
Mimicking Gene–Environment Interaction of Higher Altitude Dwellers by Intermittent Hypoxia Training: COVID-19 Preventive Strategies
 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methodology
3. Gene-Environment Interaction: A Basic Perspective to Be Targeted for COVID-19
4. High Altitude Decreased COVID-19: A Dilemmatic Implementation
5. Angiotensin-Converting Enzyme 2 (ACE2) Is the Most Reported Gene for Lower COVID Prevalence in People Living at Higher Elevations
6. IHT Proposed for Future Prevention of Oxygen-Related Diseases
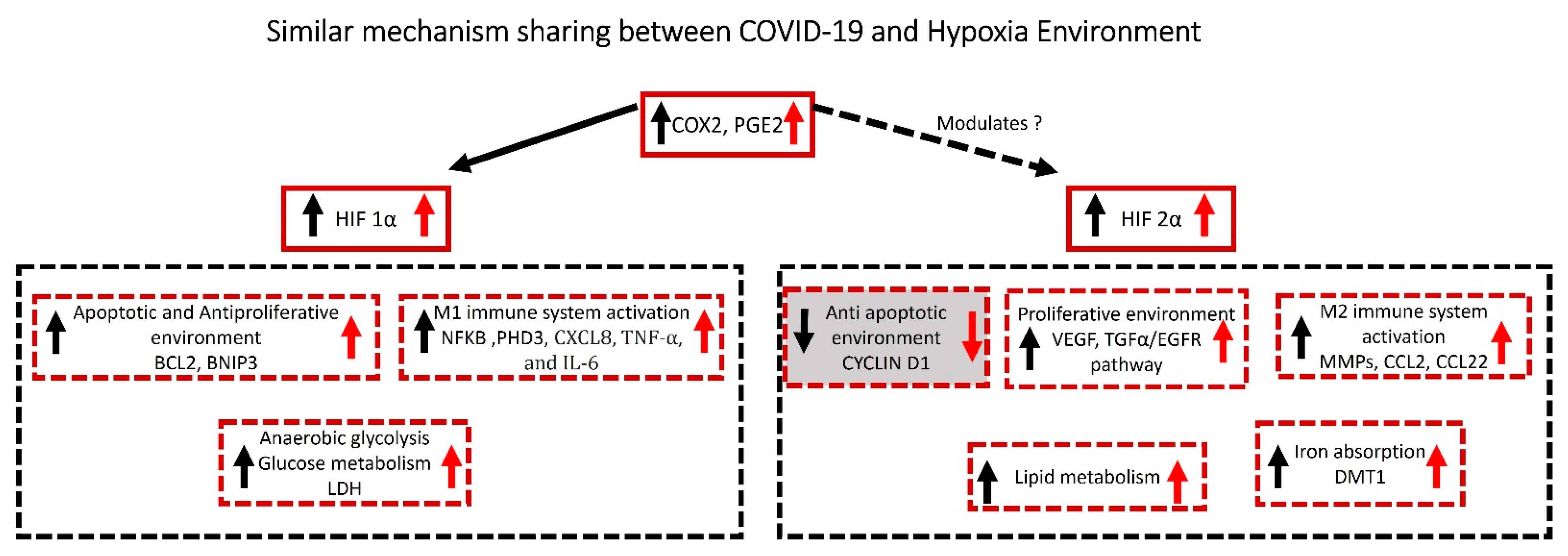
6.1. COVID-19 and Hypoxia Molecules with Similar Mechanisms
6.1.1. Hypoxic Mechanism
6.1.2. Hypoxia Mechanism in COVID-19
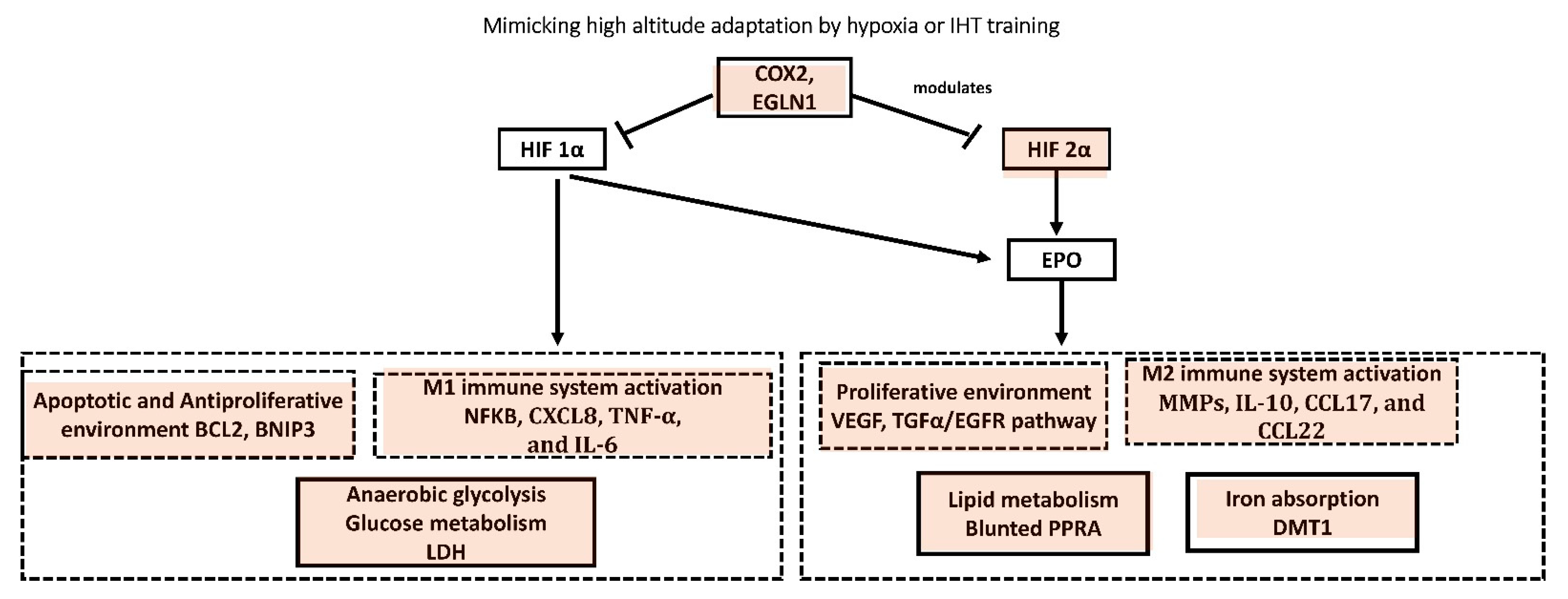
6.1.3. High Altitude Adaptation to Hypoxia
6.1.4. IHT Mimicking High Altitude Adaptation
6.2. The COX Gene as Potential Target to Evaluate the Protective Effects of Hypoxic Training on the Etiology of Respiratory Illnesses
6.2.1. COX as Potential Target for COVID-19
6.2.2. COX as Current Adjunct Therapeutic for COVID-19
6.2.3. Why Is IHT Training Needed in the Current Era?
7. Conclusions and Future Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Litalien, C.; Beaulieu, P. Molecular Mechanisms of Drug Actions. In Pediatric Critical Care; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1553–1568. [Google Scholar]
- Dutta, P.K. COVID-19: Why Coronavirus Is Less Severe in Hills | Research. India Today 2020. Available online: https://www.indiatoday.in/coronavirus-outbreak/story/covid-19-sars-cov-2-heights-acrophobic-latest-research-1685480-2020-06-04 (accessed on 28 March 2022).
- Frisancho, A.R. Human Adaptation and Accommodation; University of Michigan Press: Ann Arbor, MI, USA, 1993; ISBN 978-0472095117. [Google Scholar]
- Millet, G.P.; Debevec, T.; Brocherie, F.; Burtscher, M.; Burtscher, J. Altitude and COVID-19: Friend or foe? A narrative review. Physiol. Rep. 2021, 8, e14615. [Google Scholar] [CrossRef]
- Cai, M.; Chen, X.; Shan, J.; Yang, R.; Guo, Q.; Bi, X.; Xu, P.; Shi, X.; Chu, L.; Wang, L. Intermittent Hypoxic Preconditioning: A Potential New Powerful Strategy for COVID-19 Rehabilitation. Front. Pharmacol. 2021, 12, 643619. [Google Scholar] [CrossRef]
