
Antiviral Properties of Pennisetum purpureum Extract against Coronaviruses and Enteroviruses
by
 , ,
, ,
Yi-Ning Chen
1,*,
Wenny Mei-Wen Kao
2,
Shu-Chi Lee
1,
Jaw-Min Wu
1,
Yi-Sheng Ho
1 and
Ming-Kun Hsieh
3 1
Department of Bioscience Technology, Chung Yuan Christian University, Taoyuan 320314, Taiwan
2
Institute of Plant Biology, National Taiwan University, Taipei 10617, Taiwan
3
Graduate Institute of Microbiology and Public Health, National Chung Hsing University, Taichung City 40227, Taiwan
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(11), 1371; https://doi.org/10.3390/pathogens11111371
Submission received: 17 October 2022
/
Revised: 16 November 2022
/
Accepted: 16 November 2022
/
Published: 17 November 2022
Abstract
:Many severe epidemics are caused by enteroviruses (EVs) and coronaviruses (CoVs), including feline coronavirus (FCoV) in cats, epidemic diarrhea disease virus (PEDV) in pigs, infectious bronchitis virus (IBV) in chickens, and EV71 in human. Vaccines and antiviral drugs are used to prevent and treat the infection of EVs and CoVs, but the effectiveness is affected due to rapidly changing RNA viruses. Many plant extracts have been proven to have antiviral properties despite the continuous mutations of viruses. Napier grass (Pennisetum purpureum) has high phenolic content and has been used as healthy food materials, livestock feed, biofuels, and more. This study tested the antiviral properties of P. purpureum extract against FCoV, PEDV, IBV, and EV71 by in vitro cytotoxicity assay, TCID50 virus infection assay, and chicken embryo infection assay. The findings showed that P. purpureum extract has the potential of being disinfectant to limit the spread of CoVs and EVs because the extract can inhibit the infection of EV71, FCoV, and PEDV in cells, and significantly reduce the severity of symptoms caused by IBV in chicken embryos.
1. Introduction
Coronaviruses (CoVs) and enteroviruses (EVs) have caused severe diseases with high morbidity and mortality in humans and animals [1,2,3,4]. The ongoing COVID-19 pandemic is the most recent outbreak of severe acute respiratory syndrome coronavirus (SARS-CoV)-2. As of August 2022, more than 500 million confirmed cases and 6 million deaths associated with COVID-19 have been confirmed worldwide, and in Taiwan, more than 4 million cases and 9000 deaths associated with COVID-19 have been confirmed. The first COVID-19 outbreak in Taiwan occurred in May 2021, and the second and most severe outbreak occurred in April 2022 [5]. Animal CoVs, such as feline CoV (FCoV), porcine epidemic diarrhea virus (PEDV), and infectious bronchitis virus (IBV), also cause severe diseases with high morbidity and mortality in cats, pigs, and chickens, respectively [6,7,8,9,10]. Feline infectious peritonitis (FIP), which is caused by a virulent strain of FCoV, can lead to high fatality and progressive multi-system disorders in young cat populations [6]. PEDV causes enteric diseases in pigs of all ages, and its clinical manifestations include acute watery diarrhea, dehydration, and vomiting; it leads to high mortality among nursey piglets, which causes tremendous losses in the swine industry worldwide [7,8]. IBV, which was discovered in the 1930s, causes respiratory, reproductive, and renal diseases in chickens of all ages, and has resulted in significant economic losses in the poultry industry [9,10]. Cases of EV infection occur throughout the year in Taiwan. Young children infected with EV may experience complications such as severe neurological diseases or death. The mortality rate is between 1.3% and 33.3%. In Taiwan, the major species in the genus Enterovirus most likely to be accompanied by severe complications is enterovirus 71 (EV71) [3,4].
CoVs have a positive-sense, single-stranded RNA genome that is tightly packed and has a nucleocapsid (N) protein at the center and an outer lipid envelope. Spike (S), membrane (M), and envelope (E) proteins are inserted into the outer lipid envelope. The life cycle of CoVs begins with the binding of the S proteins of CoVs to the receptors on the surfaces of susceptible cells and the subsequent fusion of the viral and cellular membranes. After the entry of CoVs, new viral RNA and proteins are transcribed, translated, and processed by cellular and viral enzymes, including RNA-dependent RNA polymerase (RdRp), 3-chymotrypsin-like protease (3CLpro), and papain-like protease (PLpro) [11]. EV71 is a nonenveloped virus with a positive-sense, single-stranded RNA genome enclosed within a pentameric icosahedral capsid, which encodes a polyprotein that is processed into structural proteins (VP1–VP4) and nonstructural proteins (2A–2C and 3A–3D) by viral proteases (2Apro, 3Cpro, and 3CDpro) [12]. CoVs and EVs are RNA viruses that mutate rapidly. Vaccines and antiviral drugs that are effective against a specific variant may be ineffective against other variants [7,13,14,15]. Herbal extracts are based on antiviral mechanisms different from those of vaccines and antiviral drugs and may be more effective against CoV s and EV constantly emerging variants; thus, herbal extracts are considered viable prophylactic and therapeutic options to reduce the severity of viral diseases [16,17,18]. In addition, the cost of using antiviral drugs in economic animals is too high, and the demand for adding them to feed and drinking water to improve the resistance of animals to CoVs has become increasingly important.
Napier grass Taishigrass (Pennisetum purpureum Schumach) is highly efficient in its use of water and nitrogen, and its conversion of light into biomass energy; in addition, it has a high tolerance to a variety of adverse soil conditions, including high salinity and waterlogging; these properties allow the grass to grow in the marginal zone of agriculture cultivation land and yield large quantities of biomass, even in environments with limited resources [19]. The Agriculture Committee of the Executive Yuan has promoted the cultivation of P. purpureum as an energy crop [20]. P. purpureum is also a food source and can be used in knitting, medicine, papermaking, and biofuel production [21,22]. P. purpureum is known for its health benefits and has been used in beverages and food production processing. Studies of feeding, toxicity, gene mutation, hematology, serum chemistry, and pathology in mice have demonstrated that P. purpureum is non-toxic. P. purpureum extract is rich in polyphenolic compounds, which can help to scavenge free radicals and inhibits peroxidation [23]. Polyphenolic compounds, which are promising inhibitors of viruses in herbal extracts, are small molecules with conjugated fused ring structures and are categorized into flavonoids (flavanols, catechins, anthocyanins) and non-flavonoids (phenolic acids, tannins, stilbenes) [24]. Dietary polyphenols display immunomodulatory capabilities involving inflammation control and immune responses [25]. In vitro experiments have shown that epigallocatechin (EGC), belonging to one of the catechins in the flavonoids of polyphenols in green tea, inhibits the infection of PEDV in Vero cells [26]. The addition of green tea byproducts to the feed and drinking water of chickens is associated with a significant antiviral effect against the H1N1 influenza virus [27]. P. purpureum, like green tea, contains a high level of polyphenols and may also have antiviral effects. However, P. purpureum can be grown without the use of pesticides and contains only a small amount of caffeine. In addition, as a poultry feed, P. purpureum is cheaper than green tea.
We investigated the antiviral properties of the pulverized extract of P. purpureum against FCoV, PEDV, IBV, and EV71 to determine whether it could act on both CoVs, which have a lipid envelope, and EVs, which do not have a lipid envelope. The findings may be useful in determining whether P. purpureum extract is useful in COVID-19 prevention as the disinfectant of drinking water, feed, and the environment for limiting the spread of viruses in the human disease control, company animal, poultry and swine industries.
2. Materials and Methods
2.1. Preparation of Pennisetum purpureum Extract (Heyiya®)
The anaerobic decomposition reaction method was performed at 600 °C to 700 °C for 1 h to prepare a pulverized crude extract from the fresh fibers separated from the stems and leaves of P. purpureum. The P. purpureum extract was prepared by HerbRay™ Biotech, Ltd. (Taipei City, Taiwan) and named Heyiya®. The pH of diluted extract (1×, 1/2×, 1/4×, 1/8×, 1/10×, 1/100×, 1/200×, 1/300×, and 1/600×) was measured by a pH-009(I) pen type meter (RongZhan).
2.2. Determination of Total Phenolic Content
Folin–Cicalteu’s (F-C) phenol reagent (Sigma-Aldrich, St. Louis, MO, USA) was used to determine the total phenolic content (TPC) of the P. purpureum extract in the dilution of 1×, 1/2×, 1/4×, 1/8×, 1/10×, 1/100×, 1/200×, 1/300×, and 1/600× according to the method described previously [28]. Briefly, an aliquot of 20 μL samples was incubated with 100 μL of F-C reagent and 80 μL of a 3% Na2CO3 solution for 20 min. The absorbance values of optical density (OD) were measured at 765 nm using BioTek Synergy multi-detection microplate reader (BioTek, Winoosk, VT, USA), and the concentrations of TPC, which were expressed as mg/L gallic acid equivalent (GAE), were determined by a calibration curve graph (R2 = 0.9993).
