
Single-Cell RNAseq Profiling of Human γδ T Lymphocytes in Virus-Related Cancers and COVID-19 Disease

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. scRNAseq Datasets Pre-Processing
2.2. Single-Cell Signatures and Scores
2.3. Score and Gate for Digital Extraction of γδ T Lymphocytes
- Positive selection of double negative cells (= T and NK lineages) from the scatterplot of ‘B cell’ and ‘Myeloid cell’ signature scores.
- Scatterplot of ‘B cell’ signature against the addition of ‘CD8AB’ and ‘TCRγδ’ signatures to positively select the double-positive cells (= non-CD4 T cells).
- Scatterplot of ‘CD8ab’ signature against ‘TCRα constant gene TRAC’ signature to digitally extract double-positive cells (= αβ T CD8 lymphocytes) on the one hand, and negative cells (= γδ T plus CD4CD8-double negative αβ T cells) on the other hand.
- Scatterplot of the addition of ‘CD3 complex’ and ‘TCRα constant gene TRAC’ signatures against the addition of ‘CD3 complex’ and ‘TCRγδ’ signatures to extract the ‘CD3 complex’ and ’TCRγδ’ double-positive cells (=γδ T lymphocytes).
2.4. Subtyping of the TCRVγ9 and TCRVγnon9 γδ T Cells
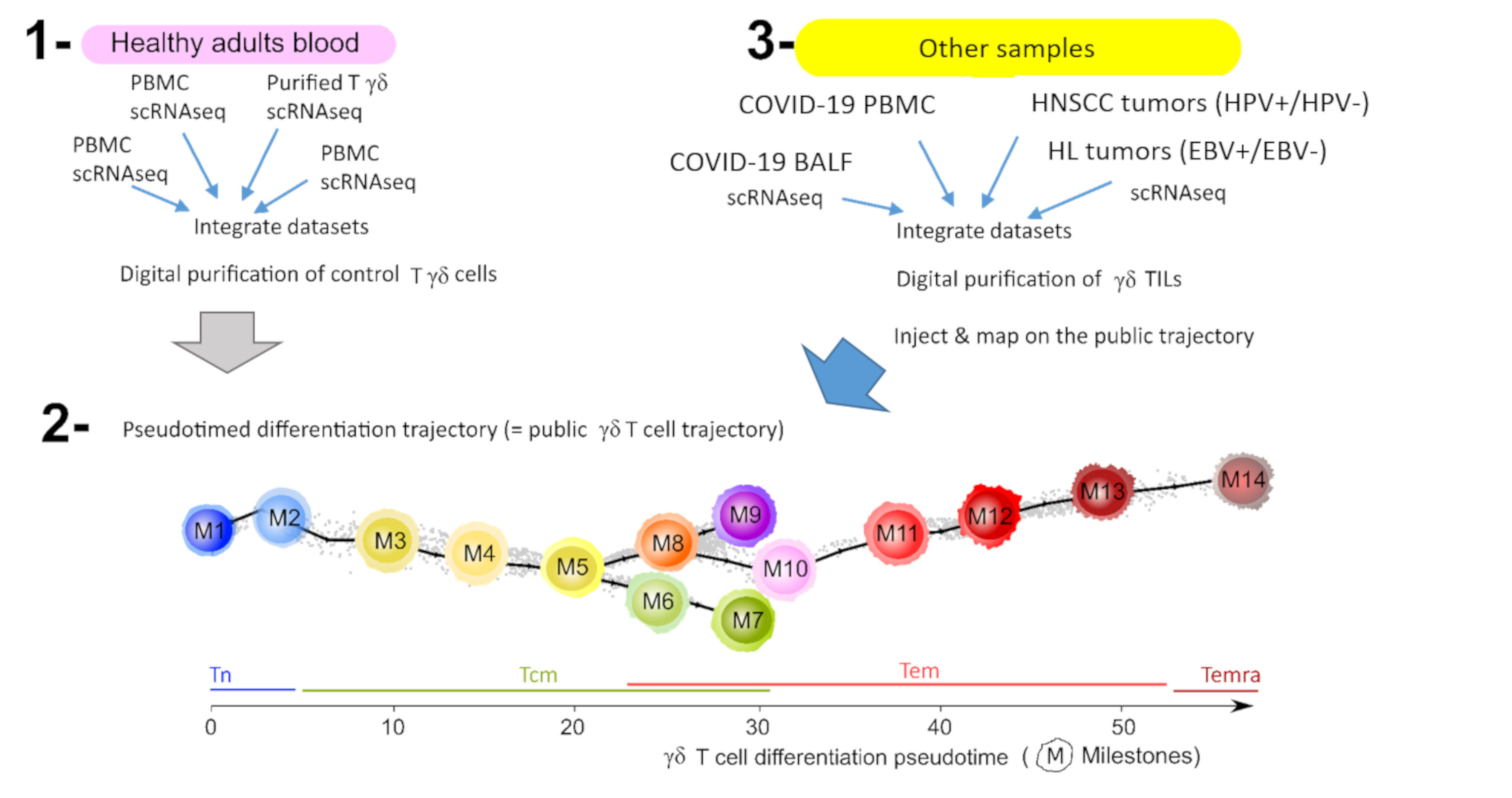
2.5. Cells Injection on Public Pseudotimed Differentiation Trajectory
- Query 1 = γδ T lymphocytes digitally extracted from COVID-19 samples;
- Reference = public trajectory;
- Integration of gene expression data from Query 1 and Reference (Seurat’s Integration).
