
Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer’s Disease


1
Institute of Theoretical and Experimental Biophysics of Russian Academy of Sciences, 142290 Pushchino, Russia
2
Hospital Clinico Research Foundation, INCLIVA Health Research Institute, 46010 Valencia, Spain
3
Pathology Department, Faculty of Medicine, University of Valencia, 46010 Valencia, Spain
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(6), 5739; https://doi.org/10.3390/ijms24065739
Submission received: 7 February 2023
/
Revised: 14 March 2023
/
Accepted: 15 March 2023
/
Published: 17 March 2023
(This article belongs to the Section Biochemistry)
{kind=link}
Abstract
:Coronavirus disease 2019 (COVID-19) is a rapidly spreading acute respiratory infection caused by SARS-CoV-2. The pathogenesis of the disease remains unclear. Recently, several hypotheses have emerged to explain the mechanism of interaction between SARS-CoV-2 and erythrocytes, and its negative effect on the oxygen-transport function that depends on erythrocyte metabolism, which is responsible for hemoglobin-oxygen affinity (Hb-O2 affinity). In clinical settings, the modulators of the Hb-O2 affinity are not currently measured to assess tissue oxygenation, thereby providing inadequate evaluation of erythrocyte dysfunction in the integrated oxygen-transport system. To discover more about hypoxemia/hypoxia in COVID-19 patients, this review highlights the need for further investigation of the relationship between biochemical aberrations in erythrocytes and oxygen-transport efficiency. Furthermore, patients with severe COVID-19 experience symptoms similar to Alzheimer’s, suggesting that their brains have been altered in ways that increase the likelihood of Alzheimer’s. Mindful of the partly assessed role of structural, metabolic abnormalities that underlie erythrocyte dysfunction in the pathophysiology of Alzheimer’s disease (AD), we further summarize the available data showing that COVID-19 neurocognitive impairments most probably share similar patterns with known mechanisms of brain dysfunctions in AD. Identification of parameters responsible for erythrocyte function that vary under SARS-CoV-2 may contribute to the search for additional components of progressive and irreversible failure in the integrated oxygen-transport system leading to tissue hypoperfusion. This is particularly relevant for the older generation who experience age-related disorders of erythrocyte metabolism and are prone to AD, and provide an opportunity for new personalized therapies to control this deadly infection.
1. Introduction
Coronavirus disease 2019 (COVID-19) is an acute infection of the respiratory tract caused by the newly discovered virus SARS-CoV-2 [1]. At present, the disease pathogenesis remains poorly understood and there are no specific antiviral drugs with therapeutic potential for the treatment of infected patients. According to the data available, the main difference between SARS-CoV-2 infection and infections with any other known previously human coronaviruses is that the former often remains asymptomatic [2], but can lead to multiple organ failure if it spreads in conjunction with acute respiratory distress syndrome (ARDS) [3,4], with increased mortality among older people [5]. There are many causes of multiple organ failure, but the most important is that at the earliest stage of infection the virus simultaneously attacks many host cells with angiotensin converting enzyme 2 (ACE2) receptors [6], including cells of the respiratory and gastrointestinal tract, liver, heart, kidneys, central nervous system, endothelium of the blood vessels, oral mucosa, and immune cells [7].
Moreover, many different pieces of data show that immune response to the viral infection may result in excessive cytokine release, evolving into inflammatory cytokine storm that can lead to multiple organ dysfunction and become life threatening [8,9,10,11,12,13,14]. Although there is sufficient evidence to indicate that multiple organ failure is not only associated with COVID-19 [15] but is also apparently the main hallmark of disease severity, the pathogenetic mechanisms by which damage to tissues and organs occurs are still not completely understood. In this connection, based on several hypotheses regarding various mechanisms of interaction between SARS-CoV-2 and erythrocytes (red blood cells, RBCs) [16,17], it is assumed that extremely restricted oxygen supply by disturbed RBCs may be a root in the development of hypoxia-induced multiple organ injury and the main cause of high mortality of critically ill COVID-19 patients [17,18,19,20,21].
Hemoglobin (Hb) is the dominant protein in RBCs, the functions of which are to bind, transport, and unload oxygen to tissues, in a manner dependent on erythrocyte metabolism [22,23] which is responsible for hemoglobin–oxygen affinity (Hb-O2 affinity). However, the efficacy of oxygen delivery by RBCs as one of the components in the integrated gas-transport system has rarely been assessed in combination with the other indicators usually measured in a clinical setting [24], in order to detect the limit of O2 transport reach. This leads in turn to erroneous conclusions regarding the establishment of the true reasons for the development of tissue hypoxia and detection of regional perfusion abnormalities [25,26,27,28,29], thereby failing to improve the process of therapeutic assistance.