2.3. Cell Lines and Viruses
To test the antiviral properties of the P. purpureum extract on the enveloped CoVs and non-enveloped EVs, FCoV serotype II FIPV strain NTU156, PEDV strain Pingtung 52, IBV strain TW-2, and EV-A71 strain 2231 (TW/2231/98) are chosen. Felis catus whole fetus-4 (fcwf-4) cells for FCoV were maintained in Minimum Essential Media (MEM). Vero cells for PEDV and Rhabdomyosarcoma (RD) cells for EV71 were maintained in Dulbecco’s Modified Eagle Medium (DMEM). Both media were cultured with 10% fetal bovine serum (FBS), 100 IU/mL penicillin, and 100 IU/mL streptomycin solution in 5% CO2 at 37 °C. IBV was propagated in 10-day-old embryonated specific pathogen-free (SPE) chicken eggs (JD-SPF Biotech Co., Ltd., Miaoli, Taiwan) by inoculating the virus into the chorioallantoic sac of eggs. After the propagation and harvest, the extracellular viruses were collected by centrifuging at 500×g for 5 min and the intracellular viruses were released via three frozen-thawed cycles. The virus titer was determined with a 50% tissue culture infectious dose (TCID50) assay.
2.4. In Vitro Cytotoxicity Assay
Cytotoxicity of the P. purpureum extract was determined by using an MTS assay (CellTiter 96® AQueous One Solution Cell Proliferation Assay, Promega, Madison, WI, USA) for measuring the activity of cellular enzymes that reduce the tetrazolium dye to its insoluble formazan. The assays measured cellular metabolic activity via NAD(P)H-dependent cellular oxidoreductase enzymes and reflect the number of viable cells present. The extract was first adjusted to pH 7 using sterile 1N NaOH and then 10-fold serially diluted to incubate with Fcwf-4 cells or RD cells for 24 h at 37 °C. Three hours after the addition of lysis enzymes into the treated cells to release the color from the cells, OD at 500 nm was measured (BioTek, Winooski, VT, USA). All assays were performed in triplicates. Cell viability percentage was calculated as (1 − (ODtest-ODmedia)/(ODDMSO-ODmedia)) × 100%. Cells treated with 100% DMSO were used as the positive control (0% reference), and the cells inoculated with DMEM only served as the negative control (100% reference).
2.5. TCID50 Assay
RD cells, Fcwf-4 cells, and Vero cells were inoculated to 96-well plates in the concentration of 4 × 104 cells in 100 μL per well and reached a confluent monolayer after 24 h of incubation. Each virus specimen was serially ten-fold diluted into several dilutions. Each dilution was put into 8 wells, and in each well, 100 μL of the diluted virus solution was added. All tests were performed in duplicates. After 3 days of incubation at 37 °C with 5% CO2, the infectivity of viruses was determined by observing the formation of cytopathic effects (CPE) and the staining of viable cells by 0.5% crystal violet. The cells were fixed first with methanol for 30 min and then stained with 0.5% crystal violet in methanol for 15 min. The cells stained with 0.5% crystal violet were not infected with viruses, and the infected cells were washed away after the staining. The infectivity percentage was calculated as follows: infectivity% = (number of wells with virus-infected cells/number of wells with virus-inoculated cells) × 100%. The dilution of virus specimen made the cells in four out of eight wells infected is TCID50/100 μL, which was calculated as follows: ((infectivity% at the dilution immediately above 50%) – 50%)/(infectivity% at the dilution immediately above 50%) – (infectivity% at the dilution immediately below 50%)).
2.6. Virus Inhibition Assay
To determine the antiviral properties of the P. purpureum extract, 20 µL of FCoV, PEDV, or EV71 was pretreated with 50 µL of 10-fold, 100-fold, or 1000-fold diluted extract at room temperature for 1 h, 6 h or 12 h. After the designed incubation time, 950 μL of MEM or DMEM cell media was added to each well. Each reaction mixture was adjusted to pH 7 by using sterile 1N NaOH and serially 10-fold diluted and each dilution was put into 8 wells (100 μL/well). All tests were performed in duplicates or triplicates. After 3 days of incubation at 37 °C with 5% CO2, the infectivity of viruses was determined by TCID50/100 μL. Cells treated with viruses without pretreatment of P. purpureum extract were used as the positive control (PC). Relative TCID50/100 μL of the virus pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the virus without pretreatment of the extract) – log10 (TCID50/100 μL of virus pretreated with the extract). The inhibition efficacy (inhibition%) was calculated as follows: ((TCID50/100 μL of the virus without pretreatment of the extract) – (TCID50/100 μL of virus pretreated with the extract))/(TCID50/100 μL of the virus without pretreatment of the extract) × 100%.
2.7. Antiviral Effect of the P. purpureum Extract against IBV
One-day-old embryonated SPF chicken eggs (JD-SPF Biotech, Miaoli, Taiwan) were kept in an incubator at 38 °C for 9 days, and their viability was checked through candling before tests. To test the toxicity of the P. purpureum extract, the extract diluted in 1×, 1/10×, 1/100×, and 1/1000× was inoculated into 10-day-old embryonated eggs, and no concentrations of the extract were toxic to chicken embryos. Briefly, 20 μL of IBV was pretreated with 50 μL of 10-fold or 100-fold diluted extracts at room temperature for 1 h or 6 h, and then 950 μL of sterile PBS was added to stop the pretreatment. Three-to-five embryonated eggs were inoculated with 100 μL of each reaction mixture (1/10–1 h, 1/10–6 h, 1/100–1 h, 1/100–6 h). Eggs inoculated with IBV without the pretreatment of extract were the IBV control group and the embryos infected with IBV would show the typical symptoms of stunted growth, hemorrhage, or death. Eggs receiving sterile PBS were served as negative control (NC). After 3 days of incubation, all eggs were candled to check embryo viability and then chilled to 4 °C for the necropsy examination of the embryo’s body length and lesions.
2.8. Statistical Analysis
Comparisons of results between the two groups were analyzed using an unpaired t-test in the GraphPad Prism 8.0.1 program. The results are expressed as mean ± standard deviation (SD). The p-values of <0.05 were regarded as statistically significance, expressed as *, p < 0.05; **, p < 0.005; ***, p < 0.001; and ##, p < 0.005.
3. Results
3.1. Characterization of Pennisetum purpureum Extract
About 300 g of the pulverized crude extract can be acquired from 2 kg of fresh fiber separated from 10 kg of stems and leaves of P. purpureum. The extract is acidic and the pH ranged from 5.0 to 5.3. The mean and standard deviation values of TPC were 12,950 ± 755 mg/L GAE calculated in 100-fold to 600-fold dilutions (Table 1).