- Trapé, Á.A.; Camacho-Cardenosa, M.; Camacho-Cardenosa, A.; Merellano-Navarro, E.; Rodrigues, J.A.L.; da Silva Lizzi, E.A.; Sorgi, C.A.; Papoti, M.; Brazo-Sayavera, J. Effects of moderate-intensity intermittent hypoxic training on health outcomes of patients recovered from COVID-19: The Aerobicovid study protocol for a randomized controlled trial. Trials 2021, 22, 534. [Google Scholar] [CrossRef]
- Hunter, D.J. Gene–environment interactions in human diseases. Nat. Rev. Genet. 2005, 6, 287–298. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Corella, D.; Demissie, S.; Cupples, L.A.; Couture, P.; Coltell, O.; Wilson, P.W.F.; Schaefer, E.J.; Tucker, K.L. Dietary Fat Intake Determines the Effect of a Common Polymorphism in the Hepatic Lipase Gene Promoter on High-Density Lipoprotein Metabolism. Circulation 2002, 106, 2315–2321. [Google Scholar] [CrossRef] [Green Version]
- Bos, G.; Dekker, J.M.; Feskens, E.J.; Ocke, M.C.; Nijpels, G.; Stehouwer, C.D.A.; Bouter, L.M.; Heine, R.J.; Jansen, H. Interactions of dietary fat intake and the hepatic lipase –480C→T polymorphism in determining hepatic lipase activity: The Hoorn Study. Am. J. Clin. Nutr. 2005, 81, 911–915. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.-L.; Hsu, L.-A.; Hsu, K.-H.; Ko, Y.-H.; Lee, Y.-S. The interactive effects of hepatic lipase gene promoter polymorphisms with sex and obesity on high-density-lipoprotein cholesterol levels in Taiwanese–Chinese. Atherosclerosis 2004, 172, 135–142. [Google Scholar] [CrossRef]
- Cohen, J.E.; Small, C. Hypsographic demography: The distribution of human population by altitude. Proc. Natl. Acad. Sci. USA 1998, 95, 14009–14014. [Google Scholar] [CrossRef] [Green Version]
- Penaloza, D.; Arias-Stella, J. The Heart and Pulmonary Circulation at High Altitudes. Circulation 2007, 115, 1132–1146. [Google Scholar] [CrossRef] [Green Version]
- Arias-Reyes, C.; Zubieta-DeUrioste, N.; Poma-Machicao, L.; Aliaga-Raduan, F.; Carvajal-Rodriguez, F.; Dutschmann, M.; Schneider-Gasser, E.M.; Zubieta-Calleja, G.; Soliz, J. Does the pathogenesis of SARS-CoV-2 virus decrease at high-altitude? Respir. Physiol. Neurobiol. 2020, 277, 103443. [Google Scholar] [CrossRef]
