2. Materials and Methods
2.1. The Clinical Samples
The Covidiagnostix is a multicenter study, approved by the San Raffaele Hospital, Milan, Italy, Institutional Ethical Review Boards (CE:199/INT/2020), which aims to monitor the antibody response of a population of healthcare professionals (HCPs) who were offered the BNT162b2 mRNA COVID-19 (Comirnaty) vaccine [
13]. This study included 1052 HCPs from the San Raffaele Hospital, Milan, Italy. All HCPs received two doses of the BNT162b2 vaccine (21 ± 1 day interval between the two doses) during January and February 2021; no exclusion criteria were applied. Blood samples were withdrawn for serological evaluation, as previously described [
14].
Eight serum samples were selected from BNT162b2 COVID-19 (Comirnaty
®) eight vaccinated subjects, 21 days after receiving the second dose (
Table S1). Subjects #2 and #5 had a clinical history of SARS-CoV-2 infection before being vaccinated, while the others were never infected by SARS-CoV-2. Sera were selected based on their previous characterization using the electrochemiluminescence immunoassay (ECLIA) Roche Anti-SARS-CoV-2-S test (Roche, Basel, Switzerland), which detects pan-immunoglobulins (IgA, IgG and IgM) against the S receptor binding domain (RBD): four samples had a high anti-RBD titer (#2, #5, #17 and #26), two intermediate titers (#32 and #25) and two with a low titer (#29 and #37) [
13].
2.2. Cell Lines and Viruses
Vero E6 (Vero C1008, clone E6; ATCC CRL-1586) cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with non-essential amino acids (NEAA), penicillin/streptomycin (P/S, 100 U/mL), HEPES buffer (10 mM) and 10% (v/v) heat-inactivated fetal bovine serum (FBS). We added 1 mg/mL of Geneticin (G418) to medium of Vero E6 that stably expressed TMPRSS2 (Vero E6/TMPRSS2, NIBSC 100978). Calu-3 (Human lung cancer cell line, ATCC HTB-55) cells were cultivated in Minimum Essential Medium (MEM) supplemented with NEAA (1×), P/S (100 U/mL), 1 mM sodium pyruvate and 10% (v/v) heat-inactivated FBS. All cell lines were incubated at 37 °C and 5% CO2 in a humidified atmosphere. All cell lines were routinely tested for mycoplasma (Lonza, LT07-218).
Six clinical isolates of SARS-CoV-2 were obtained and propagated in Vero E6 and in Vero E6/TMPRSS2 cells: D614G (GSAID accession ID: EPI_ISL_413489), Alpha (GSAID accession ID: EPI_ISL_1924880), Beta (GSAID accession ID: EPI_ISL_1599180), Gamma (GSAID accession ID: EPI_ISL_1925323), Delta (GSAID accession ID: EPI_ISL_4198505) and Omicron BA1 (GSAID accession ID: EPI_ISL_12188061).
In detail, 0.8 mL of the transport medium of the nasopharyngeal swab (COPAN’s kit UTM® universal viral transport medium—COPAN) was mixed 1:1 with DMEM without FBS and supplemented with P/S and Amphotericin B. The mixture was added to an 80% confluent Vero E6 cell monolayer seeded in a 25 cm2 tissue culture flask. After 1 h adsorption at 37 °C, 3 mL of DMEM supplemented with 2% FBS and Amphotericin B was added. One day post-infection (dpi), the monolayer was washed in PBS, and 4 mL of DMEM supplemented with 2% FBS and Amphotericin B was added. The cytopathic effect was monitored using inverted phase-contrast microscopy (Olympus CKX41), and the supernatant was collected at monolayer complete disruption (3 dpi).
The sample was heat-inactivated at 56 °C for 30 min, and the viral genome was extracted using a QIAamp Viral RNA Mini Kit following the manufacturers’ instructions. h eextracted RNA was processed with the CleanPlex® SARS-CoV-2 Panel (Paragon Genomics, Hayward, CA, USA) and sequenced with the MiSeq Reagent Kit v2 (300-cycles) (Illumina, San Diego, CA, USA) on the MiSeq platform. Genomic reconstruction was performed using the SOPHiA DDM™ platform (SoPHiA Genetics, Boston, MA, USA).
2.3. Virus Titration
Virus stocks were titrated using an Endpoint Dilutions Assay (EDA, TCID50/mL). Vero E6 cells were seeded into 96-well plates and infected at 95% of confluency with base 10 dilutions of virus stock. After 1 h of adsorption at 37 °C, the cell-free virus was removed, cells were washed with PBS 1×, and a complete medium was added to cells. After 72 h, cells were observed to evaluate the presence of a cytopathic effect (CPE). TCID50/mL of viral stocks were then determined with the Reed–Muench formula.
2.4. Pseudovirus Generation
We generated lentiviral pseudoviruses following the protocol already described [
15]. Briefly, 10
6 HEK-293T cells were seeded in a 6-well plate, and 24 h later, were co-transfected using Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) with five plasmids (BEI Resources Repository): pHAGE with CMV-driven Luciferase-IRES-ZsGreen (NR-52516), pHDM with HIV Gag-Pol (NR-52517), pHDM with HIV Tat (NR-52518), pRC with CMV-driven HIV Rev (NR-52519), pHDM-Spike D614G C-term 21 bp deletion (NR-53765). At 24 h post-transfection, the medium was changed with pre-warmed DMEM supplemented with 2% of FBS and incubated at 37 °C and 5% CO
2. At 60 h post-transfection, cell supernatant was harvested and filtered through 0.45 μm filter (Millipore, Burlington, MA, USA) to eliminate cell debris. Pseudoparticles in the media were subsequently pelleted by ultracentrifugation through a 20% sucrose cushion at 26,000 rpm for 3 h by using Beckman 328 SW28 rotor. Pseudoviruses were aliquoted and stored at −80 °C.