3.2. Cytotoxicity of Pennisetum purpureum Extract (Heyiya®)
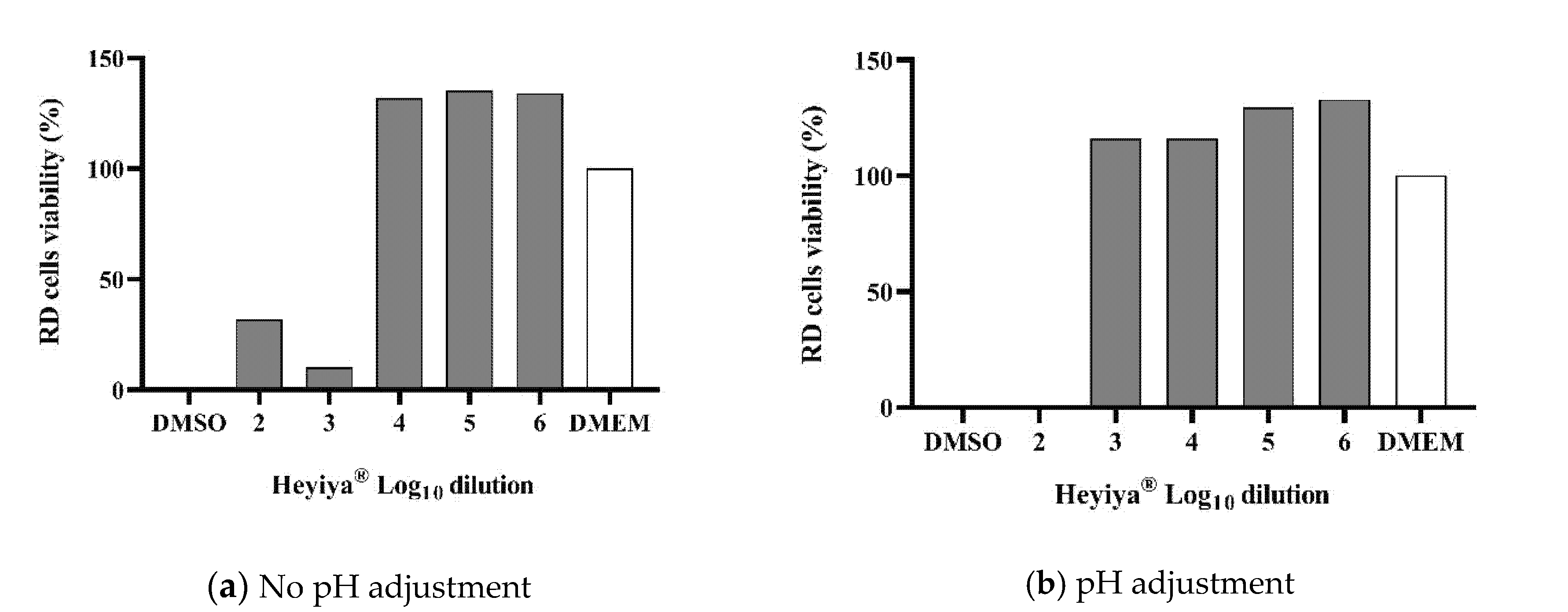
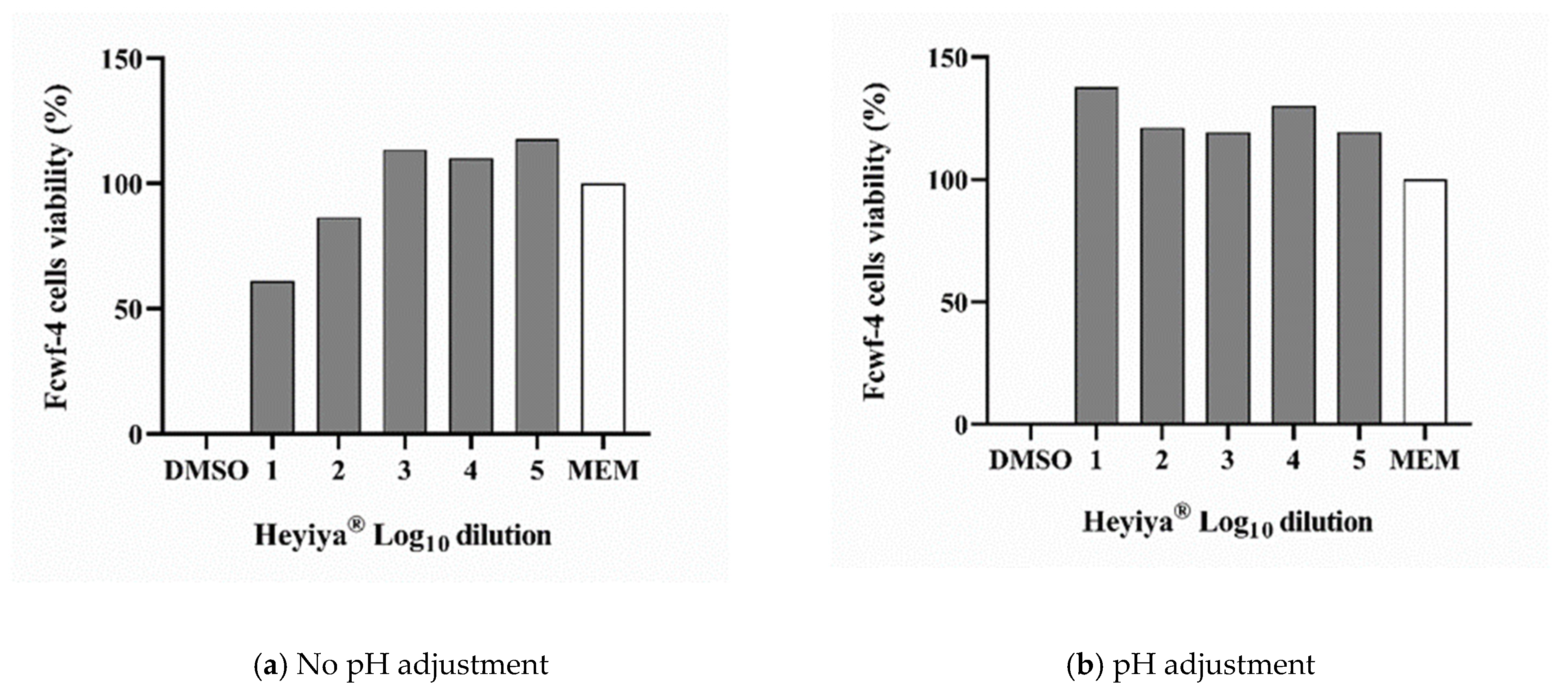
The toxicity of the P. purpureum extract to RD cells and Fcwf-4 cells, which were used to maintain EV71 and FCoV, respectively, was determined by MTS assay. Before pH adjustment, the 100-fold and 1000-fold dilutions of extract were toxic to RD cells and Fcwf-4 cells (Figure 1a and Figure 2a). The low pH of the extract contributed to its cytotoxicity. After the pH of the extract was adjusted to 7, cytotoxicity was observed only in RD cells treated with the 100-fold dilution of extract (Figure 1b and Figure 2b). Thus, the P. purpureum extract exhibited stronger cytotoxicity in RD cells than in Fcwf-4 cells.
3.3. Antiviral Effect of Pennisetum purpureum Extract (Heyiya®) against EV71
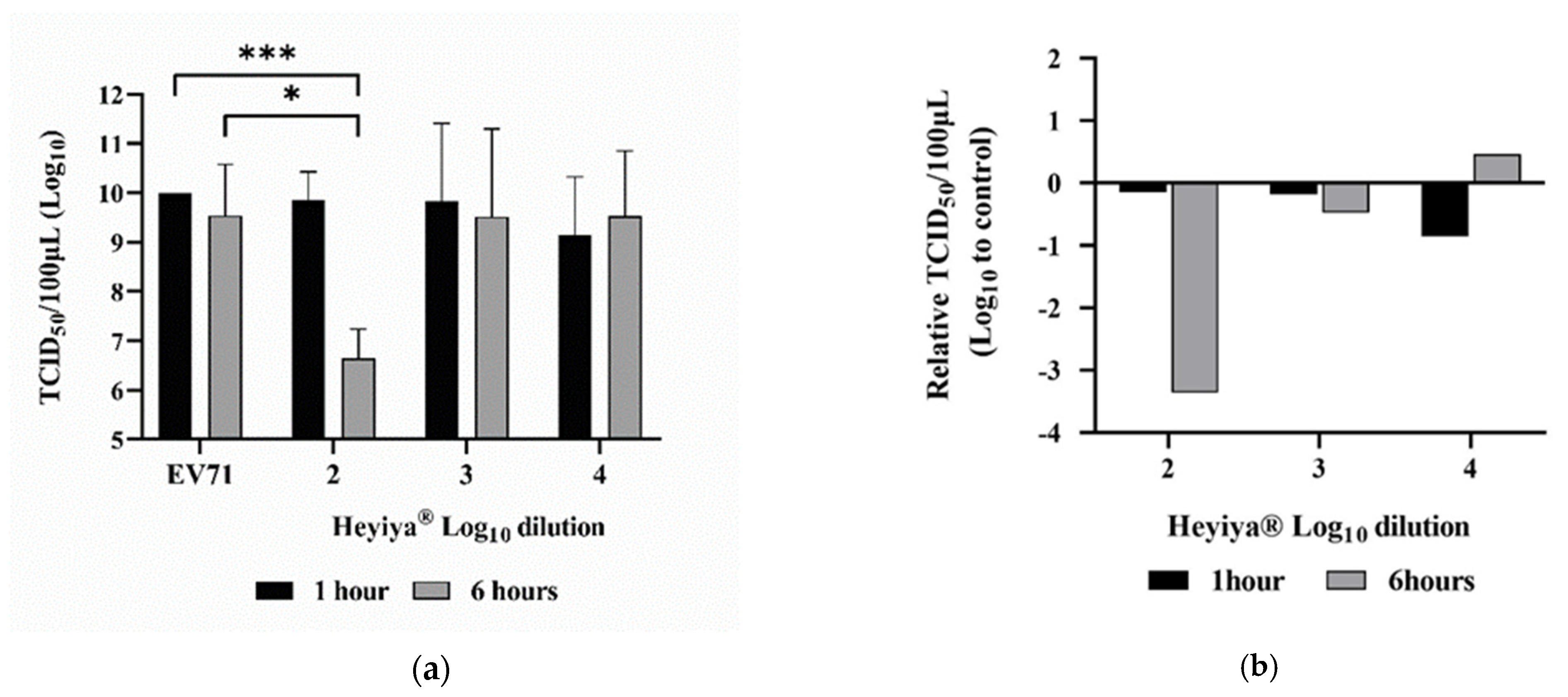
After pretreatment of EV71 with 10-fold serial dilutions (10−2–10−4, log10 dilution from 2 to 4) of P. purpureum extract for 1 or 6 h at room temperature, RD cells were inoculated with treated EV71 and the antiviral effect of the extract against EV71 was determined by TCID50 assay. The 100-fold dilution of P. purpureum extract with 6 h of incubation with EV71 achieved the highest virus inhibition rate, reaching 99.96% (***, p < 0.001) compared with no treatment of extract and the extract with 1 h of incubation (Figure 3a). Significantly reduced EV71 titer was observed in the EV71 pretreated with 100-fold diluted extract for 6 h compared with the EV71 without pretreatment of the extract (*, p < 0.05; Figure 3a). The relative TCID50/100 μL was calculated by subtracting Log10 TCID50/100 μL of the EV71 without the pretreatment of the extract from Log10 TCID50/100 μL of the EV71 pretreated with the extract. The largest reduction in viral titer was detected in the EV71 pretreated with 100-fold diluted extract for 6 h (Figure 3b). Individual values are presented in Table S1 in “Supplementary Materials”.
