
Longitudinal Randomized Cohort Study of SARS-CoV-2 Antibody Seroprevalence in the St. Petersburg Population

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Formation of the Volunteer Cohort
2.2. Vaccination of Volunteers
2.3. Analysis of the Presence of Abs to SARS-CoV-2 Nc and RBD in Volunteers
2.4. Statistical Analysis
3. Results
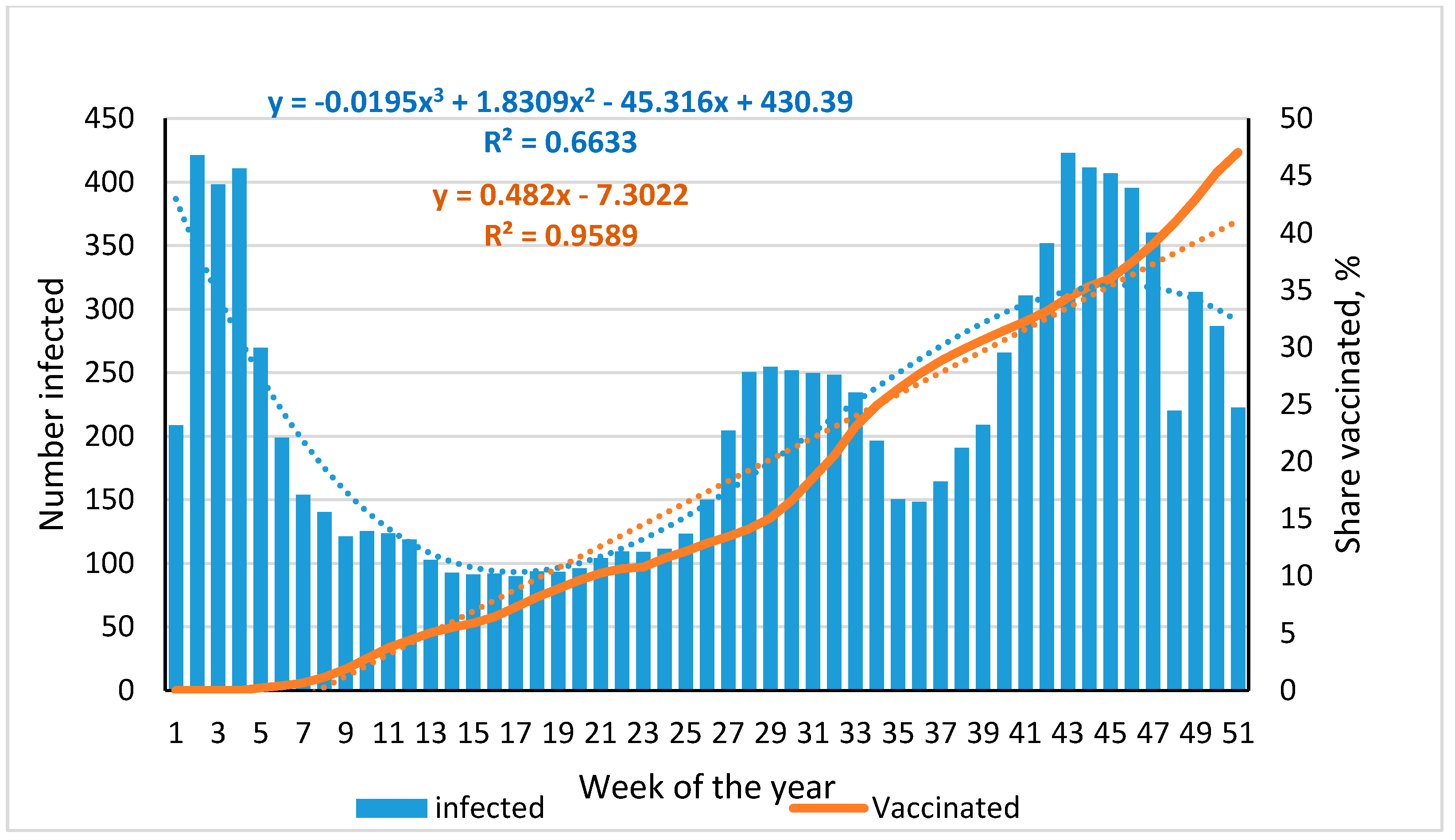
3.1. Dynamics of SARS-CoV-2 Infection and Vaccination of the St. Petersburg Population, 2021
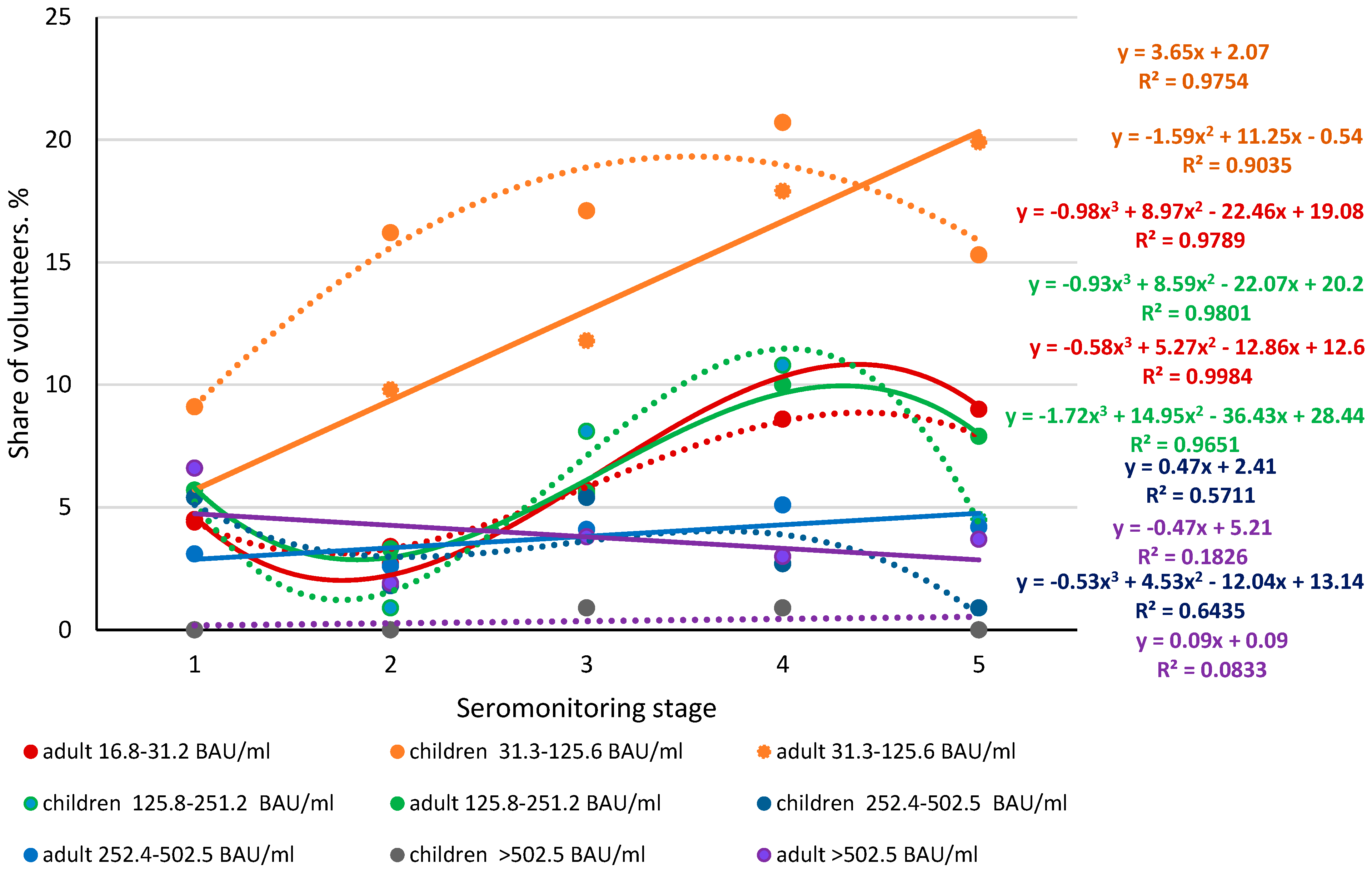
3.2. Seroprevalence of Anti-Nc Abs in the Volunteer Cohort, by Seromonitoring Stage