2.5. Pseudovirus Titration
To determine pseudovirus titers, we used a luciferase assay. In detail, 4 × 105 cells/mL were seeded in 96-well plates and cultured at 37 °C and 5% CO2 to be confluent after 24 h. The pseudovirus dilutions (toto and 1:2 serial dilutions) were added to the target cells, and spinoculation (1 h at 800 g) followed to allow pseudovirus adsorption. Subsequently, media was removed, and fresh medium was added to cells. The cell supernatant was removed 72 h post-infection, and the cells were lysed with 100 μL of Glo Lysis Buffer (Promega, Madison, WI, USA) for 15 min at room temperature. Cell lysates were transferred to a luminometer plate and 100 μL of Bright-Glo Assay Reagent (Promega) was added immediately before detection (Victor3, Perkin Elmer, Waltham, MA, USA). For further assays, we selected the dilution in which the Relative Luminescence Units (RLUs) were sufficiently (>1000-fold) above the background.
2.6. XTT Assay for Determination of Cell Viability
Cell viability assays were performed using the Cell Proliferation kit II (XTT) (Roche Diagnostics, Merck, Darmstadt, Germany). Briefly, the tetrazolium salt 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium- 5-carboxanilide (XTT) was cleaved by viable cells to form an orange formazan dye that can be quantified photometrically at 450 nm. Before the assay, Vero E6/TMPRSS2 cells (4 × 105 cells/mL) were cultured in 96-well plates for 24 h. The culture medium was replaced by medium containing the inhibitors, and cells were incubated for 72 h. XTT was added to each well, and the plates were incubated for an additional 2 h. The optical density was measured at 450 nm (reference wavelength—650 nm) using a Multiskan GO plate reader (Thermo Scientific Instruments, Waltham, MA, USA). For quantifications, the background levels of media without cultured cells were subtracted.
2.7. Inhibition of Pseudovirus Entry
Target cells (4 × 105 cells/mL) were seeded in 96-well plates and cultured for one day at 37 °C and 5% CO2. Cells were pre-treated with Bafilomycin A1 (100 nM, BFLA-1, Merck), camostat mesylate (100 μM, Merck), alone or in combination, 24 h before the transduction with pseudovirus. Cells were centrifuged at 800 g for 1 h at 37 °C and subsequently incubated for 3 days at 37 °C and 5% CO2. Cells were then lysed with 100 μL of Glo Lysis Buffer (Promega) for 15 min at room temperature. Cell lysates were transferred to a luminometer plate, and 100 μL of Bright-Glo Assay Reagent (Promega) was added immediately before detection (Victor3, Perkin Elmer). We performed six biological replicates for each condition.
2.8. Inhibition of SARS-CoV-2 Entry
Target cells (4 × 105 cells/mL) were seeded in 96-well plates and cultured for one day at 37 °C and 5% CO2. Cells were pre-treated with either one or the combination of BFLA-1 (100 nM) and camostat mesylase (100 μM) 1 h before the infection. Cells were then infected for 1 h with D614G virus variant (0.001 MOI). After virus adsorption, cells were washed three times with PBS and incubated with medium containing the inhibitors and supplemented with 2% FBS. To evaluate the infection inhibition capability of the chemical compounds in Vero and Vero E6/TMPRSS2 cells, we estimated the presence of CPE 48 h post infection (hpi).
The following scoring system was used: 0 = uninfected; 0.5 to 2.5 = increasing number/area of plaques; 3 = all cells infected. Infection control (score 3) was set as 0% infection inhibition, uninfected cells (score 0) as 100% infection inhibition. The whole surface of the wells was considered for the analysis (5× magnification) in inverted phase-contrast microscopy (Olympus CKX41).
In Calu-3 experiments, the supernatants were collected at 48 hpi, and the relative copy number of viral genomes were evaluated using Real Time RT-PCR. For the analysis of the viral entry mechanisms of the selected VOCs, Calu-3 cells were pre-treated with BFLA-1 and camostat mesylate 1 h before the infection (0.1 MOI). Medium with the same concentration of chemical compounds was replaced 24 hpi. The cell supernatants were collected at 72 hpi and analyzed as described above. All the experimental conditions were performed in triplicate.
2.9. Viral RNA Extraction and Real-Time RT-PCR
Viral RNA was purified from cell culture supernatant using the QIAamp Viral RNA Mini Kit (QIAGEN, Hilden, Germany), following the manufacturer’s instructions. Subsequently, the purified RNA was used as template to synthesize the first-strand cDNA, using the SuperScript™ First-Strand Synthesis System for RT-PCR (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacturer’s instruction. Real-time PCR, using SYBR® Green dye-based PCR amplification and detection method, was performed to detect the cDNA.
We used the SYBR™ Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), the forward primer N2F: TTA CAA ACA TTG GCC GCA AA, the reverse primer N2R: GCG CGA CAT TCC GAA and the following PCR conditions: 95 °C for 2 min, 45 cycles of 95 °C for 20 s, annealing at 55 °C for 20 s and elongation at 72 °C for 30 s, followed by a final elongation step at 72 °C for 10 min [
16]. RT-PCR was performed using the ABI-PRISM 7900HT Fast Real-Time instruments (Applied Biosystems, Waltham, MA, USA) by using optical-grade 96-well plates. Samples were run in duplicate in a total volume of 20 μL.
2.10. Real-Time qPCR Analysis of ACE2 and TMPRSS2 Expression Levels
Cellular RNA from Vero E6, Vero E6/TMPRSS2 and Calu-3 (2 × 106 cells) were extracted using the RNeasy Mini Kit (QIAGEN) according to the manufacturer’s protocol. Then, the mRNA from each sample was reverse transcribed using the SuperScript™ III First-Strand Synthesis System for RT-PCR (Thermo Fisher Scientific, Waltham, MA, USA), following the manufacture’s instruction. We analyzed 10 ng of cDNA to evaluate the expression levels of ACE2 and TMPRSS2 with Real Time RT PCR using the SYBR® Green dye-based PCR amplification and detection method.