- Huamaní, C.; Velásquez, L.; Montes, S.; Miranda-Solis, F. Propagation by COVID-19 at high altitude: Cusco case. Respir. Physiol. Neurobiol. 2020, 279, 103448. [Google Scholar] [CrossRef]
- Zeng, J.; Peng, S.; Lei, Y.; Huang, J.; Guo, Y.; Zhang, X.; Huang, X.; Pu, H.; Pan, L. Clinical and Imaging features of COVID-19 Patients: Analysis of Data from High-Altitude Areas. J. Infect. 2020, 80, e34–e36. [Google Scholar] [CrossRef]
- Arias-Reyes, C.; Carvajal-Rodriguez, F.; Poma-Machicao, L.; Aliaga-Raduán, F.; Marques, D.A.; Zubieta-DeUrioste, N.; Accinelli, R.A.; Schneider-Gasser, E.M.; Zubieta-Calleja, G.; Dutschmann, M.; et al. Decreased incidence, virus transmission capacity, and severity of COVID-19 at altitude on the American continent. PLoS ONE 2021, 16, e0237294. [Google Scholar] [CrossRef]
- Li, J.-J.; Zhang, H.-Q.; Li, P.-J.; Xin, Z.-L.; Xi, A.-Q.; Ding, Y.-H.; Yang, Z.-P.; Ma, S.-Q. Case series of COVID-19 patients from the Qinghai-Tibetan Plateau Area in China. World J. Clin. Cases 2021, 9, 7032–7042. [Google Scholar] [CrossRef]
- Woolcott, O.O.; Bergman, R.N. Mortality Attributed to COVID-19 in High-Altitude Populations. High Alt. Med. Biol. 2020, 21, 409–416. [Google Scholar] [CrossRef]
- Campos, A.; Scheveck, B.; Parikh, J.; Hernandez-Bojorge, S.; Terán, E.; Izurieta, R. Effect of altitude on COVID-19 mortality in Ecuador: An ecological study. BMC Public Health 2021, 21, 2079. [Google Scholar] [CrossRef]
- Fernandes, J.S.C.; da Silva, R.S.; Silva, A.C.; Villela, D.C.; Mendonça, V.A.; Lacerda, A.C.R. Altitude conditions seem to determine the evolution of COVID-19 in Brazil. Sci. Rep. 2021, 11, 4402. [Google Scholar] [CrossRef]
- Saavedra-Camacho, J.L.; Iglesias-Osores, S. Low COVID-19 Infection Rate in High Altitude Areas. Univ. Médica Pinareña 2021, 17, 1–3. [Google Scholar]
- Pun, M.; Turner, R.; Strapazzon, G.; Brugger, H.; Swenson, E.R. Lower Incidence of COVID-19 at High Altitude: Facts and Confounders. High Alt. Med. Biol. 2020, 21, 217–222. [Google Scholar] [CrossRef]
- Peter Stubenrauch COVID-19 (Coronavirus) & Altitude. Available online: https://www.nationaljewish.org/patients-visitors/patient-info/important-updates/coronavirus-information-and-resources/health-tips/about-covid-19/covid-19-and-altitude (accessed on 1 March 2022).