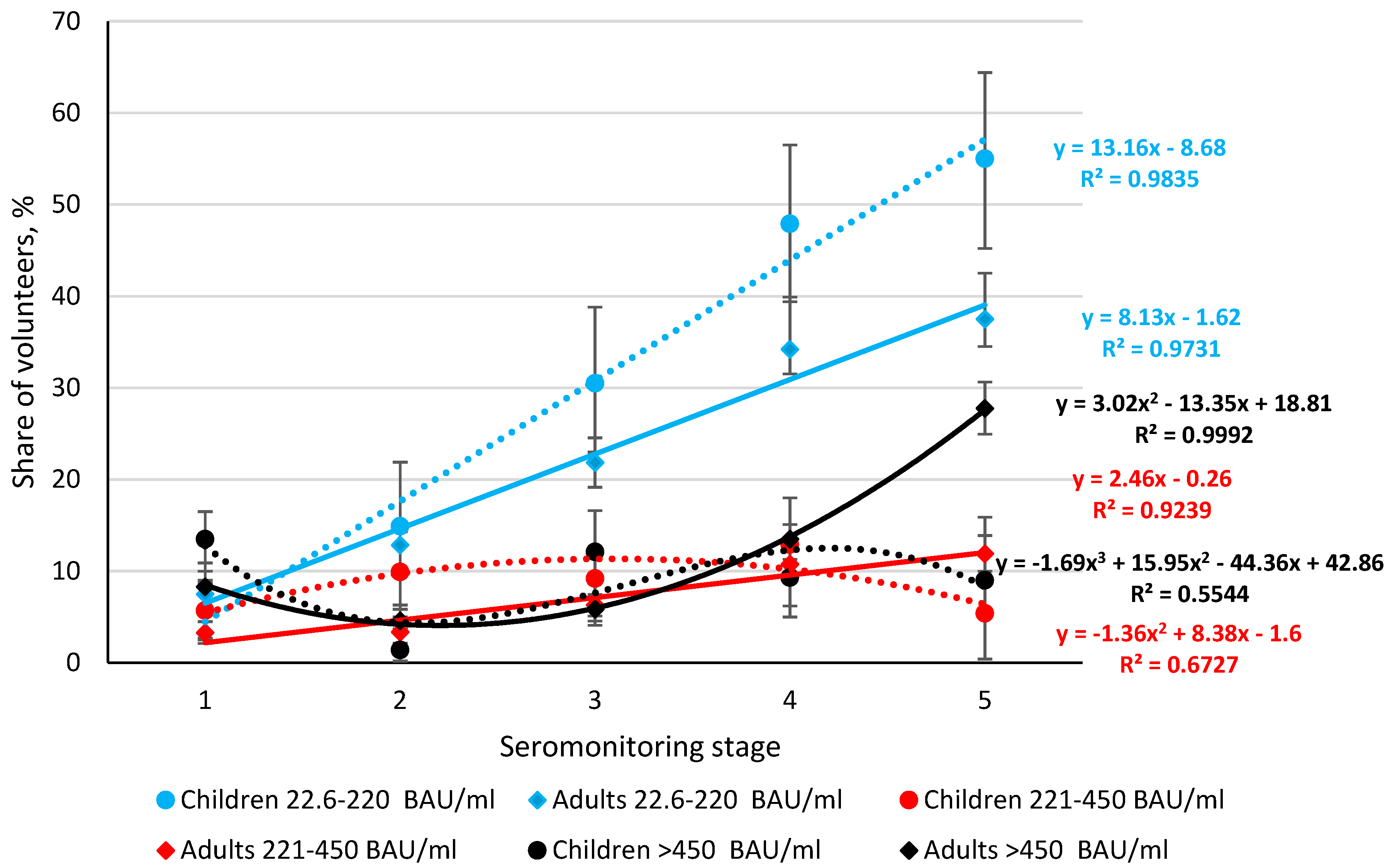
3.3. Seroprevalence of Anti-RBD Abs in the Volunteer Cohort, by Monitoring Stage
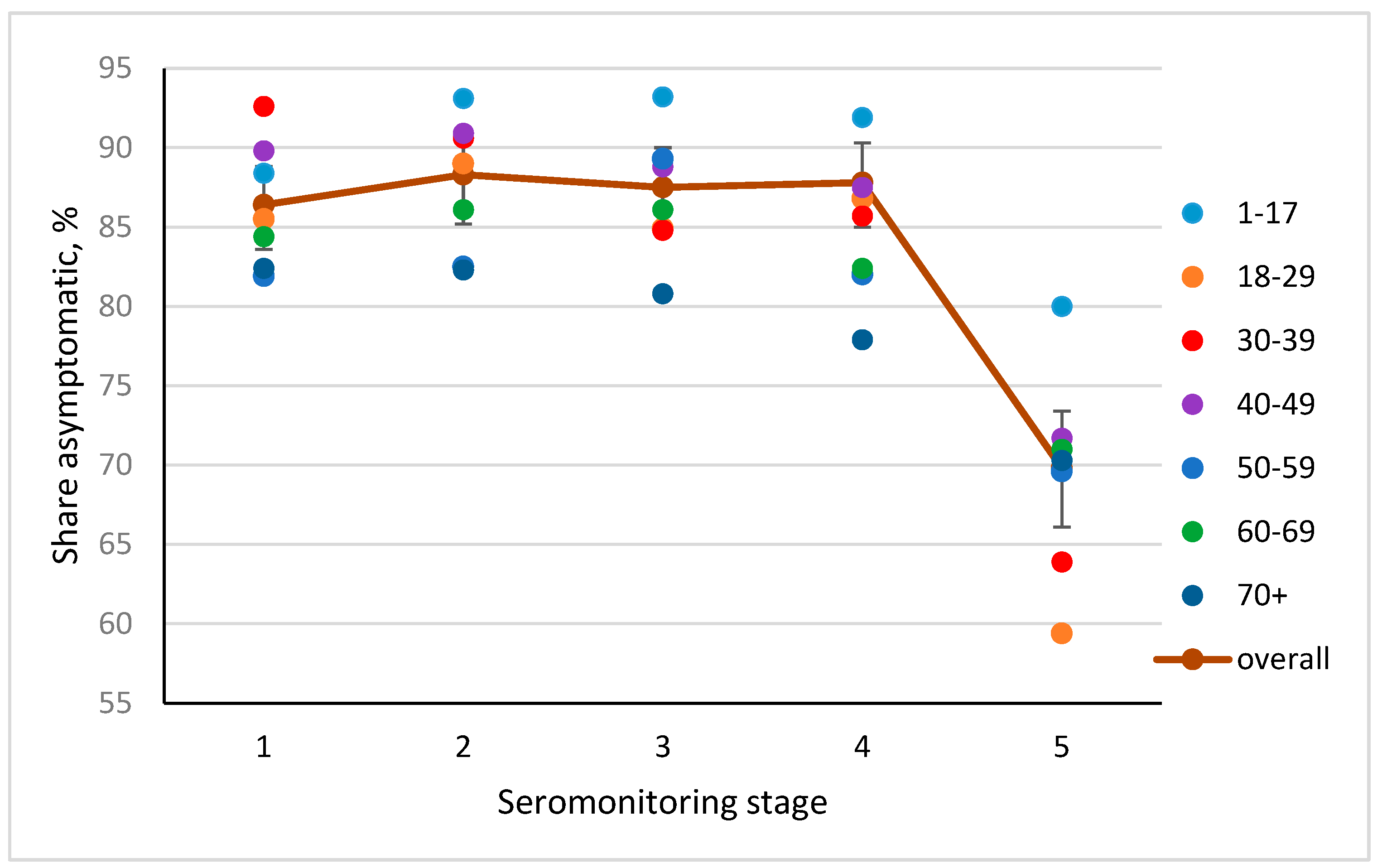
3.4. Frequency of Asymptomatic COVID Forms, by Seromonitoring Stage
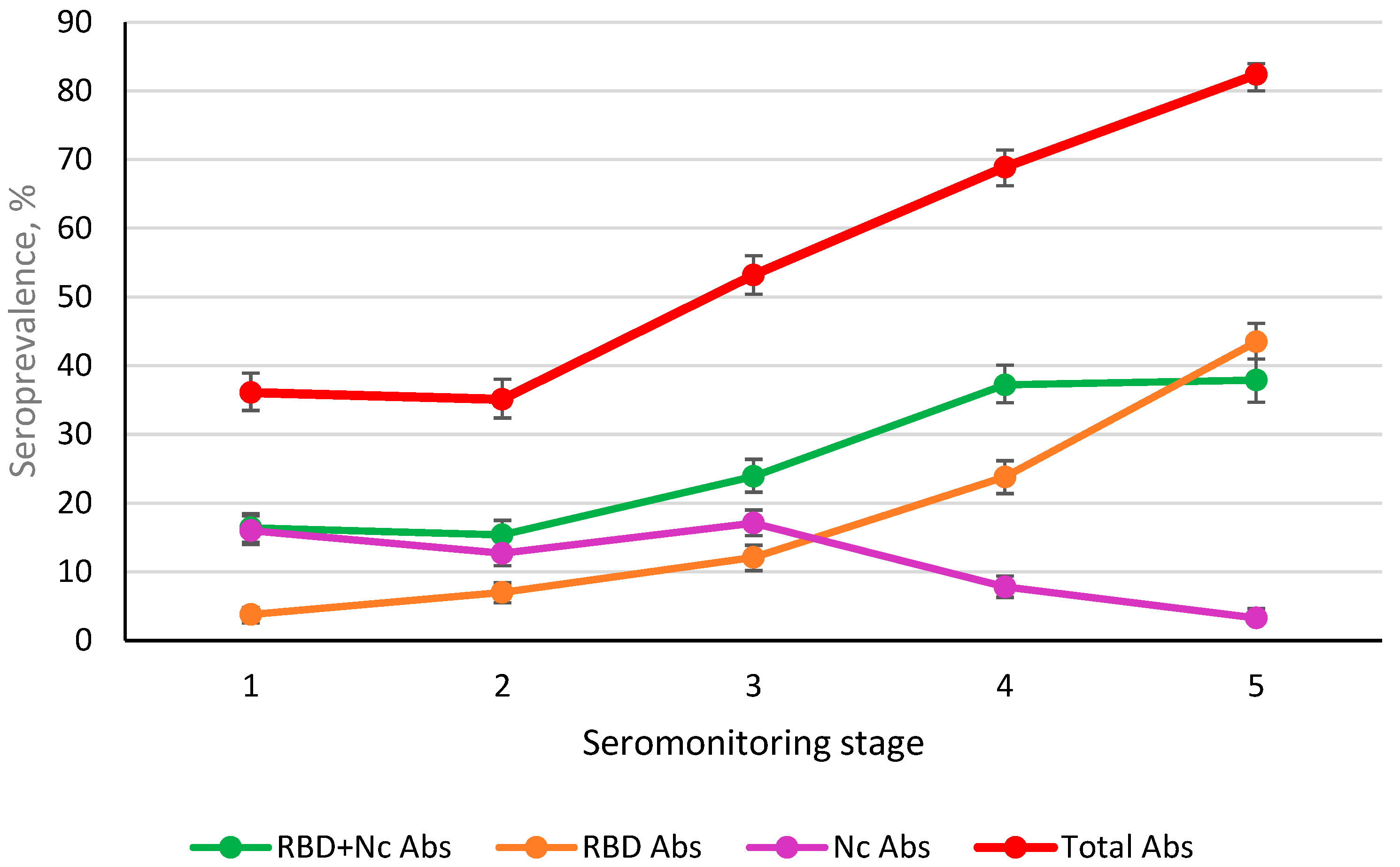
3.5. Post-Infectious and Post-Vaccination Ab Seroprevalence (Nc, RBD) in St. Petersburg Volunteers during Monitoring
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coronavirus-Monitor. Available online: https://coronavirus-monitor.info/ (accessed on 3 November 2021).