- Query 2 = Integrated (Query and Reference);
- Reference = public trajectory;
- Transferring MST coordinates from Reference metadata to Query 2 (Seurat’s Map-to-Query).
- Query 2 = pseudotime calculation using MST coordinates and R package Dynverse.
- Visualization of new data (Query 1) on the public trajectory (Reference) using Single-Cell Multilayer Viewer [28].
2.6. Classifications
3. Results
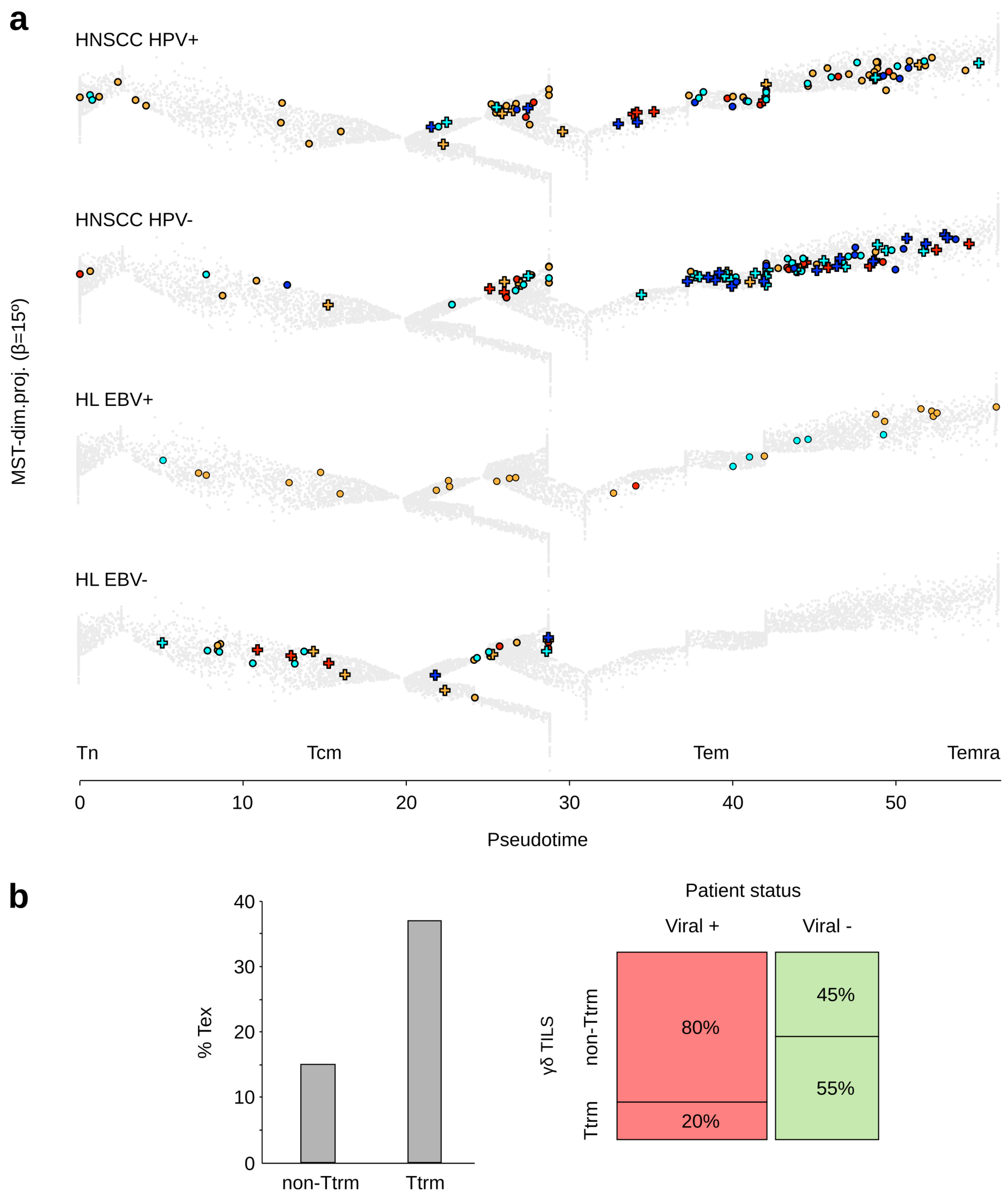
3.1. Tumor Infiltrating γδ T Lymphocytes from Virus-Positive and -Negative Cancer Patients
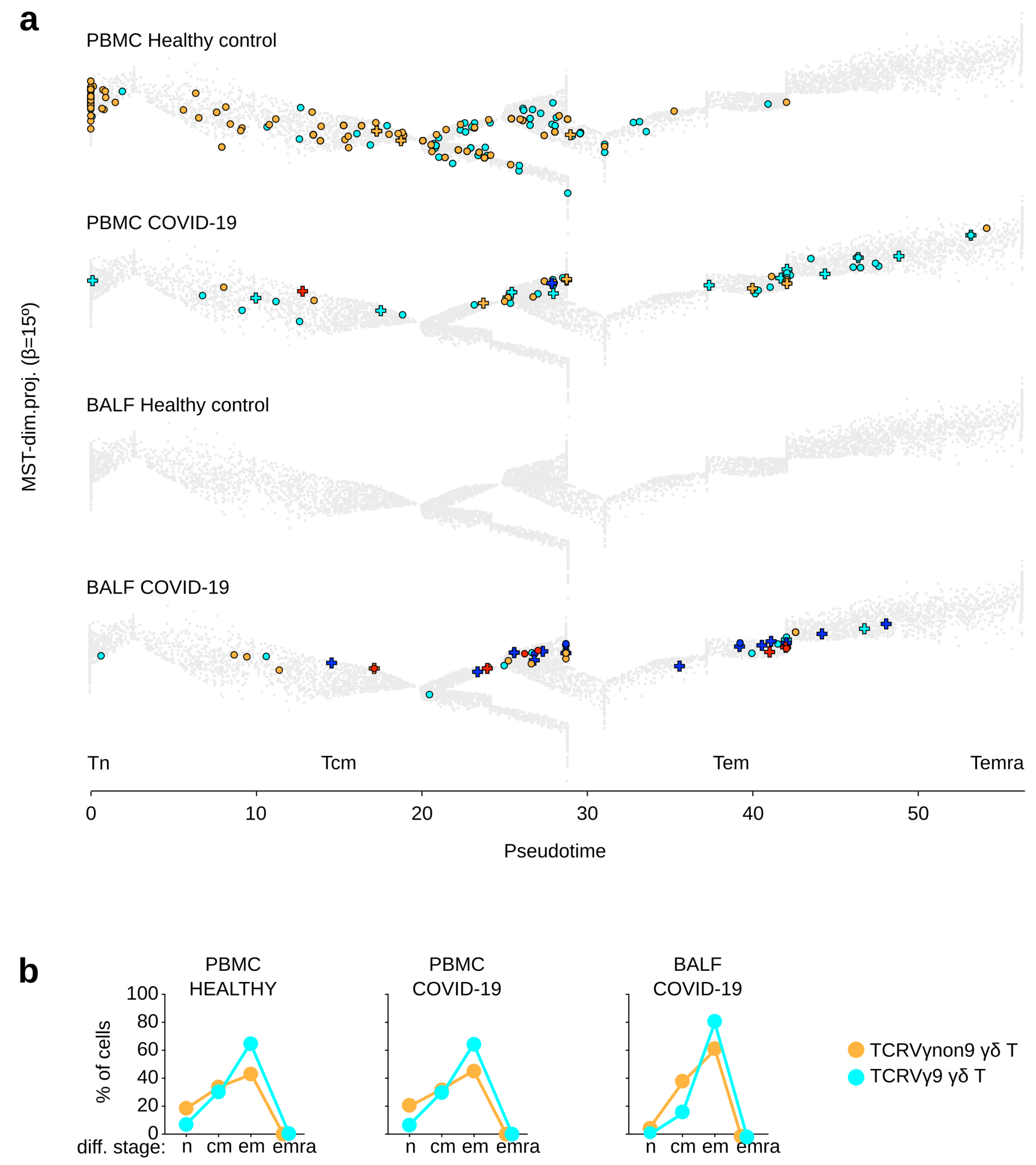
3.2. γδ T Lymphocytes from COVID-19 Patients
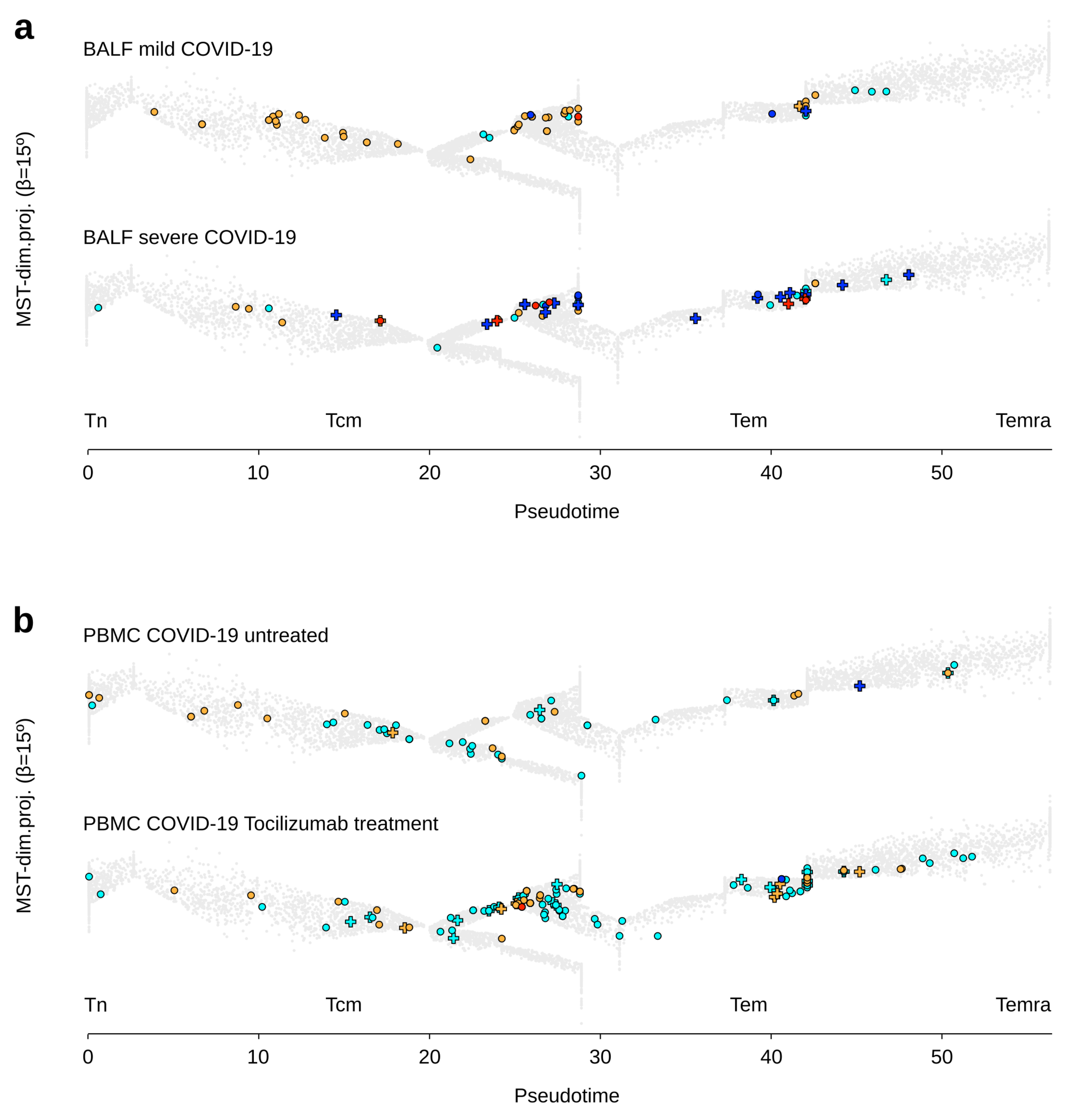
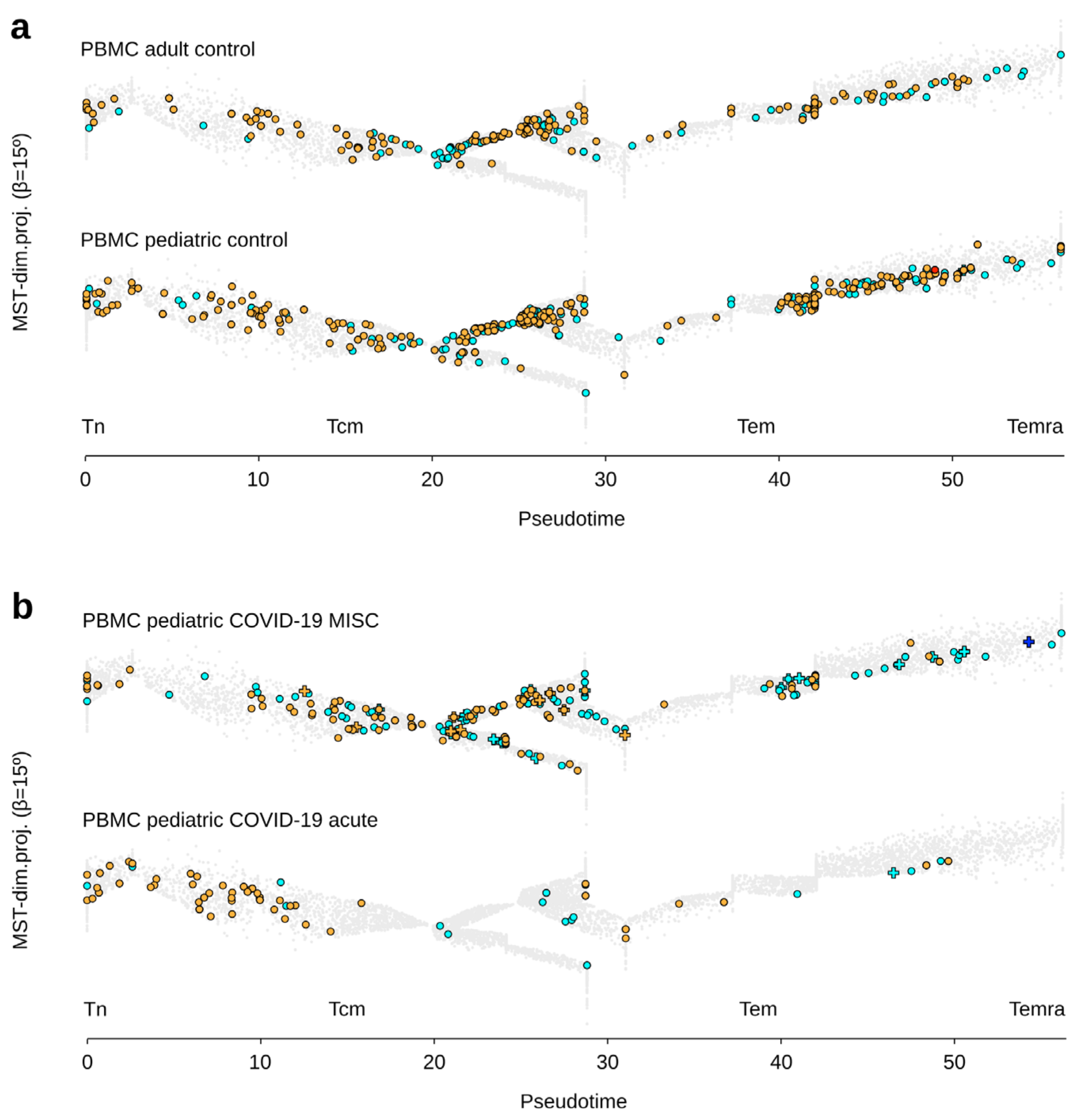
3.3. γδ T Lymphocytes in Clinical Groups of Adult Patients with COVID-19
3.4. γδ T Lymphocytes in Clinical Groups of Pediatric Patients with COVID-19
4. Discussion
Author Contributions
Funding
Institutional Review Board
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CMV | cytomegalovirus |
| COVID-19 | coronavirus disease 2019 |
| EBV | Epstein–Barr virus |
| HEV | hepatitis E virus |
| HL | Hodgkin’s lymphoma |
| HNSCC | head and neck squamous cell carcinoma |
| HPV | human papillomavirus |
| MIS-C | multisystem inflammatory syndrome in children |
| PBMC | peripheral blood mononuclear cells |
| SARS-CoV | severe acute respiratory syndrome-related coronavirus |
| scRNAseq | single-cell RNA sequencing |
| Tcm | central memory T |
| Tem | effector memory T |
| Temra | effector memory CD45RA T |
| Tex | exhausted T |
| TIL | tumor-infiltrated lymphocyte |
| Tn | naive T |
| Ttrm | tissue-resident memory T |
References
- Hayday, A.C.; Vantourout, P. The Innate Biologies of Adaptive Antigen Receptors. Annu. Rev. Immunol. 2020, 38, 487–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieli, F.; Poccia, F.; Lipp, M.; Sireci, G.; Caccamo, N.R.; DI Sano, C.; Salerno, A. Differentiation of Effector/Memory Vδ2 T Cells and Migratory Routes in Lymph Nodes or Inflammatory Sites. J. Exp. Med. 2003, 198, 391–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturm, E.; Braakman, E.; Fisch, P.; Vreugdenhil, R.J.; Sondel, P.; Bolhuis, R.L. Human V Gamma 9-V Delta 2 T Cell Receptor-Gamma Delta Lymphocytes Show Specificity to Daudi Burkitt’s Lymphoma Cells. J. Immunol. 1990, 145, 3202–3208. [Google Scholar] [PubMed]