- Cook-Johnson, R.J.; Demasi, M.; Cleland, L.G.; Gamble, J.R.; Saint, D.A.; James, M.J. Endothelial cell COX-2 expression and activity in hypoxia. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2006, 1761, 1443–1449. [Google Scholar] [CrossRef]
- Brown, J.R.; DuBois, R.N. COX-2: A Molecular Target for Colorectal Cancer Prevention. J. Clin. Oncol. 2005, 23, 2840–2855. [Google Scholar] [CrossRef]
- Liu, X.H.; Kirschenbaum, A.; Lu, M.; Yao, S.; Dosoretz, A.; Holland, J.F.; Levine, A.C. Prostaglandin E2 Induces Hypoxia-inducible Factor-1α Stabilization and Nuclear Localization in a Human Prostate Cancer Cell Line. J. Biol. Chem. 2002, 277, 50081–50086. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Alfajaro, M.M.; Wei, J.; Chow, R.D.; Filler, R.B.; Eisenbarth, S.C.; Wilen, C.B. Cyclooxgenase-2 is induced by SARS-CoV-2 infection but does not affect viral entry or replication. bioRxiv 2020. [Google Scholar] [CrossRef]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HIF-1 versus HIF-2—Is one more important than the other? Vascul. Pharmacol. 2012, 56, 245–251. [Google Scholar] [CrossRef]
- Ratcliffe, P.J. HIF-1 and HIF-2: Working alone or together in hypoxia? J. Clin. Investig. 2007, 117, 862–865. [Google Scholar] [CrossRef] [Green Version]
- Raval, R.R.; Lau, K.W.; Tran, M.G.B.; Sowter, H.M.; Mandriota, S.J.; Li, J.-L.; Pugh, C.W.; Maxwell, P.H.; Harris, A.L.; Ratcliffe, P.J. Contrasting Properties of Hypoxia-Inducible Factor 1 (HIF-1) and HIF-2 in von Hippel-Lindau-Associated Renal Cell Carcinoma. Mol. Cell. Biol. 2005, 25, 5675–5686. [Google Scholar] [CrossRef] [Green Version]
- Takeda, N.; O’Dea, E.L.; Doedens, A.; Kim, J.; Weidemann, A.; Stockmann, C.; Asagiri, M.; Simon, M.C.; Hoffmann, A.; Johnson, R.S. Differential activation and antagonistic function of HIF-α isoforms in macrophages are essential for NO homeostasis. Genes Dev. 2010, 24, 491–501. [Google Scholar] [CrossRef] [Green Version]
- Szlosarek, P.; Charles, K.A.; Balkwill, F.R. Tumour necrosis factor-α as a tumour promoter. Eur. J. Cancer 2006, 42, 745–750. [Google Scholar] [CrossRef]
- Van Ginderachter, J.A.; Movahedi, K.; Hassanzadeh Ghassabeh, G.; Meerschaut, S.; Beschin, A.; Raes, G.; De Baetselier, P. Classical and alternative activation of mononuclear phagocytes: Picking the best of both worlds for tumor promotion. Immunobiology 2006, 211, 487–501. [Google Scholar] [CrossRef]
- Crowther, M.; Brown, N.J.; Bishop, E.T.; Lewis, C.E. Microenvironmental influence on macrophage regulation of angiogenesis in wounds and malignant tumors. J. Leukoc. Biol. 2001, 70, 478–490. [Google Scholar]
- Pollard, J.W. Tumour-educated macrophages promote tumour progression and metastasis. Nat. Rev. Cancer 2004, 4, 71–78. [Google Scholar] [CrossRef]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: Potential targets of anti-cancer therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef]
- Murdoch, C.; Giannoudis, A.; Lewis, C.E. Mechanisms regulating the recruitment of macrophages into hypoxic areas of tumors and other ischemic tissues. Blood 2004, 104, 2224–2234. [Google Scholar] [CrossRef] [Green Version]
- Murdoch, C.; Lewis, C.E. Macrophage migration and gene expression in response to tumor hypoxia. Int. J. Cancer 2005, 117, 701–708. [Google Scholar] [CrossRef]
- Lewis, C.E.; Pollard, J.W. Distinct Role of Macrophages in Different Tumor Microenvironments. Cancer Res. 2006, 66, 605–612. [Google Scholar] [CrossRef] [Green Version]
- Imtiyaz, H.Z.; Williams, E.P.; Hickey, M.M.; Patel, S.A.; Durham, A.C.; Yuan, L.-J.; Hammond, R.; Gimotty, P.A.; Keith, B.; Simon, M.C. Hypoxia-inducible factor 2α regulates macrophage function in mouse models of acute and tumor inflammation. J. Clin. Investig. 2010, 120, 2699–2714. [Google Scholar] [CrossRef] [Green Version]
- Tanner, J.E.; Alfieri, C. The Fatty Acid Lipid Metabolism Nexus in COVID-19. Viruses 2021, 13, 90. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Keith, B.; Simon, M.C.; Vaulont, S.; Peyssonnaux, C. HIF-2α, but not HIF-1α, promotes iron absorption in mice. J. Clin. Investig. 2009, 119, 1159–1166. [Google Scholar] [CrossRef] [Green Version]