3.4. Antiviral Effect of Pennisetum purpureum Extract (Heyiya®) against FCoV
After pretreatment of FCoV with 10-fold serial dilutions (10−1–10−3, log10 dilution from 1 to 3) of P. purpureum extract for 1 or 6 h at room temperature, Fcwf-4 cells were inoculated with treated FCoV and the antiviral effect of the extract against FCoV was determined by TCID50 assay. As compared with the viral titer of FCoV without pretreatment of the extract, a significant reduction in viral titers was observed in the FCoV pretreated with 10-fold and 100-fold diluted extract for 1 h (**, p < 0.005) or 6 h (##, p < 0.005, Figure 4a, Table S1). The relative TCID50/100 μL was calculated by subtracting Log10 TCID50/100 μL of the FCoV without the pretreatment of the extract from Log10 TCID50/100 μL of the FCoV pretreated with the extract. A concentration-dependent reduction in the relative TCID50/100 μL was exhibited from the FCoV pretreated with the extract from 10-fold to the 1000-fold dilution (Figure 4b). Individual values are presented in Table S1 in “Supplementary Materials”.
3.5. Antiviral Effect of Pennisetum purpureum Extract (Heyiya®) against PEDV
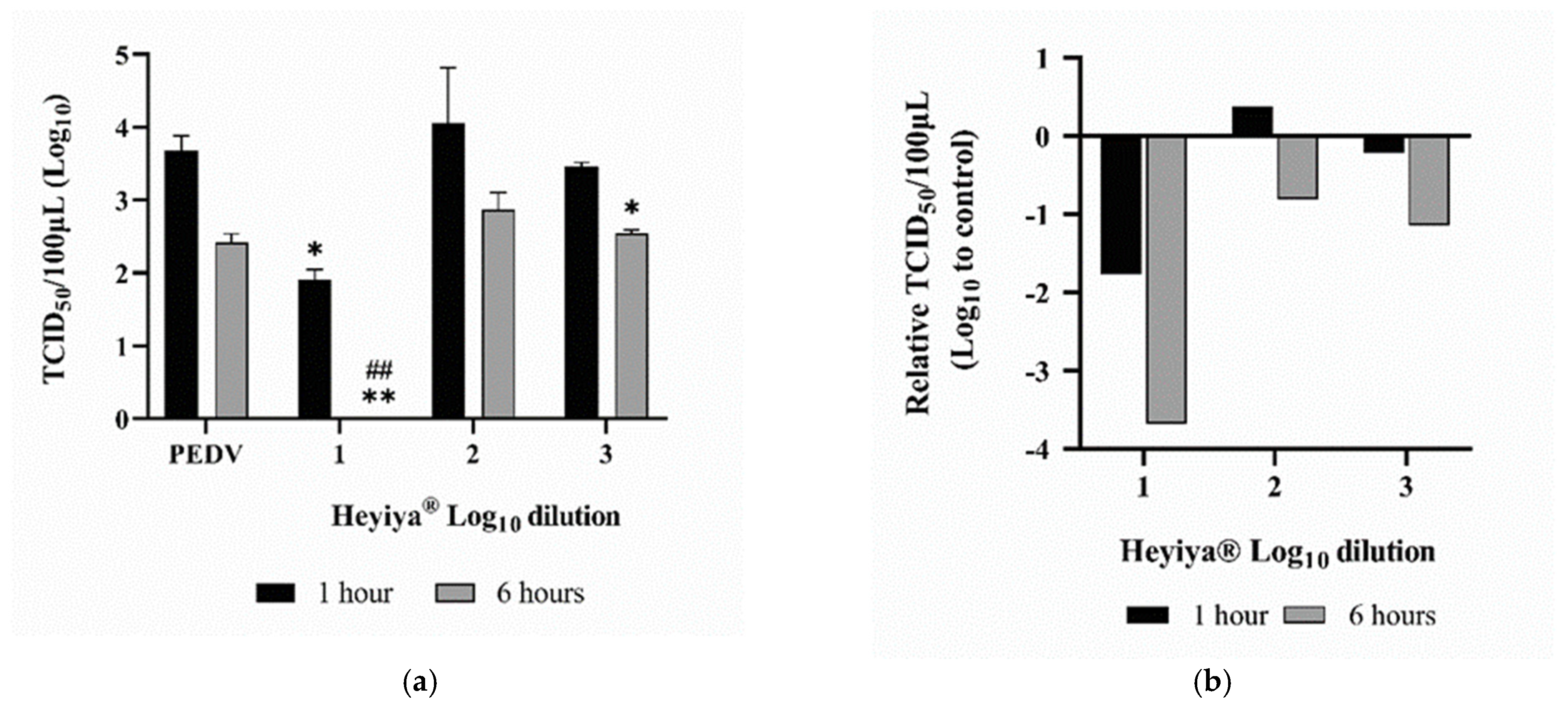
After pretreatment of PEDV with 10-fold serial dilutions (10−1–10−3, log10 dilution from 1 to 3) of P. purpureum extract for 1 or 6 h at room temperature, Vero cells were inoculated with treated PEDV and the antiviral effect of the extract against PEDV was determined by TCID50 assay. As shown in Figure 5, the infection of the PEDV pretreated with the 10-fold dilution of P. purpureum extract for 6 h was completely (100%) inhibited in VERO cells (p < 0.005). Furthermore, the infection of the PEDV pretreated with a 10-fold dilution of extract for 1 h was inhibited by 90% in VERO cells (p < 0.05). After 6 h of pretreatment with the 1000-fold dilution of extract, the viral titer of PEDV significantly decreased (p < 0.05). Individual values are presented in Table S1 in “Supplementary Materials”.