- Sciammas, R.; Johnson, R.M.; I Sperling, A.; Brady, W.; Linsley, P.S.; Spear, P.G.; Fitch, F.W.; Bluestone, J. Unique Antigen Recognition by a Herpesvirus-Specific Tcr-Gamma Delta Cell. J. Immunol. 1994, 152, 5392–5397. [Google Scholar]
- Wallace, M.; Scharko, A.M.; Pauza, C.D.; Fisch, P.; Imaoka, K.; Kawabata, S.; Fujihashi, K.; Kiyono, H.; Tanaka, Y.; Bloom, B.R.; et al. Functional gamma delta T-lymphocyte defect associated with human immunodeficiency virus infections. Molecular Med. Eng. 1997, 3, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Poccia, F.; Gougeon, M.-; Agrati, C.; Montesano, C.; Martini, F.; Pauza, C.; Fisch, P.; Wallace, M.; Malkovsky, M. Innate T-Cell Immunity in HIV Infections: The Role of Vg9Vd2 T Lymphocytes. Curr. Mol. Med. 2002, 2, 769–781. [Google Scholar] [CrossRef]
- Hong, K.; Greer, C.; Ketter, N.; Van Nest, G.; Paliard, X. Isolation and characterization of human papillomavirus type 6-specific T cells infiltrating genital warts. J. Virol. 1997, 71, 6427–6432. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, S.; Ogasahara, K.; Kosaka, M.; Inoshita, T.; Wakatsuki, S.; Uehara, H.; Matsumoto, T. Hepatosplenic gamma delta T-cell lymphoma associated with hepatitis B virus infection. J. Med Investig. 1998, 44, 215–217. [Google Scholar]
- Qin, G.; Mao, H.; Zheng, J.; Sia, S.F.; Liu, Y.; Chan, P.L.; Lam, K.; Peiris, J.S.M.; Lau, Y.; Tu, W. Phosphoantigen-Expanded Human γδ T Cells Display Potent Cytotoxicity against Monocyte-Derived Macrophages Infected with Human and Avian Influenza Viruses. J. Infect. Dis. 2009, 200, 858–865. [Google Scholar] [CrossRef]
- Sant, S.; Jenkins, M.R.; Dash, P.; Watson, K.; Wang, Z.; Pizzolla, A.; Koutsakos, M.; Nguyen, T.H.; Lappas, M.; Crowe, J.; et al. Human γδ T-cell receptor repertoire is shaped by influenza viruses, age and tissue compartmentalisation. Clin. Transl. Immunol. 2019, 8, e1079. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-Y.; Liong, K.H.; Gunalan, M.G.; Liang, L.D.S.; Lim, D.S.L.; Fisher, D.A.; Macary, P.A.; Leo, Y.S.; Wong, S.-C.; Puan, K.J.; et al. Type I IFNs and IL-18 Regulate the Antiviral Response of Primary Human γδ T Cells against Dendritic Cells Infected with Dengue Virus. J. Immunol. 2015, 194, 3890–3900. [Google Scholar] [CrossRef]
- Poccia, F.; Agrati, C.; Castilletti, C.; Bordi, L.; Gioia, C.; Horejsh, D.; Ippolito, G.; Chan, P.; Hui, D.; Sung, J.J.Y.; et al. Anti–Severe Acute Respiratory Syndrome Coronavirus Immune Responses: The Role Played by Vγ9Vδ2 T Cells. J. Infect. Dis. 2006, 193, 1244–1249. [Google Scholar] [CrossRef]