- Lorente, L.; Martín, M.M.; González-Rivero, A.F.; Pérez-Cejas, A.; Argueso, M.; Perez, A.; Ramos-Gómez, L.; Solé-Violán, J.; Marcos, Y.; Ramos, J.A.; et al. Blood concentrations of proapoptotic sFas and antiapoptotic Bcl2 and COVID-19 patient mortality. Expert Rev. Mol. Diagn. 2021, 21, 837–844. [Google Scholar] [CrossRef]
- Su, C.-M.; Wang, L.; Yoo, D. Activation of NF-κB and induction of proinflammatory cytokine expressions mediated by ORF7a protein of SARS-CoV-2. Sci. Rep. 2021, 11, 13464. [Google Scholar] [CrossRef]
- Wing, P.A.C.; Keeley, T.P.; Zhuang, X.; Lee, J.Y.; Prange-Barczynska, M.; Tsukuda, S.; Morgan, S.B.; Harding, A.C.; Argles, I.L.A.; Kurlekar, S.; et al. Hypoxic and pharmacological activation of HIF inhibits SARS-CoV-2 infection of lung epithelial cells. Cell Rep. 2021, 35, 109020. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef]
- Guo, Y.; Hu, K.; Li, Y.; Lu, C.; Ling, K.; Cai, C.; Wang, W.; Ye, D. Targeting TNF-α for COVID-19: Recent Advanced and Controversies. Front. Public Heal. 2022, 10, 833967. [Google Scholar] [CrossRef]
- NIH. COVID-19 Treatment Guidelines; NIH: Bethesda, MA, USA, 2022. [Google Scholar]
- Harrison, S.M.; Dove, B.K.; Rothwell, L.; Kaiser, P.; Tarpey, I.; Brooks, G.; Hiscox, J.A. Characterisation of cyclin D1 down-regulation in coronavirus infected cells. FEBS Lett. 2007, 581, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Moutal, A.; Martin, L.F.; Boinon, L.; Gomez, K.; Ran, D.; Zhou, Y.; Stratton, H.J.; Cai, S.; Luo, S.; Gonzalez, K.B.; et al. SARS-CoV-2 spike protein co-opts VEGF-A/neuropilin-1 receptor signaling to induce analgesia. Pain 2021, 162, 243–252. [Google Scholar] [CrossRef]
- Vagapova, E.R.; Lebedev, T.D.; Prassolov, V.S. Viral fibrotic scoring and drug screen based on MAPK activity uncovers EGFR as a key regulator of COVID-19 fibrosis. Sci. Rep. 2021, 11, 11234. [Google Scholar] [CrossRef]
- Gelzo, M.; Cacciapuoti, S.; Pinchera, B.; De Rosa, A.; Cernera, G.; Scialò, F.; Comegna, M.; Mormile, M.; Fabbrocini, G.; Parrella, R.; et al. Matrix metalloproteinases (MMP) 3 and 9 as biomarkers of severity in COVID-19 patients. Sci. Rep. 2022, 12, 1212. [Google Scholar] [CrossRef]
- Islam, H.; Chamberlain, T.C.; Mui, A.L.; Little, J.P. Elevated Interleukin-10 Levels in COVID-19: Potentiation of Pro-Inflammatory Responses or Impaired Anti-Inflammatory Action? Front. Immunol. 2021, 12, 677008. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Ricci, G.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: Further advances in our understanding the role of specific chemokines involved. Cytokine Growth Factor Rev. 2021, 58, 82–91. [Google Scholar] [CrossRef]
- Henry, B.M.; Aggarwal, G.; Wong, J.; Benoit, S.; Vikse, J.; Plebani, M.; Lippi, G. Lactate dehydrogenase levels predict coronavirus disease 2019 (COVID-19) severity and mortality: A pooled analysis. Am. J. Emerg. Med. 2020, 38, 1722–1726. [Google Scholar] [CrossRef]
- Mylonis, I.; Simos, G.; Paraskeva, E. Hypoxia-Inducible Factors and the Regulation of Lipid Metabolism. Cells 2019, 8, 214. [Google Scholar] [CrossRef] [Green Version]
- Hari Kishore, A.; Li, X.-H.; Word, R.A. Hypoxia and PGE2 Regulate MiTF-CX During Cervical Ripening. Mol. Endocrinol. 2012, 26, 2031–2045. [Google Scholar] [CrossRef]
- Wei, C.; Wang, H.; Liu, G.; Zhao, F.; Kijas, J.W.; Ma, Y.; Lu, J.; Zhang, L.; Cao, J.; Wu, M.; et al. Genome-wide analysis reveals adaptation to high altitudes in Tibetan sheep. Sci. Rep. 2016, 6, 26770. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.; Wang, K.; Gan, X.; Li, L.; Zhang, Q.; Xiang, J.; Yuan, X.; Zhang, Y.; Wang, Y. Prophylaxis of ibuprofen in acute mountain sickness. Medicine 2020, 99, e23233. [Google Scholar] [CrossRef]