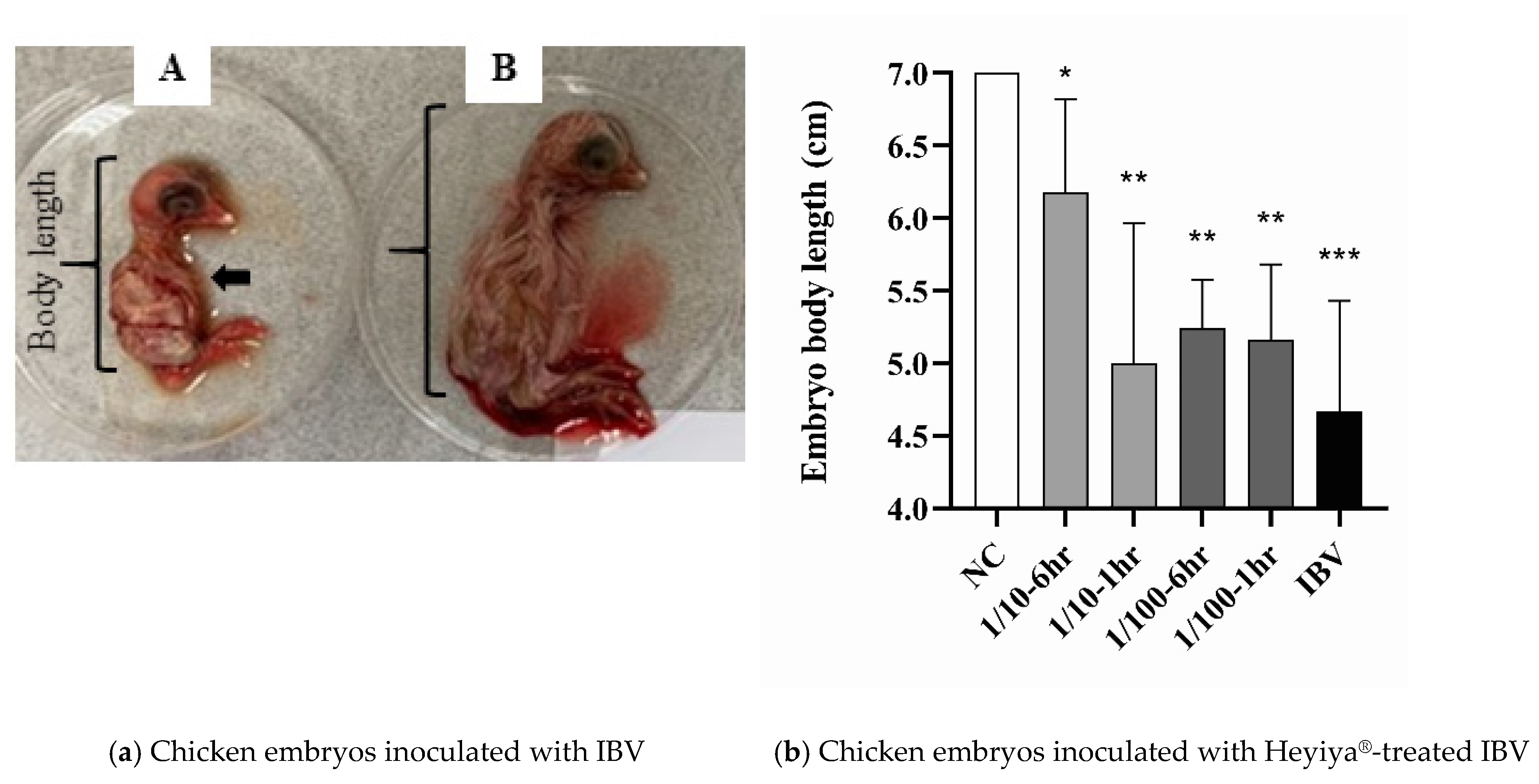
3.6. Antiviral Effect of Pennisetum purpureum Extract (Heyiya®) against IBV
After chicken eggs inoculated with IBV pretreated with 10-fold and 100-fold dilutions of P. alopecuroides extract for 1 or 6 h, the antiviral effect of the extract against the IBV TW2 strain was determined on the basis of the severity of clinical signs caused. The typical symptoms of IBV infection in 10-day-old chicken embryos included death, hemorrhage, and stunted growth (Figure 6a). The body lengths of the chicken embryos were measured to ascertain the severity of stunted growth. The pretreatment of P. purpureum extract can reduce the severity of IBV disease because the body lengths of the chicken embryos inoculated with IBV pretreated with the 10-fold or 100-fold dilution of P. purpureum extract for 1 h or 6 h were significantly greater than those of the embryos inoculated with IBV without pretreatment of the extract (Figure 6b). Individual values are presented in Table S2 in “Supplementary Materials”. The effects of P. purpureum extract were concentration and time-dependent because the chicken embryos inoculated with IBV pretreated with the 10-fold diluted extract for 6 h had the greatest body length in addition to the chicken embryo without the inoculation of IBV (NC).
4. Discussion
The use of disinfectants has become commonplace worldwide because of the COVID-19 pandemic. Alcohol and other chemical-based sanitizers recommended by the World Health Organization and United States Food and Drug Administration limit the spread of SARS-CoV-2, other CoVs, EVs, and many other viruses [29,30], but prolonged and excessive use of these sanitizers cause skin damage that facilitates the entry of harmful microbes [31,32]. Many plant-derived compounds have antimicrobial properties and are biodegradable and non-toxic; therefore, they may be useful as disinfectants [33]. Herbal extracts with antiviral properties can be added to feed and drinking water to reduce the spread of viruses as well as morbidity and disease severity [27]. In this study, pretreatment with the pulverized crude extract of P. purpureum at room temperature significantly reduced the infectivity of EV71, FCoV, and PEDV and the severity of symptoms caused by IBV in embryonated chickens, demonstrating the potential of the extract as an environmental disinfectant and sterilizing feed additive.
The most salient difference between the structures of CoVs and EVs is the outer membrane of the lipid envelope, which is derived from the cell membranes of previously infected cells. Alcohol-based disinfectants and many chemical-based disinfectants have amphiphilic properties that facilitate access to the viral lipid membrane through protein denaturation and disruption of the lipid envelope [33,34,35]. Such disinfectants are ineffective against EVs because EVs do not have a lipid envelope. However, P. purpureum extract was effective against both enveloped CoVs and non-enveloped EVs. The high TPC of the P. purpureum extract may explain its antiviral properties because polyphenols can use a variety of antiviral mechanisms in addition to disrupting the structure of the viral lipid envelope. According to the report on the website maintained by the Livestock Research Institute of the Agricultural Committee of the Executive Yuan (Taiwan), the P. purpureum extract contained 51 mg/g of flavonoids out of 120 mg/g of total phenols (42%) [21]. The phytochemical screening test conducted by the University of Port Harcourt, Nigeria, revealed that P. purpureum contained high levels of tannins, flavonoids, saponins, and alkaloids, known for their antimicrobial activities [36]. Many bioactive compounds categorized as flavonoids and non-flavonoids of polyphenols have been shown potent antiviral activity against CoVs and EVs in vitro, in vivo, and in silico findings [16,17,18], such as catechins of flavonoids against SARS-CoV [37] and PEDV [26], resveratrol (subclass stilbenes of non-flavonoids) against MERS-CoV [38] and EV71 [39], quercetin (subclass flavonols of flavonoids) against SARS-CoV-2 [40] and EV71 [41], and rosmarinic acid (subclass phenolic acids of non-flavonoids) as a pan-coronaviral main proteinase inhibitor [42] and an inhibitor against EV71 [43,44]. A recent study also showed that tannic acid-chelated zinc supplementation in the diet of newborn piglets could alleviate PEDV-induced damage of the intestinal mucosa and improve the absorptive function and growth in piglets [45]. Experimentally validated studies have found polyphenols can prevent the entry of viruses by reducing the levels of viral surface proteins to interfere with the binding of viruses with their cellular receptors [39,40,44,46]; interfere with the release of viral genomes into the host cells by inhibiting the production in viral enzymes associated with membrane fusion and viral uncoating [46,47,48,49] or interacting with viral proteins related to viral disassembly [41,43]; prevent viral replication and transcription by inhibiting the production of viral nucleocapsid proteins [37,38,50], and RNA polymerase [51,52]. Further studies are required to clarify the anti-viral mechanisms used by the P. purpureum extract to inhibit the infection of enveloped CoVs and non-enveloped EVs.
P. purpureum is the highest yielding forage species in Taiwan according to the Agriculture Committee of the Executive Yuan and is used as livestock feed, an alternative to coal and wood chip fuels, raw material for pulp, soil for mushroom cultivation, and energy biomass with high CO2 fixation capacity [22]. The Animal Testing Institute and Nuclear Research Institute have developed cellulosic alcohol with an alcohol–gasoline blend ratio of 3%, which can be used directly by most vehicles [20]. In addition, P. purpureum can be fermented with lactic acid bacteria to produce fibrous lactic acid, which can be used as industrial raw material and further synthesized into polylactic acid (PLA) particles for use in bioplastics. PLA is widely used in high-quality biomedical materials, packaging, films, fibers, plastic components of electronic goods, and 3D printing [20,22]. The P. purpureum extract (Heyiya®) tested in this study was obtained from the solid waste byproducts of such applications. Our results demonstrate the antiviral potential of P. purpureum extract (Heyiya®) and its applicability to the fields of animal husbandry, agriculture, veterinary medicine, and biomedicine for the control and prevention of the spread of viruses.
5. Conclusions
The infectivity of FCoV, PEDV, IBV, and EV71 significantly declined after the pretreatment of P. purpureum extract (Heyiya®) (Figure 7).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens11111371/s1, Table S1: Virus titers (TCID50/100 μL) of the viruses without pretreatment (EV71, FCoV, PEDV) or the viruses pretreated with 10-fold serial dilutions of the Pennisetum purpureum extract (Heyiya®) for 1 h and 6 h; Table S2: The body length (cm) of chicken embryo inoculated with infectious bronchitis virus (IBV) without the pretreatment of Pennisetum purpureum extract (Heyiya®), or with IBV pretreated with 10-fold or 100-fold diluted extract for 1 h or 6 h at room temperature, or without IBV (NC).
Author Contributions
Conceptualization, Y.-N.C. and W.M.-W.K.; methodology, Y.-N.C. and W.M.-W.K.; validation, S.-C.L., J.-M.W. and Y.-S.H.; formal analysis, S.-C.L., J.-M.W. and Y.-S.H.; resources, Y.-N.C., M.-K.H. and W.M.-W.K.; data curation, Y.-N.C., S.-C.L., J.-M.W. and Y.-S.H.; writing—original draft preparation, S.-C.L.; writing—review and editing, Y.-N.C.; visualization, S.-C.L.; supervision, Y.-N.C.; funding acquisition, Y.-N.C. and W.M.-W.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by HerbRay™ Biotech, Ltd. (Taipei City, Taiwan). The funding number in CYCU is 1092630.