Gene-specific primers for human ACE2 (FW: AAA CAT ACT GTG ACC CCG CAT; RE: CCA AGC CTC AGC ATA TTG AAC A), monkey ACE2 (FW: AAA CAT ACT GTG ACC CCG CAT; RE: GCT TCA GCA TAT TGA GCA ATT TCT G) and human TMPRSS2 (FW: AAT CGG TGT GTT CGC CTC TAC; RE: CGT AGT TCT CGT TCC AGT CGT). As endogenous control, we used β-actin (FW: CCC TGG ACT TCG AGC AAG AG; RE: ACT CCA TGC CCA GGA AGG AA). Amplification was performed under the following conditions: 94 °C for 5 min, 45 cycles of 94 °C for 30 s, annealing at 60 °C for 30 s and elongation at 68 °C for 30 s, followed by a final elongation step at 72 °C for 10 min. Samples were run in triplicate in a total volume of 20 μL.
2.11. Western Blot Assay
Vero E6, Vero E6/TMPRSS2 and Calu-3 cells (106 cells) were detached with TrypLE (Thermo Fisher, Waltham, MA, USA), pelleted at 700g for 5 min and lysed in RIPA buffer (Thermo Scientific, Waltham, MA, USA) supplemented with an EDTA-free protease inhibitor cocktail tablet (Thermo Scientific, Waltham, MA, USA) for 30 min at 4 °C. Then, lysed cells were clarified at 1000 rpm for 5 min at 4 °C. 10 μL of protein samples were added to 4× LDS sample buffer (Thermo Fisher Scientific, Waltham, MA, USA), loaded into a Bolt 4–12% Bis-Tris Gel (Thermo Fisher Scientific, Waltham, MA, USA) and electrophoresed by SDS-Page at 200 V for 40 min in MES 1× buffer (Thermo Fisher Scientific, Waltham, MA, USA). Proteins were transferred to a polyvinylidene difluoride (PVDF) membrane at 4 °C for overnight in ice-cold Western Transfer Buffer (25 mM Tris, 192 mM Glycine, MeOH 20% (v/v)). The membrane was blocked with 5% BSA in TBS containing 0.1% Tween-20 (PBS-T) for 1 h.
Next, the membrane was incubated for at least 1 h with primary antibodies specific for TMPRSS2 (Abcam, ab109131; diluted 1:1000) and β-actin (Abcam, ab8227; diluted 1:1000). The membranes were washed 3 times in PBS-T, followed by probing with horseradish peroxidase (HRP)-conjugated anti-rabbit antibody (A0545, Merck; diluted 1:4.000) and anti-mouse (A4416, Merck; diluted 1:8.000) as secondary. Signal was developed by treating membranes with SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific, Waltham, MA, USA) imaging on a ChemiDoc MP System (Bio-Rad #12003154, Hercules, CA, USA).
2.12. Immunofluorescence Assays
Vero E6, Vero E6/TMPRSS2 or Calu-3 were seeded on Matrigel® coated slides with a removable 12-well silicone Chamber (Ibidi). After 24 h, the cells were fixed and permeabilized with ice-cold methanol-acetone (1:1) for 15 min at room temperature. Cells were stained with the primary antibody for 1 h at 37 °C: rabbit pAb anti-ACE2 (Sino Biologicals, Beijng, China, 10108-T24) or rabbit pAb anti-TMPRSS2 (Sino Biologicals, 204314-T08). Then, the secondary antibody was added: goat anti-rabbit IgG Alexa Fluor 488 (Invitrogen, A-11008). Cell nuclei were stained with Hoechst 33342 (Thermo Fisher Scientific, Waltham, MA, USA). The images were acquired with Zeiss Axio Observer.Z1 microscope with QImaging Exi-Blue (Carl Zeiss, Oberkochen, Germany) at 20-fold magnification.
The evaluation of the receptor expression was performed with ImageJ software. We calculated the ratio between the total green fluorescent signal and the number of nuclei for both receptors. Then, we normalized these values on reference cell line (Calu-3 cells).
2.13. Kinetic Profiles
Vero E6, Vero E6/TMPRSS2 and Calu-3 (3 × 105 cells/mL) cells were seeded in 96-well plates and cultured for 1 day at 37 °C and 5% CO2 in a humidified atmosphere. Then, the cells were infected with the different SARS-CoV-2 variants (0.001 multiplicity of infection, MOI) in triplicate. After 1 h of adsorption, cells were washed three times with PBS to remove cell-free virus, and fresh medium was added. Cell supernatants were collected at 6 time points: 1, 3, 6, 24, 48 and 72 hpi, and viral genomes were extracted and analyzed by Real-Time RT-PCR.
2.14. Microneutralization Experiments
Cells were seeded into 96-well plates 24 h before the experiment performed at 95% cell confluency for each well. Serum samples were decomplemented by incubation at 56 °C for 30 min, and two dilutions (1:80 and 1:160) were incubated with SARS-CoV-2 at 0.01 MOI for 1 h at 37 °C. Virus–serum mixtures and positive infection control were applied to cells monolayers after a PBS 1× wash, and virus adsorption was performed at 37 °C for 1 h.
Then, the cells were washed with PBS 1× to remove cell-free virus particles and virus-containing mixtures and controls were replaced with complete medium supplemented with 2% FBS. The plates were incubated at 37 °C in the presence of CO2 for 72 h. The experiments were performed in triplicate. Neutralization activity was evaluated by comparing CPE presence detected in the presence of virus–serum mixtures to positive infection control (Vero and Vero E6/TMPRSS2 cells), or by Real-Time RT PCR (Calu-3 cells).