- Rankin, E.B.; Biju, M.P.; Liu, Q.; Unger, T.L.; Rha, J.; Johnson, R.S.; Simon, M.C.; Keith, B.; Haase, V.H. Hypoxia-inducible factor–2 (HIF-2) regulates hepatic erythropoietin in vivo. J. Clin. Investig. 2007, 117, 1068–1077. [Google Scholar] [CrossRef]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B.; et al. Genetic Evidence for High-Altitude Adaptation in Tibet. Science 2010, 329, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Wilding, J.P.; Gause-Nilsson, I.; Persson, A. Tesaglitazar, as add-on therapy to sulphonylurea, dose-dependently improves glucose and lipid abnormalities in patients with type 2 diabetes. Diabetes Vasc. Dis. Res. 2007, 4, 194–203. [Google Scholar] [CrossRef]
- Beall, C.M.; Cavalleri, G.L.; Deng, L.; Elston, R.C.; Gao, Y.; Knight, J.; Li, C.; Li, J.C.; Liang, Y.; McCormack, M.; et al. Natural selection on EPAS1 (HIF2α) associated with low hemoglobin concentration in Tibetan highlanders. Proc. Natl. Acad. Sci. USA 2010, 107, 11459–11464. [Google Scholar] [CrossRef] [Green Version]
- Yi, X.; Liang, Y.; Huerta-Sanchez, E.; Jin, X.; Cuo, Z.X.P.; Pool, J.E.; Xu, X.; Jiang, H.; Vinckenbosch, N.; Korneliussen, T.S.; et al. Sequencing of 50 Human Exomes Reveals Adaptation to High Altitude. Science 2010, 329, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Bigham, A.W.; Mao, X.; Mei, R.; Brutsaert, T.; Wilson, M.J.; Julian, C.G.; Parra, E.J.; Akey, J.M.; Moore, L.G.; Shriver, M.D. Identifying positive selection candidate loci for high-altitude adaptation in Andean populations. Hum. Genom. 2009, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Bigham, A.; Bauchet, M.; Pinto, D.; Mao, X.; Akey, J.M.; Mei, R.; Scherer, S.W.; Julian, C.G.; Wilson, M.J.; López Herráez, D.; et al. Identifying Signatures of Natural Selection in Tibetan and Andean Populations Using Dense Genome Scan Data. PLoS Genet. 2010, 6, e1001116. [Google Scholar] [CrossRef] [Green Version]
- Levett, D.Z.H.; Viganò, A.; Capitanio, D.; Vasso, M.; De Palma, S.; Moriggi, M.; Martin, D.S.; Murray, A.J.; Cerretelli, P.; Grocott, M.P.W.; et al. Changes in muscle proteomics in the course of the Caudwell Research Expedition to Mt. Everest. Proteomics 2015, 15, 160–171. [Google Scholar] [CrossRef]
- Pham, K.; Parikh, K.; Heinrich, E.C. Hypoxia and Inflammation: Insights from High-Altitude Physiology. Front. Physiol. 2021, 12, 676782. [Google Scholar] [CrossRef]
- Kuang, L.; Zheng, Y.; Lin, Y.; Xu, Y.; Jin, S.; Li, Y.; Dong, F.; Jiang, Z. High-Altitude Adaptation of Yak Based on Genetic Variants and Activity of Lactate Dehydrogenase-1. Biochem. Genet. 2010, 48, 418–427. [Google Scholar] [CrossRef]
- Verma, P.; Sodhi, M.; Bharti, D.V.; Kumar, P.; Giri, A.; Sharma, A.; Das, P.; Niranjan, S.; Kataria, R.; Kumari, P.; et al. VEGF-A locus plausibly associated with high altitude adaptation in yak is completely fixed in cattle population from high and low altitude environments. Int. J. Anim. Biotechnol. 2016, 6, 16–20. [Google Scholar]
- Jin, M.; Lu, J.; Fei, X.; Lu, Z.; Quan, K.; Liu, Y.; Chu, M.; Di, R.; Wei, C.; Wang, H. Selection Signatures Analysis Reveals Genes Associated with High-Altitude Adaptation in Tibetan Goats from Nagqu, Tibet. Animals 2020, 10, 1599. [Google Scholar] [CrossRef]
- Verma, P.; Sharma, A.; Sodhi, M.; Thakur, K.; Kataria, R.S.; Niranjan, S.K.; Bharti, V.K.; Kumar, P.; Giri, A.; Kalia, S.; et al. Transcriptome Analysis of Circulating PBMCs to Understand Mechanism of High Altitude Adaptation in Native Cattle of Ladakh Region. Sci. Rep. 2018, 8, 7681. [Google Scholar] [CrossRef] [Green Version]
- Ge, R.-L.; Simonson, T.S.; Gordeuk, V.; Prchal, J.T.; McClain, D.A. Metabolic aspects of high-altitude adaptation in Tibetans. Exp. Physiol. 2015, 100, 1247–1255. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Peyssonnaux, C. The gut in iron homeostasis: Role of HIF-2 under normal and pathological conditions. Blood 2013, 122, 885–892. [Google Scholar] [CrossRef] [Green Version]
- Mounier, R.; Pialoux, V.; Cayre, A.; Schmitt, L.; Richalet, J.-P.; Robach, P.; Lasne, F.; Roels, B.; Millet, G.; Coudert, J.; et al. Leukocyte’s Hif-1 Expression and Training-Induced Erythropoietic Response in Swimmers. Med. Sci. Sport. Exerc. 2006, 38, 1410–1417. [Google Scholar] [CrossRef]