- Déchanet, J.; Merville, P.; Lim, A.; Retière, C.; Pitard, V.; Lafarge, X.; Michelson, S.; Méric, C.; Hallet, M.-M.; Kourilsky, P.; et al. Implication of γδ T cells in the human immune response to cytomegalovirus. J. Clin. Investig. 1999, 103, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Barragué, H.; Fontaine, J.; Abravanel, F.; Mauré, E.; Péron, J.-M.; Alric, L.; Dubois, M.; Izopet, J.; Champagne, E. Mobilization of γδ T Cells and IL-10 Production at the Acute Phase of Hepatitis E Virus Infection in Cytomegalovirus Carriers. J. Immunol. 2021, 206, 1027–1038. [Google Scholar] [CrossRef]
- Barcy, S.; De Rosa, S.C.; Vieira, J.; Diem, K.; Ikoma, M.; Casper, C.; Corey, L. γδ+T Cells Involvement in Viral Immune Control of Chronic Human Herpesvirus 8 Infection. J. Immunol. 2008, 180, 3417–3425. [Google Scholar] [CrossRef] [Green Version]
- Rijkers, G.; Vervenne, T.; Van Der Pol, P. More bricks in the wall against SARS-CoV-2 infection: Involvement of γ9δ2 T cells. Cell. Mol. Immunol. 2020, 17, 771–772. [Google Scholar] [CrossRef]
- Carissimo, G.; Xu, W.; Kwok, I.; Abdad, M.Y.; Chan, Y.-H.; Fong, S.-W.; Puan, K.J.; Lee, C.Y.-P.; Yeo, N.K.-W.; Amrun, S.N.; et al. Whole blood immunophenotyping uncovers immature neutrophil-to-VD2 T-cell ratio as an early marker for severe COVID-19. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Jouan, Y.; Guillon, A.; Gonzalez, L.; Perez, Y.; Boisseau, C.; Ehrmann, S.; Ferreira, M.; Daix, T.; Jeannet, R.; François, B.; et al. Phenotypical and functional alteration of unconventional T cells in severe COVID-19 patients. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef]
- Li, C.-X.; Chen, J.; Lv, S.-K.; Li, J.-H.; Li, L.-L.; Hu, X. Whole-Transcriptome RNA Sequencing Reveals Significant Differentially Expressed mRNAs, miRNAs, and lncRNAs and Related Regulating Biological Pathways in the Peripheral Blood of COVID-19 Patients. Mediat. Inflamm. 2021, 2021, 1–22. [Google Scholar] [CrossRef]
- Kalicińska, E.; Szymczak, D.; Zińczuk, A.; Adamik, B.; Smiechowicz, J.; Skalec, T.; Nowicka-Suszko, D.; Biernat, M.; Bogucka-Fedorczuk, A.; Rybka, J.; et al. Immunosuppression as a Hallmark of Critical COVID-19: Prospective Study. Cells 2021, 10, 1293. [Google Scholar] [CrossRef]
- Shah, A.A.; Casciola-Rosen, L.; Rosen, A. Review: Cancer-Induced Autoimmunity in the Rheumatic Diseases. Arthritis Rheumatol. 2015, 67, 317–326. [Google Scholar] [CrossRef]