- Available online: https://www.gov.spb.ru/press/governor/225782/ (accessed on 16 December 2002).
- Yadegari, I.; Omidi, M.; Smith, S.R. The herd-immunity threshold must be updated for multi-vaccine strategies and multiple variants. Sci. Rep. 2021, 11, 22970. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Zhong, L.; Tan, J.; Zhang, Z.; Lyu, J.; Chen, Y.; Zhao, A.; Huang, L.; Long, Z.; Liu, N.-N.; et al. How to Understand “Herd Immunity” in COVID-19 Pandemic. Front. Cell Dev. Biol. 2020, 8, 547314. [Google Scholar] [CrossRef] [PubMed]
- Fontanet, A.; Cauchemez, S. COVID-19 herd immunity: Where are we? Nat. Rev. Immunol. 2020, 20, 583–584. [Google Scholar] [CrossRef]
- Randolph, H.E.; Barreiro, L.B. Herd Immunity: Understanding COVID-19. Immunity 2020, 52, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gayle, A.A.; Wilder-Smith, A.; Rocklöv, J. The reproductive number of COVID-19 is higher compared to SARS coronavirus. J. Travel. Med. 2020, 27, taaa021. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Yang, H.; Ji, W.; Wu, W.; Chen, S.; Zhang, W.; Duan, G. Virology, Epidemiology, Pathogenesis, and Control of COVID-19. Viruses 2020, 12, 372. [Google Scholar] [CrossRef] [Green Version]
- Sanche, S.; Lin, Y.T.; Xu, C.; Romero-Severson, E.; Hengartner, N.; Ke, R. High Contagiousness and Rapid Spread of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1470–1477. [Google Scholar] [CrossRef]
- Jefferson, T.; Del Mar, C.B.; Dooley, L.; Ferroni, E.; Al-Ansary, L.A.; Bawazeer, G.A.; van Driel, M.L.; Jones, M.A.; Thorning, S.; Beller, E.M.; et al. Physical interventions to interrupt or reduce the spread of respiratory viruses. Cochrane Database Syst. Rev. 2020, 2020, CD006207. [Google Scholar] [CrossRef]
- Al-Betar, M.A.; Alyasseri, Z.A.A.; Awadallah, M.A.; Doush, I.A. Coronavirus herd immunity optimizer (CHIO). Neural Comput. Appl. 2020, 33, 5011–5042. [Google Scholar] [CrossRef]
- Dopico, X.C.; Ols, S.; Loré, K.; Hedestam, G.B.K. Immunity to SARS-CoV-2 induced by infection or vaccination. J. Intern. Med. 2021, 291, 32–50. [Google Scholar] [CrossRef]
- Díaz-Salazar, C.; Sánchez-García, A.; Rodríguez-Gutiérrez, R.; Camacho-Ortiz, A.; Saldívar-Rodríguez, D.; González-González, J.G. Prevalence and associated characteristics of anti-SARS-CoV-2 antibodies in Mexico 5 months after pandemic arrival. BMC Infect. Dis. 2021, 21, 835. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.Y.; Ezhlova, E.B.; Mel’nikova, A.A.; Bashketova, N.S.; Fridman, R.K.; Lyalina, L.V.; Smirnov, V.S.; Chkhindzheriya, I.G.; Grechaninova, T.A.; Agapov, K.A.; et al. Herd Immunity to SARS-CoV-2 among the Population in Saint-Petersburg during the COVID-19 Epidemic. Probl. Osob. Opasnykh Infektsii = Probl. Part. Danger. Infect. 2020, 3, 124–130. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Andreeva, E.E.; Babura, E.A.; Balakhonov, S.V.; Bashketova, N.S.; Bulanov, M.V.; Valeullina, N.N.; Goryaev, D.V.; Detkovskaya, N.N.; Ezhlova, E.B.; et al. Features of developing SARS-CoV-2 nucleocapsid protein population-based seroprevalence during the first wave of the COVID-19 epidemic in the Russian Federation. Russ. J. Infect. Immun. = Infektsiya Immun. 2021, 11, 297–323. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Smirnov, V.S.; Andreeva, E.E.; Babura, E.A.; Balakhonov, S.V.; Bashketova, N.S.; Bugorkova, S.A.; Bulanov, M.V.; Valeullina, N.N.; Vetrov, V.V.; et al. SARS-CoV-2 seroprevalence structure of the russian population during the COVID-19 pandemic. Viruses 2021, 13, 1648. [Google Scholar] [CrossRef] [PubMed]