- Lundby, C.; Gassmann, M.; Pilegaard, H. Regular endurance training reduces the exercise induced HIF-1α and HIF-2α mRNA expression in human skeletal muscle in normoxic conditions. Eur. J. Appl. Physiol. 2006, 96, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Płoszczyca, K.; Langfort, J.; Czuba, M. The Effects of Altitude Training on Erythropoietic Response and Hematological Variables in Adult Athletes: A Narrative Review. Front. Physiol. 2018, 9, 375. [Google Scholar] [CrossRef] [Green Version]
- Czuba, M.; Wilk, R.; Karpiński, J.; Chalimoniuk, M.; Zajac, A.; Langfort, J. Intermittent hypoxic training improves anaerobic performance in competitive swimmers when implemented into a direct competition mesocycle. PLoS ONE 2017, 12, e0180380. [Google Scholar] [CrossRef]
- Wang, R.; Guo, S.; Tian, H.; Huang, Y.; Yang, Q.; Zhao, K.; Kuo, C.-H.; Hong, S.; Chen, P.; Liu, T. Hypoxic Training in Obese Mice Improves Metabolic Disorder. Front. Endocrinol. 2019, 10, 527. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.; Larsen, B.; Cox, A.J.; Minahan, C. The Post-Exercise Inflammatory Response to Repeated-Sprint Running in Hypoxia. J. Sports Sci. Med. 2018, 17, 533–538. [Google Scholar]
- Naderi, M.; Kordestani, H.; Sahebi, Z.; Khedmati Zare, V.; Amani-Shalamzari, S.; Kaviani, M.; Wiskemann, J.; Molanouri Shamsi, M. Serum and gene expression profile of cytokines following combination of yoga training and vitamin D supplementation in breast cancer survivors: A randomized controlled trial. BMC Womens. Health 2022, 22, 90. [Google Scholar] [CrossRef]
- Timon, R.; Martínez-Guardado, I.; Camacho-Cardeñosa, A.; Villa-Andrada, J.M.; Olcina, G.; Camacho-Cardeñosa, M. Effect of intermittent hypoxic conditioning on inflammatory biomarkers in older adults. Exp. Gerontol. 2021, 152, 111478. [Google Scholar] [CrossRef]
- Kröpfl, J.M.; Kammerer, T.; Faihs, V.; Gruber, H.-J.; Stutz, J.; Rehm, M.; Stelzer, I.; Schäfer, S.T.; Spengler, C.M. Acute Exercise in Hypobaric Hypoxia Attenuates Endothelial Shedding in Subjects Unacclimatized to High Altitudes. Front. Physiol. 2020, 10, 1632. [Google Scholar] [CrossRef]
- Cheviron, Z.A.; Brumfield, R.T. Genomic insights into adaptation to high-altitude environments. Heredity 2012, 108, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Lomax, L. Hypoxic Training: The Effects, Adaptations and Benefits in Boxing. Available online: https://eis2win.co.uk/article/hypoxic-training-the-effects-adaptations-and-benefits-in-boxing/ (accessed on 22 March 2022).
- Liu, L.; Li, R.; Pan, Y.; Chen, J.; Li, Y.; Wu, J.; Zhu, Y. High-throughput screen of protein expression levels induced by cyclooxygenase-2 during influenza a virus infection. Clin. Chim. Acta 2011, 412, 1081–1085. [Google Scholar] [CrossRef]
- Carey, M.A.; Bradbury, J.A.; Rebolloso, Y.D.; Graves, J.P.; Zeldin, D.C.; Germolec, D.R. Pharmacologic Inhibition of COX-1 and COX-2 in Influenza A Viral Infection in Mice. PLoS ONE 2010, 5, e11610. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.Y.; Cheung, C.; Nicholls, J.M.; Hui, K.P.Y.; Leung, C.Y.H.; Uiprasertkul, M.; Tipoe, G.L.; Lau, Y.; Poon, L.L.M.; Ip, N.Y.; et al. Hyperinduction of Cyclooxygenase-2–Mediated Proinflammatory Cascade: A Mechanism for the Pathogenesis of Avian Influenza H5N1 Infection. J. Infect. Dis. 2008, 198, 525–535. [Google Scholar] [CrossRef]
- Da Silva, G.M.S.; Lima, L.M.; Fraga, C.A.M.; Sant’Anna, C.M.R.; Barreiro, E.J. The molecular basis for coxib inhibition of p38α MAP kinase. Bioorg. Med. Chem. Lett. 2005, 15, 3506–3509. [Google Scholar] [CrossRef]
- Kim, N.; Lannan, K.L.; Thatcher, T.H.; Pollock, S.J.; Woeller, C.F.; Phipps, R.P. Lipoxin B 4 Enhances Human Memory B Cell Antibody Production via Upregulating Cyclooxygenase-2 Expression. J. Immunol. 2018, 201, 3343–3351. [Google Scholar] [CrossRef]