- Meraviglia, S.; Presti, E.L.; Tosolini, M.; La Mendola, C.; Orlando, V.; Todaro, M.; Catalano, V.; Stassi, G.; Cicero, G.; Vieni, S.; et al. Distinctive features of tumor-infiltrating γδ T lymphocytes in human colorectal cancer. OncoImmunology 2017, 6, e1347742. [Google Scholar] [CrossRef] [Green Version]
- Gentles, A.J.; Newman, A.; Liu, C.L.; Bratman, S.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef]
- Wu, Y.; Kyle-Cezar, F.; Woolf, R.T.; Naceur-Lombardelli, C.; Owen, J.; Biswas, D.; Lorenc, A.; Vantourout, P.; Gazinska, P.; Grigoriadis, A.; et al. An innate-like Vδ1 + γδ T cell compartment in the human breast is associated with remission in triple-negative breast cancer. Sci. Transl. Med. 2019, 11, eaax9364. [Google Scholar] [CrossRef]
- Mikulak, J.; Oriolo, F.; Bruni, E.; Roberto, A.; Colombo, F.S.; Villa, A.; Bosticardo, M.; Bortolomai, I.; Presti, E.L.; Meraviglia, S.; et al. NKp46-expressing human gut-resident intraepithelial Vδ1 T cell subpopulation exhibits high antitumor activity against colorectal cancer. JCI Insight 2019, 4, 125884. [Google Scholar] [CrossRef] [Green Version]
- Foord, E.; Arruda, L.C.M.; Gaballa, A.; Klynning, C.; Uhlin, M. Characterization of ascites- and tumor-infiltrating γδ T cells reveals distinct repertoires and a beneficial role in ovarian cancer. Sci. Transl. Med. 2021, 13, 0192. [Google Scholar] [CrossRef]
- Pizzolato, G.; Kaminski, H.; Tosolini, M.; Franchini, D.-M.; Pont, F.; Martins, F.; Valle, C.; Labourdette, D.; Cadot, S.; Quillet-Mary, A.; et al. Single-cell RNA sequencing unveils the shared and the distinct cytotoxic hallmarks of human TCRVδ1 and TCRVδ2 γδ T lymphocytes. Proc. Natl. Acad. Sci. USA 2019, 116, 11906–11915. [Google Scholar] [CrossRef] [Green Version]
- Cerapio, J.-P.; Perrier, M.; Balança, C.-C.; Gravelle, P.; Pont, F.; Devaud, C.; Franchini, D.-M.; Féliu, V.; Tosolini, M.; Valle, C.; et al. Phased differentiation of γδ T and T CD8 tumor-infiltrating lymphocytes revealed by single-cell transcriptomics of human cancers. OncoImmunology 2021, 10, 1939518. [Google Scholar] [CrossRef]
- Aoki, T.; Chong, L.C.; Takata, K.; Milne, K.; Hav, M.; Colombo, A.; Chavez, E.A.; Nissen, M.; Wang, X.; Miyata-Takata, T.; et al. Single-Cell Transcriptome Analysis Reveals Disease-Defining T-cell Subsets in the Tumor Microenvironment of Classic Hodgkin Lymphoma. Cancer Discov. 2019, 10, 406–421. [Google Scholar] [CrossRef] [Green Version]