2.15. Statistical Analysis
Two-way ANOVA and Sidak’s multiple comparisons were performed to analyze the RLUs obtained from pseudovirus particles experiments with BFLA1 and camostat. When using BFLA1 and camostat against the whole virus, CPE observed for different experimental settings in Vero E6 cells using, alone or in combination, were normalized to corresponding virus infection control.
For Calu-3 experiments, Real Time-PCR results were analyzed calculating Delta (Δ) Ct as the difference between Ct values obtained for experimental settings and infection control. In both cases, two-way ANOVA and Tukey’s multiple comparisons were performed to analyze the results. ΔCt was calculated also in kinetics profiles experiments with Calu-3 cells as the difference between Ct values obtained for the different time points and Ct(1 h).
Two-way ANOVA and Tukey’s multiple comparisons were performed for both the comparison of the different time points and the general trends in the different cell lines. Sidak’s multiple comparisons was performed to analyze the BFLA-1 and camostat mesylate inhibitory activity against the VOCs. Two-way ANOVA and Dunnett’s multiple comparisons were performed for XTT cell viability evaluation. All the analyses were performed on GraphPad Prism 8.
4. Discussion
An in vitro model that resembles in vivo SARS-CoV-2 entry dynamics would be fundamental to dissect both the viral entry process and the neutralizing antibody response elicited by vaccination.
The Vero E6 cell line has been broadly used to study the lifecycle of several viruses, due to its natural dysregulation of the IFN response [
17,
18]. This represents the most used cell model for SARS-CoV-2 due to the high expression of ACE2 on their surface. However, the absence of TMPRSS2 on the plasma membrane of Vero E6 offers SARS-CoV-2 could only one of the two entry pathways representing a limitation when studying the different entry mechanisms of the SARS-CoV-2 VOCs [
3].
For this reason, we compared three in vitro models: Vero E6, engineered Vero E6 that stably express TMPRSS2 and Calu-3 cells. Since Calu-3 are human-derived lung adenocarcinoma cells, which is closer to the in vivo target of SARS-CoV-2 infection, they were used as a reference to compare the expression of ACE2 and TMPRSS2 [
19].
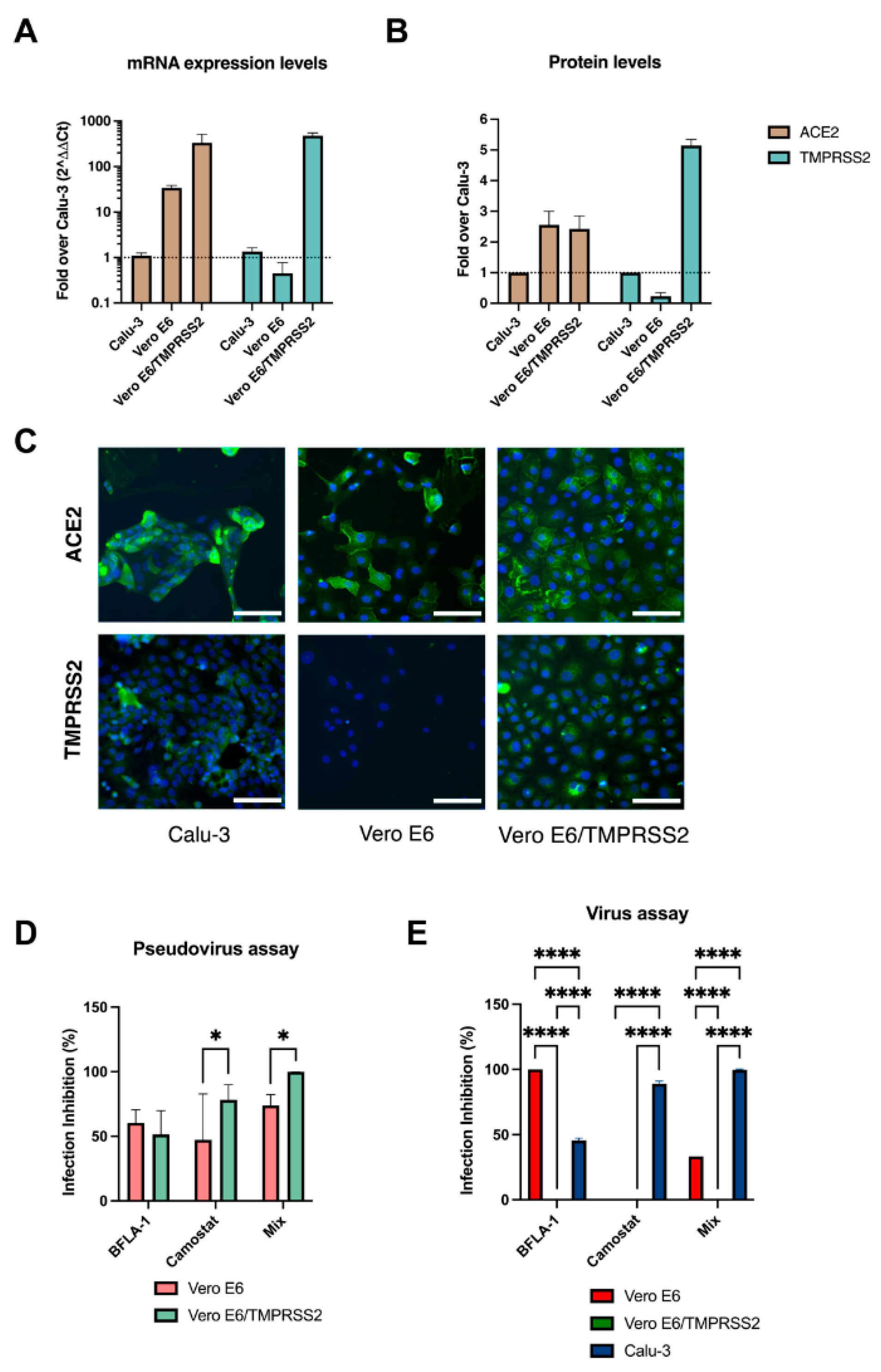
In accordance with the mRNA level investigation, the IF analysis confirmed the discrepancies between the expression levels of ACE2 and TMPRSS2 between Vero E6, Vero E6/TMPRSS2 and Calu-3. ACE2 levels were remarkably higher in Vero E6 and engineered Vero E6 compared to Calu-3. Our studies confirmed the lack of TMPRSS2 expression in Vero E6 cells, which, in contrast, was overexpressed in engineered Vero E6 (four-fold higher) compared to human-derived cells.