- Popova, A.Y.; Ezhlova, E.B.; Melnikova, A.A.; Smirnov, V.S.; Lyalina, L.V.; Kozlovskikh, D.N.; Luchinina, S.V.; Romanov, S.V.; Valeullina, N.N.; Dikonskaya, O.V.; et al. Peculiarities of seroprovalence to SARS-CoV-2 in the population of the Middle and Southern Urals in the early period of the COVID-19 pandemic. Epidemiol. Vaccinal Prev. 2021, 20, 8–18. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Smirnov, V.S.; Ezhlova, E.B.; Melnikova, A.A.; Trotsenko, O.E.; Kurganova, O.P.; Detkovskaya, T.N.; Lyalina, L.V.; Abbasova, E.I.; Yurgina, O.M.; et al. Herd immunity to SARS-CoV-2 in the population of the Southern regions of the Far East of Russia. Acta Biomed. Sci. 2021, 6, 253–273. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Andreeva, E.E.; Babura, E.A.; Balakhonov, S.V.; Bashketova, N.S.; Bugorkova, S.A.; Bulanov, M.V.; Valeullina, N.N.; Goryaev, D.V.; Detkovskaya, N.N.; et al. Peculiarities of SARS-CoV-2 nucleocapsid in children during the COVID-19 epidemic of 2020. Pediatriya Named After GN Speransky 2021, 100, 97–106. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Ezhlova, E.B.; Melnikova, A.A.; Smirnov, V.S.; Lyalina, L.V.; Goryaev, D.V.; Khodov, D.A.; Chepizhko, T.G.; Rusin, M.V.; Kuznetsova, N.N.; et al. Structure of herd immunity to SARS-CoV-2 in the Krasnoyarsk region population in the COVID-19 epidemic. Acta Biomed. Sci. 2021, 6, 228–238. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Tarasenko, A.A.; Smolenski, V.Y.; Egorova, S.A.; Smirnov, V.S.; Dashkevich, A.M.; Svetogor, T.N.; Glinskaya, I.N.; Skuranovich, A.L.; Milichkina, A.M.; et al. Herd immunity to SARS-CoV-2 among the population of the Republic of Belarus amid the COVID-19 pandemic. Russ. J. Infect. Immun. = Infektsiya Immun. 2021, 11, 887–904. [Google Scholar] [CrossRef]
- Popova, A.Y.; Kasymov, O.T.; Smolenski, V.Y.; Smirnov, V.S.; Egorova, S.A.; Nurmatov, Z.S.; Milichkina, A.M.; Suranbaeva, G.S.; Khamitova, I.V.; Zueva, E.V.; et al. SARS-CoV-2 Herd Immunity of the Kyrgyz Population in 2021. Res. Sq. 2021; preprint. [Google Scholar] [CrossRef]
- Popova, A.Y.; Totolian, A.A. Methodology for assessing herd immunity to the SARS-CoV-2 virus in the context of the COVID-19 pandemic. Russ. J. Infect. Immun. = Infektsiya Immun. 2021, 11, 609–616. (In Russian) [Google Scholar] [CrossRef]
- WHO. Protocol for a Population-Based Age-Stratified Sero-Epidemiological Study of COVID-19 Infection in Humans. Version 2.0. 2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/332188/WHO-2019CoV-Seroepidemiology-2020.2-rus.pdf (accessed on 18 August 2021).
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatulin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomised phase 1/2 studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Jonesa, I.; Royb, P. Sputnik V COVID-19 vaccine candidate appears safe and effective. Lancet 2021, 397, 642–643. [Google Scholar] [CrossRef]
- Ryzhikov, A.B.; Ryzhikov, E.A.; Bogryantseva, M.P.; Usova, S.V.; Danilenko, E.D.; Nechaeva, E.A.; Pyankov, O.V.; Pyankova, O.G.; Gudymo, A.S.; Bodnev, S.A.; et al. A single blind, placebo-controlled randomized study of the safety, reactogenicity and immunogenicity of the “EpiVacCorona” Vaccine for the prevention of COVID-19, in volunteers aged 18–60 years (phase I–II). Russ. J. Infect. Immun. = Infektsiya Immun. 2021, 11, 283–296. (In Russian) [Google Scholar] [CrossRef]
- Wald, A.; Wolfowitz, J. Confidence Limits for Continuous Distribution Functions. Ann. Math. Stat. 1939, 10, 105–118. Available online: www.jstor.org/stable/2235689 (accessed on 10 October 2021). [CrossRef]
- Agresti, A.; Coull, B.A. Approximate Is Better than “Exact” for Interval Estimation of Binomial Proportions. Am. Stat. 1998, 52, 119–126. [Google Scholar] [CrossRef]
- Significant Difference Calculator (z-Test). RADAR Research Company. Available online: https://radar-research.ru/software/z-test_calculator (accessed on 7 October 2021). (In Russian).