- Cillo, A.R.; Kürten, C.H.; Tabib, T.; Qi, Z.; Onkar, S.; Wang, T.; Liu, A.; Duvvuri, U.; Kim, S.; Soose, R.J.; et al. Immune Landscape of Viral- and Carcinogen-Driven Head and Neck Cancer. Immuni 2020, 52, 183–199.e21. [Google Scholar] [CrossRef]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Papalexi, E.; Mauck, W.M., III; Hao, Y.; Stoeckius, M.; Smibert, P.; Satija, R. Comprehensive Integration of Single-Cell Data. Cell 2019, 177, 1888–1902. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.; Healy, J. UMAP: Uniform Manifold Approximation and Projection for Dimension Reduction. Available online: https://arxiv.org/abs/1802.03426 (accessed on 12 October 2021).
- Pont, F.; Tosolini, M.; Fournié, J.J. Single-Cell Signature Explorer for comprehensive visualization of single cell signatures across scRNA-seq datasets. Nucleic Acids Res. 2019, 47, e133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pont, F.; Tosolini, M.; Gao, Q.; Perrier, M.; Madrid-Mencía, M.; Huang, T.S.; Neuvial, P.; Ayyoub, M.; Nazor, K.; Fournié, J.-J. Single-Cell Virtual Cytometer allows user-friendly and versatile analysis and visualization of multimodal single cell RNAseq datasets. NAR Genom. Bioinform. 2020, 2, lqaa025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chihara, N.; Madi, A.; Kondo, T.; Zhang, H.; Acharya, N.; Singer, M.; Nyman, J.; Marjanovic, N.D.; Kowalczyk, M.S.; Wang, C.; et al. Induction and transcriptional regulation of the co-inhibitory gene module in T cells. Nat. Cell Biol. 2018, 558, 454–459. [Google Scholar] [CrossRef] [Green Version]
- Alfei, F.; Kanev, K.; Hofmann, M.; Wu, M.; Ghoneim, H.E.; Roelli, P.; Utzschneider, D.T.; Von Hoesslin, M.; Cullen, J.G.; Fan, Y.; et al. TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 2019, 571, 265–269. [Google Scholar] [CrossRef]
- Khan, O.; Giles, J.R.; McDonald, S.; Manne, S.; Ngiow, S.F.; Patel, K.P.; Werner, M.T.; Huang, A.C.; Alexander, K.A.; Wu, J.E.; et al. TOX transcriptionally and epigenetically programs CD8+ T cell exhaustion. Nature 2019, 571, 211–218. [Google Scholar] [CrossRef]
- Tosolini, M.; Algans, C.; Pont, F.; Ycart, B.; Fournié, J.-J. Large-scale microarray profiling reveals four stages of immune escape in non-Hodgkin lymphomas. OncoImmunology 2016, 5, e1188246. [Google Scholar] [CrossRef] [Green Version]