Once assessed the expression of host factors involved in viral entry process, we dissected the entry mechanisms of SARS-CoV-2 in the three selected cell lines. First, we used pseudoviral particles since they only recapitulated the viral entry event [
20,
21,
22,
23]. However, we observed important differences between the cell cultures, thus, being an important limit to our aim of comparing the three cell lines. The subsequent use of inhibitors confirmed this imbalance.
We used inhibitors that selectively interfere with the two viral entry processes, camostat mesylate and BFLA-1. The first targets TMPRSS2, hampering its capability to cleave S protein and preventing the direct fusion between the host plasma membrane and the viral envelope. To interfere with the endocytic pathway, we used BFLA-1, a vacuolar H+-ATPase inhibitor. The acid pH environment in the endosomal compartment is fundamental for activating cathepsin L, which is the endosomal protease responsible for the cleavage of S protein and the exposure of the FP domain. Thus, the inhibition of the phagosome-lysosome fusion hampered SARS-CoV-2 entry.
We observed that, to obtain consistent results, we had to used 10-fold more virus in wild type Vero E6 compared to engineered cells. At the best of our knowledge, no previous studies reported these inconsistences between the capability of pseudoviruses to infect these two cell lines. Due to this significant difference in the pseudoparticles titers, we did not continue the characterization on the Calu-3 and decided to use a clinical isolate for the subsequent experiments. In accordance with previous studies, treating Vero E6 cells with BFLA-1 resulted in complete protection from the infection [
3,
6,
24], suggesting that SARS-CoV-2 could enter this cell line only through the endocytic pathway. In our analysis, the same compound showed no effect on Vero E6/TMPRSS2 cell infection, indicating how the overexpression of the serine protease led to the entry of all viral particles at the plasma membrane level.
Interestingly, targeting the TMPRSS2 activity in the engineered Vero E6 with camostat mesylate did not interfere with the viral entry in this cellular model. We speculated that the inhibitor concentration used [
3,
25] was not sufficient to interfere with the activity of the overexpressed TMPRSS2 in the engineered cells. To test our hypothesis, we pre-treated the cells with the highest non-toxic concentration of camostat mesylate; however, it did not block virus entry.
Therefore, we underlined an unbalance entry dynamic in both cellular models: the endocytic or the direct fusion at the plasma membrane were exploited by SARS-CoV-2 in Vero E6 and Vero E6/TMPRSS2, respectively. The unexpected effect of the combination of the two inhibitors on Vero E6 cells may derive from a non-predicted interaction between the BFLA-1 and camostat mesylate. When we tested the mixture in the pseudovirus assay, which recapitulates only the early steps of viral entry to target cells, we did not observe any decrease in its activity. However, we observed a BFLA-1 power loss when tested in combination with camostat in the more complex environment resulting from the infection with the whole virus. It is possible that, when used in a system that recapitulates all the variables involved in the virus infection, its biological activity result is weaker.
We also treated the Calu-3 cells with the two inhibitors to characterize the entry dynamics. The selective inhibition of the two pathways showed a different degree of protection from the infection, implying that SARS-CoV-2 did not exploit equally the two entry mechanisms. In detail, the interference with TMPRSS2 activity displayed a remarkable inhibitory effect than the one obtained targeting the endosomal acidification. Literature data showed that the time required for the entry process depends on the exploited pathway [
6].
Since the endocytic pathway involves many complex mechanisms, it is slower than the direct fusion between the plasma membrane and the viral envelope. Consequently, interfering with the latter resulted in a higher inhibitory effect than the one obtained by the inhibition of the endocytic pathway. However, only hampering both entry mechanisms led to complete protection from viral infection in Calu-3. In contrast to the other two cell lines, Calu-3 allowed the evaluation of both entry pathways. These data suggested that infection of Calu-3 looks similar to the entry of SARS-CoV-2 during natural infection, where the virus could differently exploit both entry mechanisms depending on its biological features and the site of the viral replication.
Point mutations, insertions and deletions were documented in the entire viral genome, especially in the S protein for its exposure to selective pressure. S-gene sequencing is the key for SARS-CoV-2 surveillance, as mutations could impact the different transmissibility of the viral variants or their escape capability from neutralizing antibody response [
26,
27,
28].
From the analysis of replication kinetics of SARS-CoV-2 variants in the selected cell models, we observed that the Alpha variant infects the three cell lines with the same efficiency. On the contrary, our results confirmed the rapid replication of the Delta variant in Calu-3 and Vero E6/TMPRSS2 [
29] while being extremely slow in Vero E6 cells. D614G, Beta, Gamma and Omicron were also slower in Vero E6 cells. However, although for the first variant, the replication kinetics between engineered cells and Calu-3 was similar, for Beta and Omicron, we observed that they were much faster in Vero E6/TMPRSS2 than in Calu-3. Little is known about the possibility that the VOCs differently exploit the two viral entry mechanisms.