Data Availability Statement
Not applicable.
Acknowledgments
The authors like to thank Hui-Wen Chen from the Institute of Veterinary Medicine at the National Taiwan University for providing the FCoV strain NTU156, Hui-Wen Chang from the Institute of Molecular and Comparative Pathogen at the National Taiwan University for providing PEDV strain Pingtung 52, Ming-Kun Hsieh at Graduate Institute of Microbiology and Public Health at National Chung Hsing University for providing IBV strain TW-2, and Jim-Tong Horng from Department of Biochemistry and Molecular Biology at Chang Gung University for providing EV71 strain 2231 (TW/2231/08).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Tang, G.; Liu, Z.; Chen, D. Human coronaviruses: Origin, host and receptor. J. Clin. Virol. 2022, 155, 105246. [Google Scholar] [CrossRef] [PubMed]
- Compton, S.R. Overview of coronaviruses in veterinary medicine. Comp. Med. 2021, 71, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Chen, E.R.; Hsu, K.H.; Twu, S.J.; Chen, K.T.; Tsai, S.F.; Wang, J.R.; Shih, S.R. An epidemic of enterovirus 71 infection in Taiwan. Taiwan Enterovirus Epidemic Working Group. N. Engl. J. Med. 1999, 341, 929–935. [Google Scholar] [CrossRef]
- Chang, P.C.; Chen, S.C.; Chen, K.T. The current status of the disease caused by enterovirus 71 infections: Epidemiology, pathogenesis, molecular epidemiology, and vac-cine development. Int. J. Environ. Res. Public. Health 2016, 13, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taiwan Centers for Disease Control. COVID-19. 2022. Available online: https://sites.google.com/cdc.gov.tw/2019ncov (accessed on 22 August 2022).
- Tekes, G.; Thiel, H.J. Feline coronaviruses: Pathogenesis of feline infectious peritonitis. Adv. Virus Res. 2016, 96, 193–218. [Google Scholar] [PubMed]
- Song, D.; Park, B. Porcine epidemic diarrhoea virus: A comprehensive review of molecular epidemiology, diagnosis, and vaccines. Virus Genes 2012, 44, 167–175. [Google Scholar] [CrossRef]
- Sung, M.H.; Lin, C.N.; Chiou, M.T.; Cheng, I.J.; Thanh, Q.H.; Chao, D.Y.; Lan, Y.C. Phylogeographic investigation of 2014 porcine epidemic diarrhea virus (PEDV) transmission in Taiwan. PLoS ONE 2019, 14, e0213153. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.T.; Tsai, H.F.; Wang, C.H. Detection of infectious bronchitis virus strains similar to Japan in Taiwan. J. Vet. Med. Sci. 2016, 78, 867–871. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.T.; Chen, T.C.; Lin, S.Y.; Mase, M.; Murakami, S.; Horimoto, T.; Chen, H.W. Emerging lethal infectious bronchitis coronavirus variants with multiorgan tropism. Transbound Emerg. Dis. 2020, 67, 884–893. [Google Scholar] [CrossRef] [Green Version]
- Masters, P.S. The molecular biology of coronaviruses. Adv. Virus Res. 2006, 65, 193–292. [Google Scholar]
- Solomon, T.; Lewthwaite, P.; Perera, D.J.; McMinn, P.; Solomon, T.; Lewthwaite, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef]
- Pedersen, N.C. An update on feline infectious peritonitis: Diagnostics and therapeutics. Vet. J. 2014, 201, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Wit, J.J.; Cook, J.K. Factors influencing the outcome of infectious bronchitis vaccination and challenge experiments. Avian. Pathol. 2014, 43, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, L.; Lu, J.; Kung, H.F.; He, M.L. The virology and developments towards control of human enterovirus 71. Crit. Rev. Microbiol. 2011, 37, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Remali, J.; Aizat, W.M. A review on plant bioactive compounds and their modes of action against coronavirus infection. Front Pharm. 2021, 11, 589044. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.P.G. Fighting coronaviruses with natural polyphenols. Biocatal. Agric. Biotechnol. 2021, 37, 102179. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, J.; Wang, L.; Ma, S.; Liu, Y. Anti-enterovirus 71 agents of natural products. Molecules 2015, 20, 16320–16333. [Google Scholar] [CrossRef] [Green Version]
- Council of Agriculture (COA). Brief introduction of Taiwanese Pennisetum Cultivated Varieties (Taishigrass). Agric. Policy Rev. 2015, 277. Available online: https://www.coa.gov.tw/us.php?id=2503170 (accessed on 30 July 2015). (In Chinese).
- Council of Agriculture (COA). Taiwan. Livestock Research Institute. Potential Energy Plants: Napiergrass Taishigrass No.4. 11 October 2017. Available online: https://kmweb.coa.gov.tw/theme_data.php?theme=technology_newa&id=6535 (accessed on 5 October 2021). (In Chinese)
- Council of Agriculture (COA). Taiwan. Research and Development of Pennisetum Food Materials. 21 September 2020. Available online: https://kmweb.coa.gov.tw/subject/subject.php?id=42050 (accessed on 5 October 2021). (In Chinese)
- Cheng, Y.K. Pennisetum breeding and diversified utilization. Sci. Dev. 2006, 407, 24–29. (In Chinese) [Google Scholar]
- Mathew, G.E.; Mathew, B.; Gokul, S.; Krishna, R.; Farisa, M.P. Determination of in vitro free radical scavenging and antiproliferative effect of Pennisetum alopecuroides on cultured A549 human lung cancer cells. Anc. Sci. Life 2015, 34, 175–178. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Somerville, V.S.; Braakhuis, A.J.; Hopkins, W.G. Effect of flavonoids on upper respiratory tract infections and immune function: A systematic review and meta-analysis. Adv. Nutr. 2016, 7, 488–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huan, C.; Xu, W.; Ni, B.; Guo, T.; Pan, H.; Jiang, L.; Li, L.; Yao, J.; Gao, S. Epigallocatechin-3-gallate, the main polyphenol in green tea, inhibits porcine epidemic diarrhea virus in vitro. Front. Pharm. 2021, 12, 628526. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, Y.N.; Youn, H.N.; Lee, D.H.; Kwak, J.H.; Seong, B.L.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Anti-influenza virus activity of green tea by-products in vitro and efficacy against influenza virus infection in chickens. Poult. Sci. 2012, 91, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Jouneghani, R.S.; Castro, A.H.F.; Panda, S.K.; Swennen, R.; Luyten, W. Antimicrobial activity of selected banana cultivars against important human pathogens, including Candida biofilm. Foods 2020, 9, 435. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Guide to Local Production: WHO-Recommended Handrub Formulations. 2020. Available online: https://www.who.int/gpsc/5may/Guide_to_Local_Production.pdf (accessed on 5 October 2021).
- Food and Drug Administration (FDA). Center for Drug Evaluation and Research. Policy for Temporary Compounding of Certain Alcohol-Based Hand Sanitizer Products during the Public Health Emergency. March 2020, Updated February 2021. Docket Number FDA-2020-D-1106. Available online: https://www.fda.gov/media/136118/download (accessed on 5 October 2021).