- Balança, C.-C.; Scarlata, C.-M.; Michelas, M.; Devaud, C.; Sarradin, V.; Franchet, C.; Gomez, C.M.; Gomez-Roca, C.; Tosolini, M.; Heaugwane, D.; et al. Dual Relief of T-lymphocyte Proliferation and Effector Function Underlies Response to PD-1 Blockade in Epithelial Malignancies. Cancer Immunol. Res. 2020, 8, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.V.; Ma, W.; Miron, M.; Granot, T.; Guyer, R.S.; Carpenter, D.J.; Senda, T.; Sun, X.; Ho, S.-H.; Lerner, H.; et al. Human Tissue-Resident Memory T Cells Are Defined by Core Transcriptional and Functional Signatures in Lymphoid and Mucosal Sites. Cell Rep. 2017, 20, 2921–2934. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.D.; Madireddi, S.; de Almeida, P.E.; Banchereau, R.; Chen, Y.-J.J.; Chitre, A.S.; Chiang, E.Y.; Iftikhar, H.; O’Gorman, W.E.; Au-Yeung, A.; et al. Peripheral T cell expansion predicts tumour infiltration and clinical response. Nat. Cell Biol. 2020, 579, 274–278. [Google Scholar] [CrossRef]
- Alpert, A.; Moore, L.S.; Dubovik, T.; Shen-Orr, S.S. Alignment of single-cell trajectories to compare cellular expression dynamics. Nat. Methods 2018, 15, 267–270. [Google Scholar] [CrossRef]
- Trapnell, C.; Cacchiarelli, D.; Grimsby, J.; Pokharel, P.; Li, S.; Morse, M.; Lennon, N.J.; Livak, K.J.; Mikkelsen, T.S.; Rinn, J.L. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells. Nat. Biotechnol. 2014, 32, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Berge, K.V.D.; de Bézieux, H.R.; Street, K.; Saelens, W.; Cannoodt, R.; Saeys, Y.; Dudoit, S.; Clement, L. Trajectory-based differential expression analysis for single-cell sequencing data. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lo Presti, E.; Dieli, F. and Meraviglia, S. Lymphopenia in COVID-19: Gammadelta T Cells-Based Therapeutic Opportunities. Vaccines (Basel) 2021, 9, 562. [Google Scholar] [CrossRef]
- Caron, J.; Ridgley, L.A.; Bodman-Smith, M. How to Train Your Dragon: Harnessing Gamma Delta T Cells Antiviral Functions and Trained Immunity in a Pandemic Era. Front. Immunol. 2021, 12, 666983. [Google Scholar] [CrossRef]
- Yazdanifar, M.; Mashkour, N.; Bertaina, A. Making a case for using γδ T cells against SARS-CoV-2. Crit. Rev. Microbiol. 2020, 46, 689–702. [Google Scholar] [CrossRef]
- Orumaa, K.; Dunne, M.R. The role of unconventional T cells in COVID-19. Ir. J. Med Sci. 2021, 1–10. [Google Scholar] [CrossRef]
- Lei, L.; Qian, H.; Yang, X.; Zhang, X.; Zhang, D.; Dai, T.; Guo, R.; Shi, L.; Cheng, Y.; Zhang, B.; et al. The phenotypic changes of γδ T cells in COVID-19 patients. J. Cell. Mol. Med. 2020, 24, 11603–11606. [Google Scholar] [CrossRef]