- Del Rio, C.; Omer, S.B.; Malani, P.N. Winter of Omicron—The Evolving COVID-19 Pandemic. JAMA 2021, 22, 319–320. [Google Scholar] [CrossRef]
- Mohiuddin, M.; Kasahara, K. Investigating the aggressiveness of the COVID-19 Omicron variant and suggestions for possible treatment options. Respir. Med. 2021, 191, 106716. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: Omicron and the need for boosters. BMJ 2021, 375, n3079. [Google Scholar] [CrossRef]
- Yang, H.S.; Costa, V.; Racine-Brzostek, S.E.; Acker, K.P.; Yee, J.; Chen, Z.; Karbaschi, M.; Zuk, R.; Rand, S.; Sukhu, A.; et al. Association of Age With SARS-CoV-2 Antibody Response. JAMA Netw. Open 2021, 4, e214302. [Google Scholar] [CrossRef]
- Rosati, M.; Terpos, E.; Ntanasis-Stathopoulos, I.; Agarwal, M.; Bear, J.; Burns, R.; Hu, X.; Korompoki, E.; Donohue, D.; Venzon, D.J.; et al. Sequential Analysis of Binding and Neutralizing Antibody in COVID-19 Convalescent Patients at 14 Months After SARS-CoV-2 Infection. Front. Immunol. 2021, 12, 793953. [Google Scholar] [CrossRef] [PubMed]
- Koerber, N.; Priller, A.; Yazici, S.; Cheng, C.-C.; Mijočević, H.; Wintersteller, H.; Jeske, S.; Vogel, E.; Feuerherd, M.; Tinnefeld, K.; et al. Dynamics of spike-and nucleocapsid specific immunity during long-term follow-up and vaccination of SARS-CoV-2 convalescents. Nat. Commun. 2022, 13, 153. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liang, B.-Y.; Fang, Y.-H.; Wang, H.; Yang, X.-L.; Shen, S.; Chen, L.-K.; Li, S.-M.; Lu, S.-H.; Xiang, T.-D.; et al. Occurrence of COVID-19 Symptoms During SARS-CoV-2 Infection Defines Waning of Humoral Immunity. Front. Immunol. 2021, 12, 722027. [Google Scholar] [CrossRef]
- Sayampanathan, A.A.; Heng, C.S.; Pin, P.H.; Pang, J.; Leong, T.Y.; Leea, V.J. Infectivity of asymptomatic versus symptomatic COVID-19. Lancet 2021, 397, 93–94. [Google Scholar] [CrossRef]
- Almadhi, M.A.; Abdulrahman, A.; Sharaf, S.A.; AlSaad, D.; Stevenson, N.J.; Atkin, S.L.; AlQahtani, M.M. The high prevalence of asymptomatic SARS-CoV-2 infection reveals the silent spread of COVID-19. Int. J. Infect. Dis. 2021, 105, 656–661. [Google Scholar] [CrossRef]
- Jarjour, N.N.; Masopust, D.; Jameson, S.C. T Cell Memory: Understanding COVID-19. Immunity 2021, 54, 14–18. [Google Scholar] [CrossRef]
- De Candia, P.; Prattichizzo, F.; Garavelli, S.; Matarese, G. T Cells: Warriors of SARS-CoV-2 Infection. Trends Immunol. 2021, 42, 18–30. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Vita, R.; Peters, B.; Crotty, S.; Weiskopf, D.; Sette, A. SARS-CoV-2 human T cell epitopes: Adaptive immune response against COVID-19. Cell Host Microbe 2021, 29, 1076–1092. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Niessl, J.; Sekine, T.; Buggert, M. T cell immunity to SARS-CoV-2. Semin. Immunol. 2021, 55, 101505. [Google Scholar] [CrossRef]
- Alkharaan, H.; Bayati, S.; Hellström, C.; Aleman, S.; Olsson, A.; Lindahl, K.; Bogdanovic, G.; Healy, K.; Tsilingaridis, G.; De Palma, P.; et al. Persisting Salivary IgG Against SARS-CoV-2 at 9 Months After Mild COVID-19: A Complementary Approach to Population Surveys. J. Infect. Dis. 2021, 224, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Huynh, A.; Arnold, D.M.; Smith, J.W.; Moore, J.C.; Zhang, A.; Chagla, Z.; Harvey, B.J.; Stacey, H.D.; Ang, J.C.; Clare, R.; et al. Characteristics of Anti-SARS-CoV-2 Antibodies in Recovered COVID-19 Subjects. Viruses 2021, 13, 697. [Google Scholar] [CrossRef] [PubMed]
- Mahallawi, W.; Alzahrani, M.; Alahmadey, Z. Durability of the humoral immune response in recovered COVID-19 patients. Saudi J. Biol. Sci. 2021, 28, 2802–2806. [Google Scholar] [CrossRef] [PubMed]
- Gundlapalli, A.V.; Salerno, R.M.; Brooks, J.T.; Averhoff, F.; Petersen, L.R.; McDonald, L.; Iademarco, M.F. CDC COVID-19 Response. ARS-CoV-2 Serologic Assay Needs for the Next Phase of the US COVID-19 Pandemic Response Open. Forum Infect. Dis. 2020, 8, ofaa555. [Google Scholar] [CrossRef] [PubMed]
- The Coronavirus in SPB. Available online: https://www.gov.spb.ru/covid-19/ (accessed on 9 January 2022).