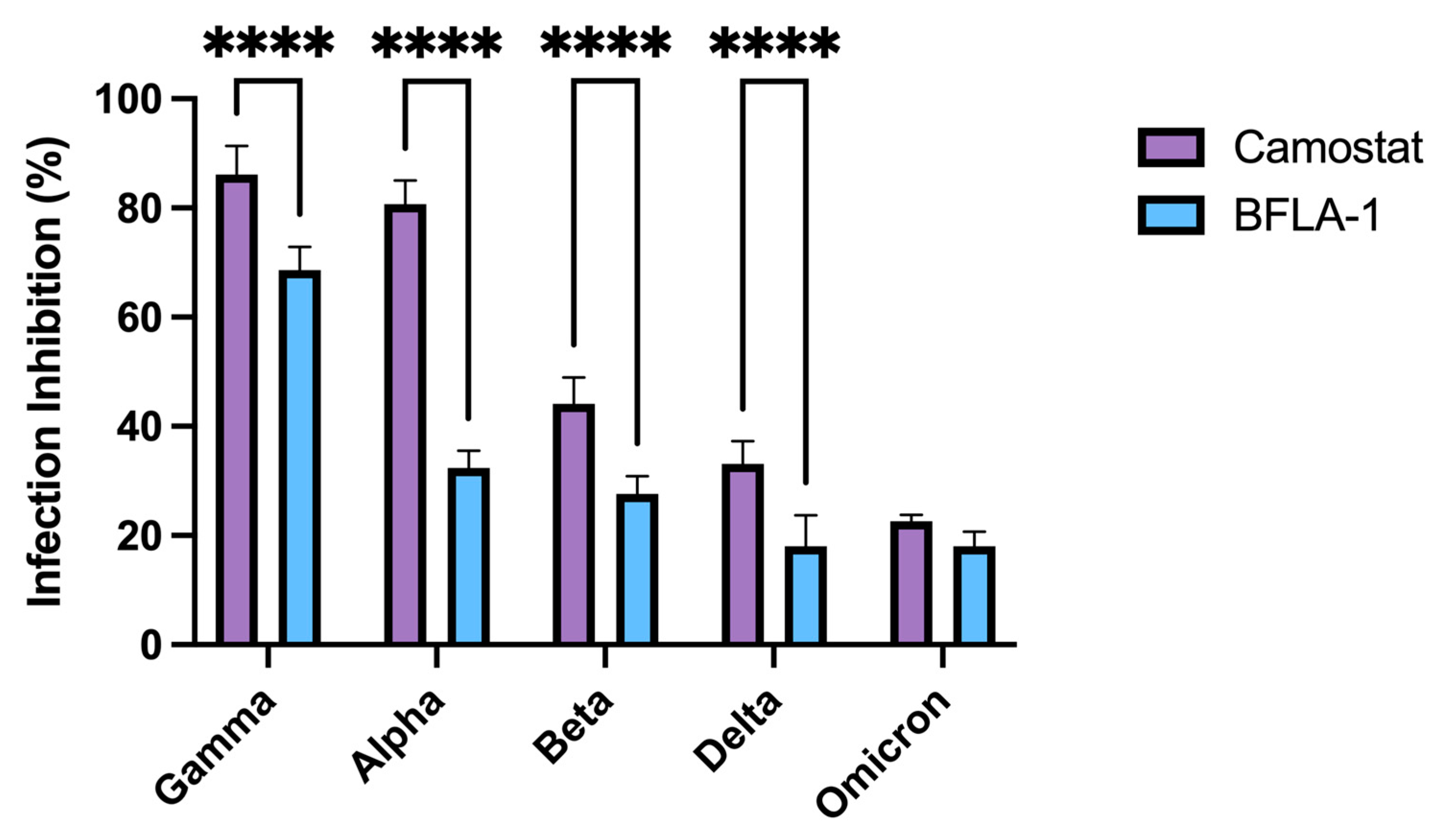
To deeply characterize the observed differences in the replication kinetics of viral variants, we investigated the inhibitory effect of either BFLA-1 or camostat mesylate in Calu-3 cells. Since our data suggested that SARS-CoV-2 uses only one entry mechanism in both Vero E6 and Vero E6/TMPRSS2 cells, the evaluation of the VOC entry mechanism was performed only on the in vitro model that most resembled the physiological target of the infection. We analyzed how the two inhibitors interfered with the entry process of the Alpha, Beta, Gamma, Delta and Omicron variants.
In contrast to what was previously reported [
10,
30], we observed that both compounds equally interfered with the entry process of the Omicron variant, whereas infection from all the other clinical isolates was differently hampered by camostat mesylate or BFLA-1. This characterization underlined the importance of having a model in which the virus could enter host cells through both entry mechanisms. Such model will allow the evaluation of the entry mechanisms exploited by different SARS-CoV-2 variants, which are likely to be those used by the virus during the in vivo infection.
The surveillance and the isolation of viral variants are not only crucial to analyze their global spread and biological features, they also allow determination of whether plasma from vaccinated individuals can neutralize circulating SARS-CoV-2 VOCs. We analyzed the neutralizing capability of sera from eight subjects, selected from a panel of well-characterized samples [
13] against viral variants of epidemiological interest. This evaluation was performed not only on the gold standard model (Vero E6) but also on the other two characterized cell lines. The reference methods to assess the neutralizing activity of sera against SARS-CoV-2 infection are those relying on the CPE evaluation, which include the microneutralization assay [
13,
27,
31,
32,
33,
34,
35]. However, virus infection cannot be assessed through CPE evaluation in the Calu-3 model; therefore, the detection method had to be different.
We tested the Alpha variant because, since its identification in the middle of 2020, it raised concern for its increased transmissibility and rapid spread. Secondly, we assessed the Beta variant: it presents mutations that allowed an easier interaction with ACE2, and it seems to be less sensitive to neutralizing antibodies [
27,
36]. Last, we included the Omicron variant, which emerged at the end of 2021 and quickly spread across the world causing the displacement of the Delta variant. It has been demonstrated that Omicron’s high number of S mutations lead to partial immune evasion from even polyclonal antibody responses, allowing frequent re-infection and vaccine breakthroughs [
11]. Unfortunately, it was not possible to use the Delta variant in this experimental setting for its inability to infect cells lacking TMPRSS2.