- Carey, M.A.; Bradbury, J.A.; Seubert, J.M.; Langenbach, R.; Zeldin, D.C.; Germolec, D.R. Contrasting Effects of Cyclooxygenase-1 (COX-1) and COX-2 Deficiency on the Host Response to Influenza A Viral Infection. J. Immunol. 2005, 175, 6878–6884. [Google Scholar] [CrossRef] [Green Version]
- Jolly, M.K.; Ward, C.; Eapen, M.S.; Myers, S.; Hallgren, O.; Levine, H.; Sohal, S.S. Epithelial-mesenchymal transition, a spectrum of states: Role in lung development, homeostasis, and disease. Dev. Dyn. 2018, 247, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Rout-Pitt, N.; Farrow, N.; Parsons, D.; Donnelley, M. Epithelial mesenchymal transition (EMT): A universal process in lung diseases with implications for cystic fibrosis pathophysiology. Respir. Res. 2018, 19, 136. [Google Scholar] [CrossRef]
- Fabbrini, P.; Schilte, M.N.; Zareie, M.; ter Wee, P.M.; Keuning, E.D.; Beelen, R.H.J.; van den Born, J. Celecoxib treatment reduces peritoneal fibrosis and angiogenesis and prevents ultrafiltration failure in experimental peritoneal dialysis. Nephrol. Dial. Transplant. 2009, 24, 3669–3676. [Google Scholar] [CrossRef] [Green Version]
- Prasher, P.; Sharma, M.; Gunupuru, R. Targeting cyclooxygenase enzyme for the adjuvant COVID-19 therapy. Drug Dev. Res. 2021, 82, 469–473. [Google Scholar] [CrossRef]
- Simmons, C.; Farrar, J. Insights into Inflammation and Influenza. N. Engl. J. Med. 2008, 359, 1621–1623. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.-J.; Chan, K.-W.; Lin, Y.-P.; Zhao, G.-Y.; Chan, C.; Zhang, H.-J.; Chen, H.-L.; Wong, S.S.Y.; Lau, S.K.P.; Woo, P.C.Y.; et al. Delayed antiviral plus immunomodulator treatment still reduces mortality in mice infected by high inoculum of influenza A/H5N1 virus. Proc. Natl. Acad. Sci. USA 2008, 105, 8091–8096. [Google Scholar] [CrossRef] [Green Version]
- Gimeno, A.; Mestres-Truyol, J.; Ojeda-Montes, M.J.; Macip, G.; Saldivar-Espinoza, B.; Cereto-Massagué, A.; Pujadas, G.; Garcia-Vallvé, S. Prediction of Novel Inhibitors of the Main Protease (M-pro) of SARS-CoV-2 through Consensus Docking and Drug Reposition. Int. J. Mol. Sci. 2020, 21, 3793. [Google Scholar] [CrossRef]
- Ke, Y.-Y.; Peng, T.-T.; Yeh, T.-K.; Huang, W.-Z.; Chang, S.-E.; Wu, S.-H.; Hung, H.-C.; Hsu, T.-A.; Lee, S.-J.; Song, J.-S.; et al. Artificial intelligence approach fighting COVID-19 with repurposing drugs. Biomed. J. 2020, 43, 355–362. [Google Scholar] [CrossRef]
- Masclee, G.M.C.; Straatman, H.; Arfè, A.; Castellsague, J.; Garbe, E.; Herings, R.; Kollhorst, B.; Lucchi, S.; Perez-Gutthann, S.; Romio, S.; et al. Risk of acute myocardial infarction during use of individual NSAIDs: A nested case-control study from the SOS project. PLoS ONE 2018, 13, e0204746. [Google Scholar] [CrossRef]
- Qin, L.; Yao, Z.; Chang, Q.; Zhao, Y.; Liu, N.; Zhu, X.; Liu, Q.; Wang, L.; Yang, A.; Gao, C.; et al. Swimming attenuates inflammation, oxidative stress, and apoptosis in a rat model of dextran sulfate sodium-induced chronic colitis. Oncotarget 2017, 8, 7391–7404. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Feng, L.; Zhang, L.; Lu, Y.; Xu, J.; Lucia, A. Effects of Hypoxic Living and Training on Gene Expression in an Obese Rat Model. Med. Sci. Sport. Exerc. 2012, 44, 1013–1020. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supriya, R.; Singh, K.P.; Gao, Y.; Tao, D.; Cheour, S.; Dutheil, F.; Baker, J.S. Mimicking Gene–Environment Interaction of Higher Altitude Dwellers by Intermittent Hypoxia Training: COVID-19 Preventive Strategies. Biology 2023, 12, 6. https://doi.org/10.3390/biology12010006
Supriya R, Singh KP, Gao Y, Tao D, Cheour S, Dutheil F, Baker JS. Mimicking Gene–Environment Interaction of Higher Altitude Dwellers by Intermittent Hypoxia Training: COVID-19 Preventive Strategies. Biology. 2023; 12(1):6. https://doi.org/10.3390/biology12010006
Chicago/Turabian StyleSupriya, Rashmi, Kumar Purnendu Singh, Yang Gao, Dan Tao, Sarah Cheour, Frederic Dutheil, and Julien S. Baker. 2023. "Mimicking Gene–Environment Interaction of Higher Altitude Dwellers by Intermittent Hypoxia Training: COVID-19 Preventive Strategies" Biology 12, no. 1: 6. https://doi.org/10.3390/biology12010006