- Mahmood, A.; Eqan, M.; Pervez, S.; Alghamdi, H.A.; Tabinda, A.B.; Yasar, A.; Brindhadevi, K.; Pugazhendhi, A. COVID-19 and frequent use of hand sanitizers; human health and environmental hazards by exposure pathways. Sci. Total Environ. 2020, 742, 140561. [Google Scholar] [CrossRef]
- Jing, J.L.J.; Pei, Y.T.; Bose, R.J.C.; McCarthy, J.R.; Tharmalingam, N.; Madheswaran, T. Hand sanitizers: A review on formulation aspects, adverse effects, and regulations. Int. J. Environ. Res. Public Health 2020, 17, 3326. [Google Scholar] [CrossRef]
- Daverey, A.; Dutta, K. COVID-19: Eco-friendly hand hygiene for human and environ-mental safety. J. Environ. Chem. Eng. 2021, 9, 104754. [Google Scholar] [CrossRef]
- Lin, Q.; Lim, J.Y.C.; Xue, K.; Yew, P.Y.M.; Owh, C.; Chee, P.L.; Loh, X.J. Sanitizing agents for virus inactivation and disinfection. View 2020, 1, e16. [Google Scholar] [CrossRef]
- Singh, D.; Joshi, K.; Samuel, A.; Patra, J.; Mahindroo, N. Alcohol-based hand sanitizers as first line of defense against SARS-CoV-2: A review of biology, chemistry and formulations. Epidemiol. Infect. 2020, 148, e229. [Google Scholar] [CrossRef]
- Okaraonye, C.C.; Ikewuchi, J.C. Nutritional and antinutritional components of Pennisetum purpureum (Schumach). Pak. J. Nutr. 2009, 8, 32–34. [Google Scholar] [CrossRef] [Green Version]
- Roh, C. A facile inhibitor screening of SARS coronavirus N protein using nanoparticle-based RNA oligonucleotide. Int. J. Nanomed. 2012, 7, 2173–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.C.; Ho, C.T.; Chuo, W.H.; Li, S.; Wang, T.T.; Lin, C.C. Effective inhibition of MERS-CoV infection by resveratrol. BMC Infect Dis. 2017, 17, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, Y.; Gu, Z.; Wang, Y.; Shi, M.; Ji, Y.; Sun, J.; Xu, X.; Zhang, L.; Jiang, J.; et al. Resveratrol inhibits enterovirus 71 replication and pro-inflammatory cytokine secretion in rhabdosarcoma cells through blocking IKKs/NF-κB signaling pathway. PLoS ONE 2015, 10, e0116879. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.H.; Wu, K.L.; Zhang, X.; Deng, S.Q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef]
- Yao, C.; Xi, C.; Hu, K.; Gao, W.; Cai, X.; Qin, J.; Lv, S.; Du, C.; Wei, Y. Inhibition of enterovirus 71 replication and viral 3C protease by quercetin. Virol. J. 2018, 15, 116. [Google Scholar] [CrossRef] [Green Version]
- Shahhamzehei, N.; Abdelfatah, S.; Efferth, T. In silico and in vitro identification of pan-coronaviral main protease inhibitors from a large natural product library. Pharmaceuticals 2022, 15, 308. [Google Scholar] [CrossRef]
- Lin, W.Y.; Yu, Y.J.; Jinn, T.R. Evaluation of the virucidal effects of rosmarinic acid against enterovirus 71 infection via in vitro and in vivo study. Virol. J. 2019, 16, 94–103. [Google Scholar] [CrossRef]
- Hsieh, C.F.; Jheng, J.R.; Lin, G.H.; Chen, Y.L.; Ho, J.Y.; Liu, C.J.; Hsu, K.Y.; Chen, Y.S.; Chan, Y.F.; Yu, H.M.; et al. Rosmarinic acid exhibits broad anti-enterovirus A71 activity by inhibiting the interaction between the five-fold axis of capsid VP1 and cognate sulfated receptors. Emerg. Microbes. Infect. 2020, 9, 1194–1205. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Zheng, L.; Hou, Y.; Guo, S.; Wang, L.; Zhu, L.; Deng, C.; Wu, T.; Yi, D.; et al. Tannic acid-chelated zinc supplementation alleviates intestinal injury in piglets challenged by porcine epidemic diarrhea virus. Front Vet. Sci. 2022, 9, 1033022. [Google Scholar] [CrossRef]
- Poochi, S.P.; Easwaran, M.; Balasubramanian, B.; Anbuselvam, M.; Meyyazhagan, A.; Park, S.; Bhotla, H.K.; Anbuselvam, J.; Arumugam, V.A.; Keshavarao, S.; et al. Employing bioactive compounds derived from Ipomoea obscura (L.) to evaluate potential inhibitor for SARS-CoV-2 main protease and ACE2 protein. Food Front. 2020, 1, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.W.; Tsai, F.J.; Tsai, C.H.; Lai, C.C.; Wan, L.; Ho, T.Y.; Hsieh, C.C.; Chao, P.D.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Yuk, H.J.; Ryu, H.W.; Lim, S.H.; Kim, K.S.; Park, K.H.; Ryu, Y.B.; Lee, W.S. Evaluation of polyphenols from Broussonetia papyrifera as coronavirus protease inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Theerawatanasirikul, S.; Kuo, C.J.; Phetcharat, N.; Lekcharoensuk, P. In silico and in vitro analysis of small molecules and natural compounds targeting the 3CL protease of feline infectious peritonitis virus. Antivir. Res 2020, 174, 104697. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, J.; Zhang, Z.; Ma, Y.; Liao, F.; Zhang, Y.; Wu, G. Forsythoside a inhibits the avian infectious bronchitis virus in cell culture. Phytother. Res. 2011, 25, 338–342. [Google Scholar] [CrossRef]
- Lung, J.; Lin, Y.S.; Yang, Y.H.; Chou, Y.L.; Shu, L.H.; Cheng, Y.C. The potential chemical structure of anti-SARS-CoV-2 RNA-dependent RNA polymerase. J. Med. Virol. 2020, 92, 693–697. [Google Scholar] [CrossRef]
- Xu, L.; Su, W.; Jin, J.; Chen, J.; Li, X.; Zhang, X.; Sun, M.; Sun, S.; Fan, P.; An, D.; et al. Identification of luteolin as enterovirus 71 and coxsackievirus A16 inhibitors through reporter viruses and cell viability-based screening. Viruses 2014, 6, 2778–2795. [Google Scholar] [CrossRef]
Figure 1.
Cytotoxicity of Pennisetum purpureum extract (Heyiya®) in RD cells determined by MTS assay: (a) RD cells were treated with 10-fold serial dilutions of original extract (pH 5~5.3). (b) RD cells were treated with 10-fold serial dilutions of pH-adjusted extract (pH 7). The dilutions tested starting from 10−2 to 10−6, expressed as log10 dilution 2 to 6. All assays were performed in triplicate. Cell viability percentage was calculated as (1 − (ODtest − ODmedia)/(ODDMSO − ODmedia)) × 100%. Cells treated with 100% DMSO were used as the positive control (0% reference) and cells inoculated with medium DMEM only served as the negative control (100% reference).
Figure 1.
Cytotoxicity of Pennisetum purpureum extract (Heyiya®) in RD cells determined by MTS assay: (a) RD cells were treated with 10-fold serial dilutions of original extract (pH 5~5.3). (b) RD cells were treated with 10-fold serial dilutions of pH-adjusted extract (pH 7). The dilutions tested starting from 10−2 to 10−6, expressed as log10 dilution 2 to 6. All assays were performed in triplicate. Cell viability percentage was calculated as (1 − (ODtest − ODmedia)/(ODDMSO − ODmedia)) × 100%. Cells treated with 100% DMSO were used as the positive control (0% reference) and cells inoculated with medium DMEM only served as the negative control (100% reference).

Figure 2.
Cytotoxicity of Pennisetum purpureum extract (Heyiya®) in Fcwf-4 cells determined by MTS assay: (a) Fcwf-4 cells were treated with 10-fold serial dilutions of original extract (pH 5–5.3). (b) Fcwf-4 cells were treated with 10-fold serial dilutions of pH-adjusted extract (pH 7–7.2). The dilutions tested starting from 10−2 to 10−6, expressed as log10 dilution 2 to 6. All assays were performed in triplicate. Cell viability percentage was calculated as (1 − (ODtest − ODmedia)/(ODDMSO − ODmedia)) × 100%. Cells treated with 100% DMSO were used as the positive control (0% reference) and cells inoculated with medium MEM only were served as the negative control (100% reference).
Figure 2.
Cytotoxicity of Pennisetum purpureum extract (Heyiya®) in Fcwf-4 cells determined by MTS assay: (a) Fcwf-4 cells were treated with 10-fold serial dilutions of original extract (pH 5–5.3). (b) Fcwf-4 cells were treated with 10-fold serial dilutions of pH-adjusted extract (pH 7–7.2). The dilutions tested starting from 10−2 to 10−6, expressed as log10 dilution 2 to 6. All assays were performed in triplicate. Cell viability percentage was calculated as (1 − (ODtest − ODmedia)/(ODDMSO − ODmedia)) × 100%. Cells treated with 100% DMSO were used as the positive control (0% reference) and cells inoculated with medium MEM only were served as the negative control (100% reference).

Figure 3.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against enterovirus 71 (EV71): (a) Titers (TCID50/100 μL) of the EV71 pretreated with 10-fold serial dilutions of the extract starting from 10−2 to 10 −4 (log10 dilution from 2 to 4) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The EV71 without pretreatment of the extract was used as the positive control (EV71). Bars indicate means and error bars indicate standard deviation of triplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. The p-values of <0.05 were regarded as statistically significant, expressed as *, p < 0.05; ***, p < 0.001. (b) Relative TCID50/100 μL of EV71 pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the EV71 without pretreatment of the extract)-log10 (TCID50/100 μL of the EV71 pretreated with the extract), plotted against the log10 dilutions from 2 to 4 of the extract. The largest reduction in viral titer was detected in the EV71 pretreated with 100-fold diluted extract for 6 h.