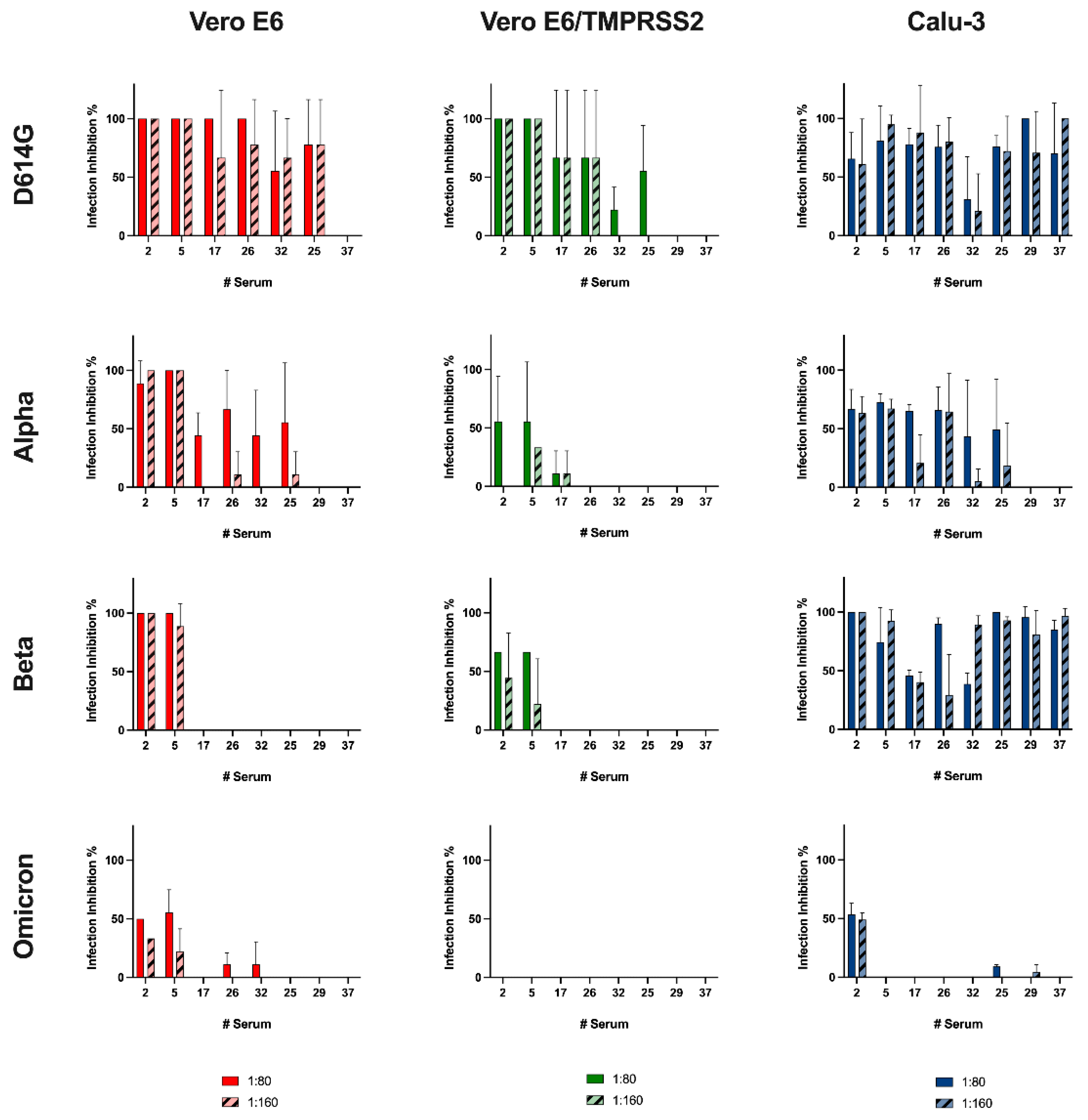
The results obtained from the assay on Vero E6 reflected the literature data: vaccine-induced antibody showed decreasing protection going from Alpha to Beta and to Omicron variants [
27]. On the other hand, we observed almost no protection other than against D614G infection in engineered cells, remarkably, with zero protection against the Omicron variant. Interestingly, an opposite trend was seen using Calu-3 cells: the sera had a remarkable neutralizing effect against the Beta variant compared with against the Alpha variant. Notably, also sera with a low anti-RBD titer inhibited the Beta variant entry at both tested dilutions. Yet, even in this case, no serum except #2 was found to be capable of hampering Omicron infection. Thus, we demonstrated that the choice of an appropriate cell model can have a decisive influence on the determination of the neutralizing capacity of serum.
