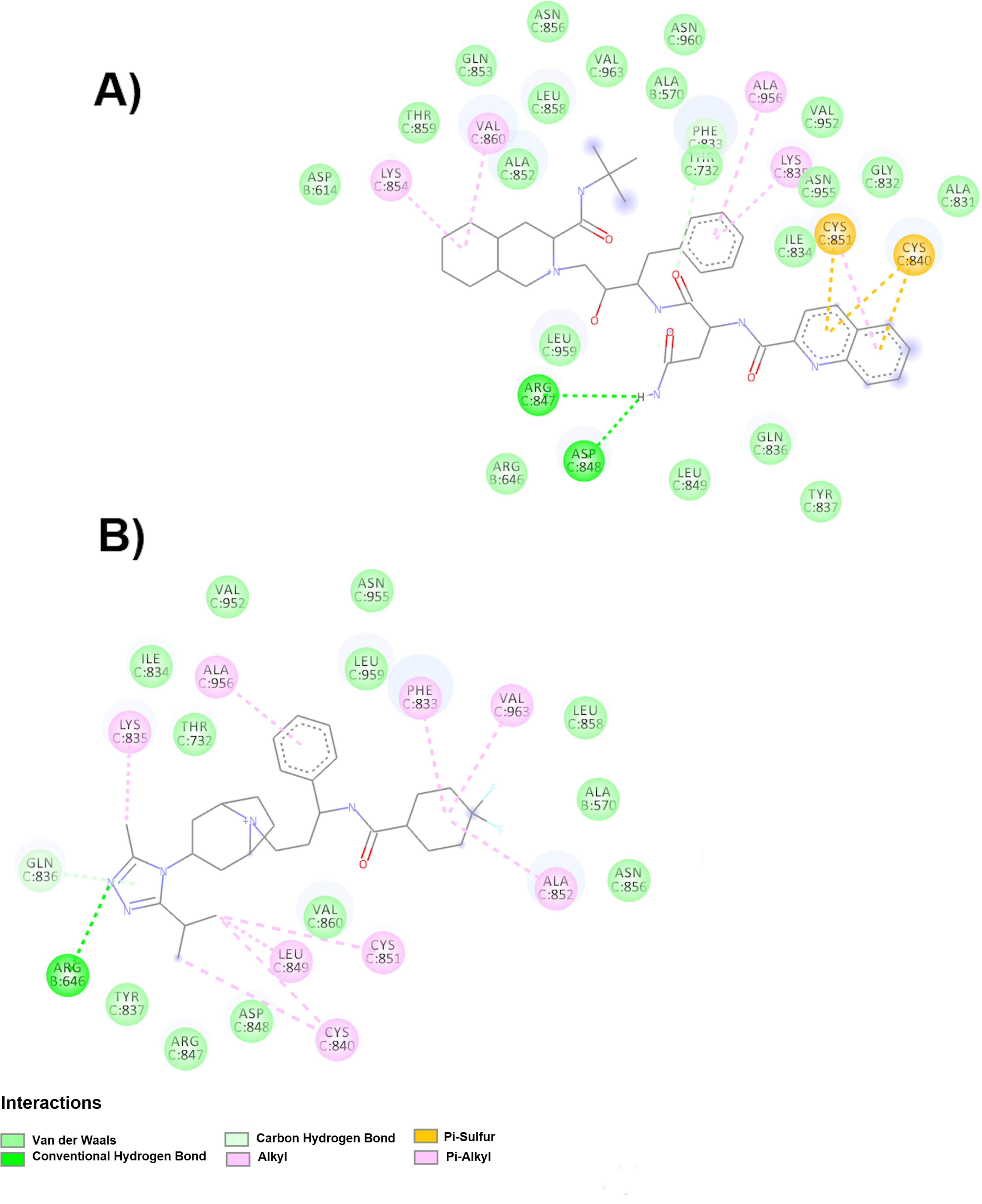
On the other hand, when the protein is in the up position, 10 drugs were found to bind FP, of which five (maraviroc, ritonavir, bosentan, fosinopril sodium, ceftazidime) are in clinical trials. Maraviroc, a C-C chemokine receptor 5 (CCR5) antagonist, is well tolerated without significant side effects in its current use in patients with HIV; in patients with COVID-19, it has been used as a drug against the main protease (Mpro) of SARS-CoV-2 (determined by computer methods) [
30]. The drug found in the largest number of clinical trials is ritonavir, which is mostly used in combination with lopinavir. Both are protease inhibitors to treat HIV and have been used against other coronaviruses (SARS-CoV and MERS-CoV) [
31]. Use against SARS-CoV-2 also seeks to inhibit cellular proteases to prevent completion of the viral replication cycle; however, some clinical trials have not been concluded, and others have concluded that there is no difference in using ritonavir in hospitalized adult patients with severe COVID-19 and standard medical care treatment [
32]. Another class of drugs that have been used for the treatment of COVID-19 is ACE inhibitors, such as fosinopril sodium, which is being studied in two clinical trials (18) as a means to block the virus receptor. Therefore, these three drugs, in addition to the effects described in the clinical trial, could prevent fusion of the viral membrane with the cell membrane.
Furthermore, the drug bosentan is a dual endothelin receptor antagonist used in the treatment of pulmonary hypertension (PHT) [
18], and ceftazidime is an antibiotic used to treat lower respiratory tract infections; both of these drugs and others (i.e., budesonide, cefdinir, cefepime, clindamycin, clobazam, dexamethasone, dexmedetomidine, fosfomycin, dextroamphetamine, etc.) are part of a clinical study called Pharmacokinetics, Pharmacodynamics, and Safety Profile of Understudied Drugs Administered to Children Per Standard of Care (POPS or POP02) in the treatment of COVID-19.
Interestingly, saquinavir has been found to have the highest affinity for FP in both conformations (up and down), showing an affinity of −11 kcal/mol, and it binds to four crucial amino acids in the membrane fusion process (Cys851C, Cys840C, Leu849C and Asp848C). Furthermore, it can bind to the RBM in the down position. Therefore, this drug is a potential candidate to inhibit the FP and RBM sites of the spike glycoprotein.
Considering the data reported, the use of multitarget drugs can be proposed, and by interfering at different points in the replication cycle, it could exert a better effect. It is even possible to propose combined therapies that prevent both viral entry (spike target) and replication cycle events inside the host cell as preventive and therapeutic treatments.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}