Figure 3.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against enterovirus 71 (EV71): (a) Titers (TCID50/100 μL) of the EV71 pretreated with 10-fold serial dilutions of the extract starting from 10−2 to 10 −4 (log10 dilution from 2 to 4) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The EV71 without pretreatment of the extract was used as the positive control (EV71). Bars indicate means and error bars indicate standard deviation of triplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. The p-values of <0.05 were regarded as statistically significant, expressed as *, p < 0.05; ***, p < 0.001. (b) Relative TCID50/100 μL of EV71 pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the EV71 without pretreatment of the extract)-log10 (TCID50/100 μL of the EV71 pretreated with the extract), plotted against the log10 dilutions from 2 to 4 of the extract. The largest reduction in viral titer was detected in the EV71 pretreated with 100-fold diluted extract for 6 h.

Figure 4.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against feline coronavirus (FCoV): (a) Titers (TCID50/100 μL) of the FCoV pretreated with 10-fold serial dilutions of the extract starting from 10−1 to 10−3 (log10 dilution from 1 to 3) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The FCoV without pretreatment of the extract was used as the positive control (FCoV). Bars indicate means and error bars indicate standard deviation of triplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “**, p < 0.005” indicated the comparisons were between the FCoV without pretreatment of the extract and the FCoV pretreated with 10-fold or 100-fold of the extract for 6 h. “##, p < 0.005” indicated the comparisons were between the FCoV without pretreatment of the extract for 1 h and the FCoV pretreated with 10-fold or 100-fold of the extract for 6 h. (b) Relative TCID50/100 μL of FCoV pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the FCoV without pretreatment of the extract)-log10 (TCID50/100 μL of the FCoV pretreated with the extract), plotted against the log10 dilutions from 1 to 3 of the extract. The reduction in viral titer was in concentration-dependent manner.
Figure 4.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against feline coronavirus (FCoV): (a) Titers (TCID50/100 μL) of the FCoV pretreated with 10-fold serial dilutions of the extract starting from 10−1 to 10−3 (log10 dilution from 1 to 3) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The FCoV without pretreatment of the extract was used as the positive control (FCoV). Bars indicate means and error bars indicate standard deviation of triplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “**, p < 0.005” indicated the comparisons were between the FCoV without pretreatment of the extract and the FCoV pretreated with 10-fold or 100-fold of the extract for 6 h. “##, p < 0.005” indicated the comparisons were between the FCoV without pretreatment of the extract for 1 h and the FCoV pretreated with 10-fold or 100-fold of the extract for 6 h. (b) Relative TCID50/100 μL of FCoV pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the FCoV without pretreatment of the extract)-log10 (TCID50/100 μL of the FCoV pretreated with the extract), plotted against the log10 dilutions from 1 to 3 of the extract. The reduction in viral titer was in concentration-dependent manner.

Figure 5.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against porcine epidemic diarrhea virus (PEDV): (a) Titers (TCID50/100 μL) of the PEDV pretreated with 10-fold serial dilutions of the extract starting from 10 −1 to 10 −3 (log10 dilution from 1 to 3) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The PEDV without pretreatment of the extract was used as the positive control (PEDV). Bars indicate means and error bars indicate standard deviation of duplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “*, p < 0.05” indicated the comparisons were between the PEDV without pretreatment of the extract for 1 h and the PEDV pretreated with the 10-fold dilution of extract for 1 h, or the PEDV pretreated with the 1000-fold dilution of extract for 6 h. “**, p < 0.005” indicated the comparisons were between the PEDV without pretreatment of the extract for 1 h and the PEDV pretreated with 10-fold dilution of extract for 6 h. “##, p < 0.005” indicated the comparisons were between the PEDV without pretreatment of the extract for 6 h and the PEDV pretreated with 10-fold dilution of extract for 6 h. (b) Relative TCID50/100 μL of PEDV pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the PEDV without pretreatment of the extract)-log10 (TCID50/100 μL of the PEDV pretreated with the extract), plotted against the log10 dilutions from 1 to 3 of the extract.
Figure 5.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against porcine epidemic diarrhea virus (PEDV): (a) Titers (TCID50/100 μL) of the PEDV pretreated with 10-fold serial dilutions of the extract starting from 10 −1 to 10 −3 (log10 dilution from 1 to 3) for 1 h (black) and 6 h (grey) were determined by TCID50 assay. The PEDV without pretreatment of the extract was used as the positive control (PEDV). Bars indicate means and error bars indicate standard deviation of duplicate experimental data. Comparisons were performed through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “*, p < 0.05” indicated the comparisons were between the PEDV without pretreatment of the extract for 1 h and the PEDV pretreated with the 10-fold dilution of extract for 1 h, or the PEDV pretreated with the 1000-fold dilution of extract for 6 h. “**, p < 0.005” indicated the comparisons were between the PEDV without pretreatment of the extract for 1 h and the PEDV pretreated with 10-fold dilution of extract for 6 h. “##, p < 0.005” indicated the comparisons were between the PEDV without pretreatment of the extract for 6 h and the PEDV pretreated with 10-fold dilution of extract for 6 h. (b) Relative TCID50/100 μL of PEDV pretreated with the extract was calculated as follows: log10 (TCID50/100 μL of the PEDV without pretreatment of the extract)-log10 (TCID50/100 μL of the PEDV pretreated with the extract), plotted against the log10 dilutions from 1 to 3 of the extract.

Figure 6.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against infectious bronchitis virus (IBV): (a) Chicken embryos inoculated with IBV (A) had symptoms of hemorrhage (black arrow) and stunted growth with a lesser body length by comparing to the body length of chicken embryos without IBV inoculation (B). (b) The body lengths of chicken embryos without IBV inoculation (NC), inoculated with IBV without the pretreatment of P. purpureum extract (IBV), and inoculated with IBV pretreated with 10-fold or 100-fold diluted extract for 1 h or 6 h at room temperature (1/10–6 h, 1/10–1 h, 1/100–6 h, 1/100–1 h) were measured and compared through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “*, p < 0.05; **, p < 0.005; ***, p < 0.001” indicated the comparisons between NC to other groups, respectively.
Figure 6.
Antiviral effect of Pennisetum purpureum extract (Heyiya®) against infectious bronchitis virus (IBV): (a) Chicken embryos inoculated with IBV (A) had symptoms of hemorrhage (black arrow) and stunted growth with a lesser body length by comparing to the body length of chicken embryos without IBV inoculation (B). (b) The body lengths of chicken embryos without IBV inoculation (NC), inoculated with IBV without the pretreatment of P. purpureum extract (IBV), and inoculated with IBV pretreated with 10-fold or 100-fold diluted extract for 1 h or 6 h at room temperature (1/10–6 h, 1/10–1 h, 1/100–6 h, 1/100–1 h) were measured and compared through unpaired t-tests in GraphPad Prism 8.0.1. Statistical significance was indicated by a p-value of <0.05. “*, p < 0.05; **, p < 0.005; ***, p < 0.001” indicated the comparisons between NC to other groups, respectively.

Figure 7.
Illustration of antiviral properties of Pennisetum purpureum extract (Heyiya®) against coronaviruses and enterovirus.
Figure 7.
Illustration of antiviral properties of Pennisetum purpureum extract (Heyiya®) against coronaviruses and enterovirus.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The pH and total phenolic content of Pennisetum purpureum extract.
| Extract Dilution | pH | Total Phenolic Content (TPC) mg/L Gallic Acid Equivalent (GAE) |
|---|---|---|
| 1× | 5.3 | 1413.708 |
| 2× | 5.3 | 1328.513 |
| 4× | 5.3 | 1315.921 |
| 8× | 5.0 | 984.228 |
| 10× | 5.1 | 916.995 |
| 100× | 5.1 | 128.490 |
| 200× | 5.1 | 66.128 |
| 300× | 5.1 | 39.936 |
| 600× | 5.0 | 23.188 |
The concentrations of TPC were determined by a calibration curve (y = 2.3374 × + 0.0767, R2 = 0.9993) using Folin–Ciocalteu’s phenol reagent, expressed as mg/L GAE.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, Y.-N.; Kao, W.M.-W.; Lee, S.-C.; Wu, J.-M.; Ho, Y.-S.; Hsieh, M.-K. Antiviral Properties of Pennisetum purpureum Extract against Coronaviruses and Enteroviruses. Pathogens 2022, 11, 1371. https://doi.org/10.3390/pathogens11111371
AMA Style
Chen Y-N, Kao WM-W, Lee S-C, Wu J-M, Ho Y-S, Hsieh M-K. Antiviral Properties of Pennisetum purpureum Extract against Coronaviruses and Enteroviruses. Pathogens. 2022; 11(11):1371. https://doi.org/10.3390/pathogens11111371
Chicago/Turabian StyleChen, Yi-Ning, Wenny Mei-Wen Kao, Shu-Chi Lee, Jaw-Min Wu, Yi-Sheng Ho, and Ming-Kun Hsieh. 2022. "Antiviral Properties of Pennisetum purpureum Extract against Coronaviruses and Enteroviruses" Pathogens 11, no. 11: 1371. https://doi.org/10.3390/pathogens11111371
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.