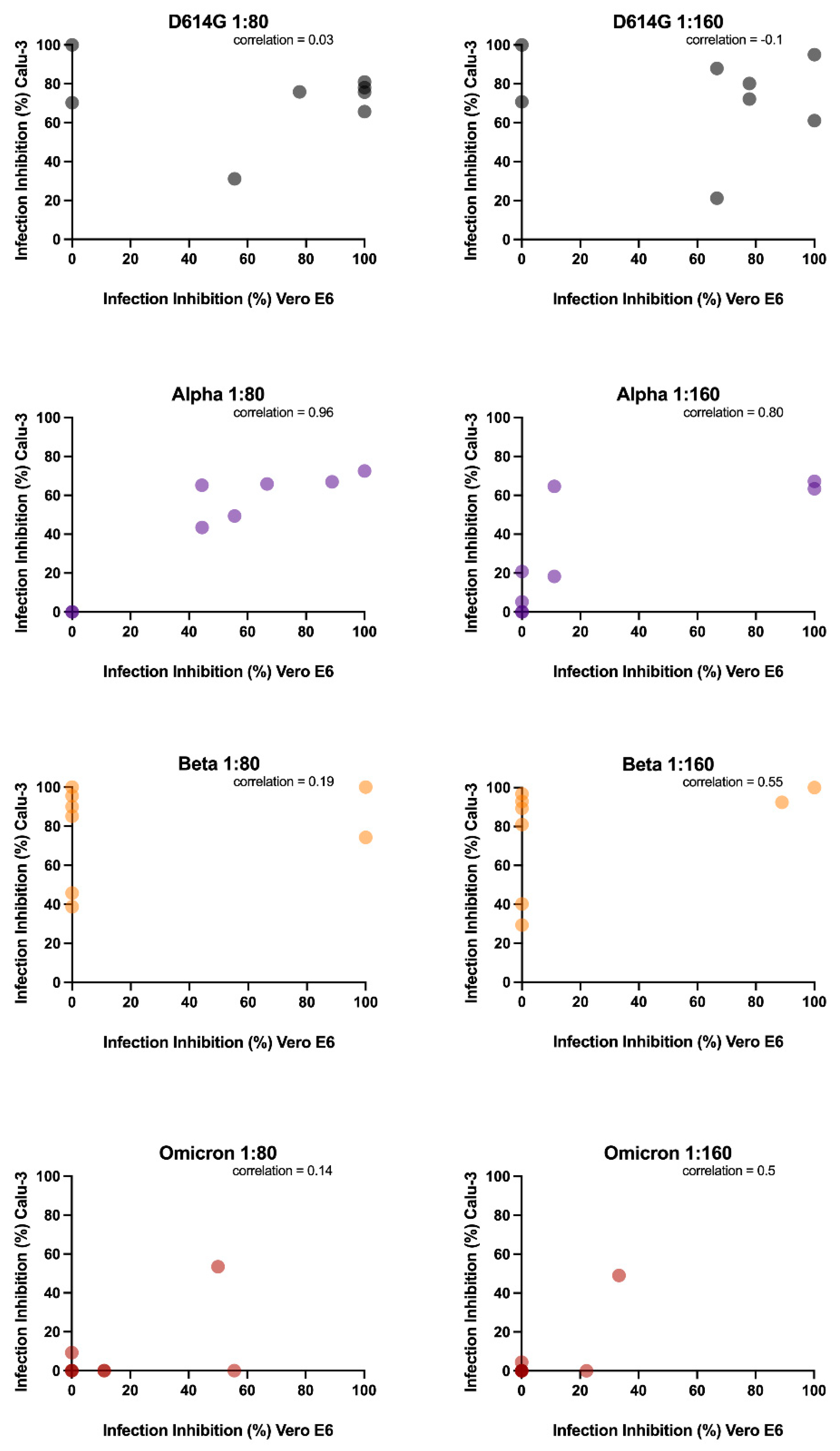
Then, we wanted to assess if Vero E6 and Calu-3, were comparable in dissecting the antibody response against SARS-CoV-2 infection. The impossibility of testing the Delta variant in one of the two models has already suggested how fundamental it is to choose a suitable cellular model. However, to strengthen our hypothesis, we correlated the neutralization data obtained. Different outcomes resulted in considering the four analyzed virus variants. In detail, we found that only the neutralizing data against the Alpha variant observed in Vero E6 experiments are consistent with Calu-3 results, which supports the lack of differences between the cell lines observed in the replication kinetic experiments.
The latter did not happen, however, with the remaining variants analyzed in this study. The replication kinetics of the Alpha variant in Calu-3 were slower than the Beta and Omicron variants, even if the first was more inhibited by camostat mesylate than the others. Additional experiments are required to deeply dissect this aspect and better comprehend the differences between neutralizing experiments in the two cell lines. The antibodies stimulated by the vaccination might target other epitopes of the S protein rather than the RBD, interfering with the TMPRSS2 activity or creating a steric hindrance that prevents the internalization via endocytosis. These molecular aspects deserve further studies aimed at evaluating the serum neutralizing activity in the presence of entry inhibitors.
5. Conclusions
A reliable in vitro model is fundamental to investigate the biological characteristics of SARS-CoV-2 variants, which could give evidence of their spread and infectivity. A better comprehension of the viral entry mechanisms could translate into new therapeutic targets to hamper the first steps of viral infection. To study these mechanisms, pseudoviruses have been widely used due to their safety and versatility. However, in our study, we demonstrated their limits to recapitulate the viral entry process in cells that expressed physiological levels of host factors.
Even though conventional cell lines are easy to handle and enable the simple study of the basics of viral infections, these models do not reflect the native tissues where the early infection stages occur. Monolayer cell cultures cannot recapitulate cellular composition, matrix complexity, tissue diversity and three-dimensional architecture. Our study could be a starting point to further investigate vaccine-induced antibody protection in a platform that provides increased similarity to the in vivo physiology while retaining the benefits of immortalized cell cultures. Even if in vitro models could only partially recapitulate the clinical and immunological features of COVID-19, they could contribute to advances in the study of this infection to identify novel tailored therapies.
As a proof of concept, our data on the antibody neutralizing activity tested on two different cell models showed significant discrepancies. Therefore, these findings suggest that using a system that considers only one of the two viral entry pathways may not fully reflect the neutralizing activity elicited in vaccinated subjects. An appropriate pre-clinical model resembling what occurs during human infection is thus needed to confirm the in vitro observations.
An association between the nasopharyngeal expression of ACE2 and TMPRSS2 genes and the need for oxygen therapy during COVID-19 was described, underlining the importance of considering a cellular model that not only constitutively expresses the two proteins but also has levels that are the closest to the physiological condition [
37]. While the serum neutralizing activity represents, thus far, the best surrogate of protection for COVID-19, it is only one of the correlates of protection from the disease, and, to complete the picture and assess the protection stimulated by vaccination, other immune mechanisms, such as the role of long-lived memory B and T cells, need to be elucidated.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