- Moratto, D.; Chiarini, M.; Giustini, V.; Serana, F.; Magro, P.; Roccaro, A.M.; Imberti, L.; Castelli, F.; Notarangelo, L.D.; Quiros-Roldan, E. Flow Cytometry Identifies Risk Factors and Dynamic Changes in Patients with COVID-19. J. Clin. Immunol. 2020, 40, 970–973. [Google Scholar] [CrossRef]
- Carter, M.J.; Fish, M.; Jennings, A.; Doores, K.J.; Wellman, P.; Seow, J.; Acors, S.; Graham, C.; Timms, E.; Kenny, J.; et al. Peripheral immunophenotypes in children with multisystem inflammatory syndrome associated with SARS-CoV-2 infection. Nat. Med. 2020, 26, 1701–1707. [Google Scholar] [CrossRef]
- Guo, C.; Li, B.; Ma, H.; Wang, X.; Cai, P.; Yu, Q.; Zhu, L.; Jin, L.; Jiang, C.; Fang, J.; et al. Single-cell analysis of two severe COVID-19 patients reveals a monocyte-associated and tocilizumab-responding cytokine storm. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef]
- Presti, E.L.; Corsale, A.M.; Dieli, F.; Meraviglia, S. γδ cell-based immunotherapy for cancer. Expert Opin. Biol. Ther. 2019, 19, 887–895. [Google Scholar] [CrossRef]
- Simoni, Y.; Becht, E.; Fehlings, M.G.; Loh, C.Y.; Koo, S.-L.; Teng, K.W.W.; Yeong, J.P.S.; Nahar, R.; Zhang, T.; Kared, H.; et al. Bystander CD8+ T cells are abundant and phenotypically distinct in human tumour infiltrates. Nature 2018, 557, 575–579. [Google Scholar] [CrossRef]
- Oliveira, G.; Stromhaug, K.; Klaeger, S.; Kula, T.; Frederick, D.T.; Le, P.M.; Forman, J.; Huang, T.; Li, S.; Zhang, W.; et al. Phenotype, specificity and avidity of antitumour CD8+ T cells in melanoma. Nat. Cell Biol. 2021, 596, 119–125. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerapio, J.P.; Perrier, M.; Pont, F.; Tosolini, M.; Laurent, C.; Bertani, S.; Fournie, J.-J. Single-Cell RNAseq Profiling of Human γδ T Lymphocytes in Virus-Related Cancers and COVID-19 Disease. Viruses 2021, 13, 2212. https://doi.org/10.3390/v13112212
Cerapio JP, Perrier M, Pont F, Tosolini M, Laurent C, Bertani S, Fournie J-J. Single-Cell RNAseq Profiling of Human γδ T Lymphocytes in Virus-Related Cancers and COVID-19 Disease. Viruses. 2021; 13(11):2212. https://doi.org/10.3390/v13112212
Chicago/Turabian StyleCerapio, Juan Pablo, Marion Perrier, Fréderic Pont, Marie Tosolini, Camille Laurent, Stéphane Bertani, and Jean-Jacques Fournie. 2021. "Single-Cell RNAseq Profiling of Human γδ T Lymphocytes in Virus-Related Cancers and COVID-19 Disease" Viruses 13, no. 11: 2212. https://doi.org/10.3390/v13112212