- Thakur, V.; Ratho, R.K. OMICRON (B.1.1.529): A new SARS-CoV-2 variant of concern mounting worldwide fear. J. Med. Virol. 2021, 94, 1821–1824. [Google Scholar] [CrossRef]
- Kannan, S.R.; Spratt, A.N.; Sharma, K.; Chand, H.S.; Byrareddy, S.N.; Singh, K. Omicron SARS-CoV-2 variant: Unique features and their impact on pre-existing antibodies. J. Autoimmun. 2022, 126, 102779. [Google Scholar] [CrossRef]
- Russell, R.S. Omicron: A Speculation on Its Potential Superpowers. Viral Immunol. 2021, 34, 664–665. [Google Scholar] [CrossRef]
- Majumder, J.; Minko, T. Recent Developments on Therapeutic and Diagnostic Approaches for COVID-19. AAPS J. 2021, 23, 14. [Google Scholar] [CrossRef]
- Gavriatopoulou, M.; Ntanasis-Stathopoulos, I.; Korompoki, E.; Fotiou, D.; Migkou, M.; Tzanninis, I.-G.; Psaltopoulou, T.; Kastritis, E.; Terpos, E.; Dimopoulos, M.A. Emerging treatment strategies for COVID-19 infection. Clin. Exp. Med. 2021, 21, 167–179. [Google Scholar] [CrossRef]
- Smirnov, V.S.; Totolian Areg, A. Some opportunities for immunotherapy in coronavirus infection. Russ. J. Infect. Immun. 2020, 10, 446–458. [Google Scholar] [CrossRef]
- Boyton, R.J.; Altmann, D.M. The immunology of asymptomatic SARS-CoV-2 infection: What are the key questions. Nat. Rev. Immunol. 2021, 21, 762–768. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Cohort (n = 1000) | Vaccinated (n = 131) | |||
|---|---|---|---|---|
| Nc+, n % (95% CI) | Nc−, n % (95% CI) | Nc+, n % (95% CI) | Nc−, n % (95% CI) | |
| RBD+, n | 381 | 223 | 41 | 73 |
| % (95% CI) | 38.1 (35.1–41.3) | 22.3 (19.8–25.1) | 31.3 (23.5–40) | 55.7 (46.8–64.4) |
| RBD−, n | 82 | 212 | 5 | 12 |
| % (95% CI) | 8.2 (6.6–10.1) | 21.2 (28.4–34.2) | 3.8 (1.2–8.7) | 9.2 (4.8–15.0) |
| Total Cohort (n = 1000) | Vaccinated (n = 419) | |||
|---|---|---|---|---|
| Nc+, n % (95% CI) | Nc−, n % (95% CI) | Nc+, n % (95% CI) | Nc−, n % (95% CI) | |
| RBD+, n | 379 | 404 | 132 | 259 |
| % (95% CI) | 37.9 (34.9–41.0) | 40.4 (37.3–43.5) | 36.3 (31.7–41.1) | 61.8 (56.9–66.40) |
| RBD−, n | 41 | 176 | 361.8 | 25 |
| % (95% CI) | 4.1 (3.0–5.5) | 17.6 (15.3–20.1) | (57.0–66.5) | 6.0 (3.9–8.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smirnov, V.S.; Lyalina, L.V.; Milichkina, A.M.; Khamitova, I.V.; Zueva, E.V.; Ivanov, V.A.; Zaguzov, V.S.; Totolian, A.A. Longitudinal Randomized Cohort Study of SARS-CoV-2 Antibody Seroprevalence in the St. Petersburg Population. Viruses 2022, 14, 913. https://doi.org/10.3390/v14050913
Smirnov VS, Lyalina LV, Milichkina AM, Khamitova IV, Zueva EV, Ivanov VA, Zaguzov VS, Totolian AA. Longitudinal Randomized Cohort Study of SARS-CoV-2 Antibody Seroprevalence in the St. Petersburg Population. Viruses. 2022; 14(5):913. https://doi.org/10.3390/v14050913
Chicago/Turabian StyleSmirnov, Vyacheslav S., Liudmila V. Lyalina, Anzhelika M. Milichkina, Irina V. Khamitova, Elena V. Zueva, Valery A. Ivanov, Vitaly S. Zaguzov, and Areg A. Totolian. 2022. "Longitudinal Randomized Cohort Study of SARS-CoV-2 Antibody Seroprevalence in the St. Petersburg Population" Viruses 14, no. 5: 913. https://doi.org/10.3390/v14050913