Certainly, tissue oxygenation depends not only on RBCs and Hb function, but also on the inter-dependent interplay between the respiratory and cardiovascular systems involved in oxygen transport [30,31,32]. However, despite the importance of these major systems, the final stages of oxygen delivery to tissues where gas exchange occurs are carried out exclusively by RBCs that must be functionally active and, consequently, absolutely “healthy” [33]. Therefore, when factors determining oxygen binding, transport and delivery are identified, clinicians should also focus on the indices of metabolic abnormalities of RBCs [30,34,35]. Quantitative assessment of these parameters in patients with COVID-19 has already begun, but the data from different researchers are not only contradictory [24,36,37,38,39,40] but have caused heated debate [41,42,43], and the question remains open of whether global abnormalities in erythrocyte composition found in infected patients [20] are a contributing factor to the pathophysiology of tissue hypoxia caused by SARS-CoV-2, and further research and discussion are needed (discussed below).
In light of the foregoing, this review presents emerging evidence supporting the need for more knowledge about the relationship between biochemical aberrations and the efficacy of oxygen transport by RBCs as an additional cause of progressive and irreversible failure in the integrated oxygen-transport system. This will help minimize inaccurate and even erroneous conclusions for patients, especially the older generation with age-related disorders of RBC metabolism who are prone to AD, as well as individuals with acquired changes in Hb function and those with Hb congenital anomalies [25,26,44,45].
We also briefly review potential causes for the discrepancies between the available data and certain difficulties in analyzing Hb-O2 affinity changes in laboratory and clinical conditions. Meanwhile, there are emerging reports that patients with severe COVID-19 experience various Alzheimer’s disease (AD)-related symptoms ranging from depression and agnosia to hypometabolic/hypoxic encephalopathy and dementia [46,47,48,49,50,51,52,53], suggesting that their brains have been changed in ways that increase the risk of developing the sporadic (non-genetic) form of AD, regardless of age [54]. However, it should be noted that the sporadic form of AD is recognized as multifactorial disorder [55], and debates continue regarding what pathological factors trigger the development of the disease.
Cell viability in the brain and the organ’s myriad functions are critically dependent on an uninterrupted delivery of oxygen by the blood, and even a short pause in the oxygen supply to the brain causes its damage [56]. Chronic oxygen deficiency eventually leads to irreversible brain pathology and the further advancement of permanent cognitive impairment [57,58,59], and apparently plausible vascular/erythrocytic hypotheses [60,61] attribute these findings to hypoperfusion and disturbed RBC metabolism and function, underlying the development of energy crises in the brain, neurodegeneration, and cognitive deficit [56,58,59,62,63,64,65,66,67]. According to data on the role of structural metabolic abnormalities underlying functional deficits of RBCs in AD pathophysiology [33,61,67,68,69,70,71,72,73,74,75,76], we further summarize the available knowledge indicating the possible uniformity of COVID-19 neurocognitive impairments in the context of known mechanisms of brain dysfunction in AD.
2. Intracellular RBC Metabolism, Allosteric Effectors, and Oxygen-Transport Function
RBCs are specialized cells that carry and release oxygen to the tissues for supporting aerobic energy metabolism, coupled with the synthesis of ATP in mitochondria. However, the oxygen-transport function of RBCs is often viewed in a simplified way and RBCs themselves are perceived merely as small bags filled with hemoglobin, that easily bind oxygen in the lungs and just as easily release oxygen to tissues. Meanwhile, the presence of oxygen in red blood cells is the main condition, but not the only requirement. It is generally accepted that the capacity of RBCs for binding, transport, and delivery of oxygen to tissues depends on heterotropic allosteric modulators, formed as a result of metabolic processes in membranes and inside RBCs [22,23], which in addition to hemoglobin oxygen affinity [77], regulate NO-dependent hypoxic vasodilation [78] and tissue perfusion [79,80], and preserve ion and systemic metabolic homeostasis [81].
Interestingly, RBC indices commonly measured in clinical settings, such as number of circulating RBCs, erythrocyte sedimentation rate (ESR), hemoglobin count, hematocrit test, and others, including their correct interpretation based on accepted reference values, are widely considered to play a crucial role in assessing health status and clarifying accurate diagnosis of disease [82,83]. Meanwhile, in parallel with this diagnostic, no estimate is usually given on how changes in the results of these hematology clinical laboratory tests outside the accepted reference ranges (for example, a sharp increase in ESR) affect the oxygen-transport function of RBCs.
It should furthermore be noted that RBCs are cells that “work” even under physiological conditions, in an environment that is not always comfortable. In order to resist harmful factors that occur both in the bloodstream and inside the cells [84] and may impair RBC function and promote cell death, the RBCs should be “healthy” with intact membranes and no changes in intracellular metabolic pathways [22,77,85].
Indeed, the formation of superoxide radicals constantly takes place in RBCs [86] during oxygen binding to hemoglobin in the lungs. In kidneys, when the cells are surrounded by a hypertonic environment, they experience osmotic shock. In tight capillaries where gas exchange occurs, RBCs are exposed to extensive mechanical stress [87].
However, it is recognized that under physiological conditions these factors seem to affect the function and survival of only senescent or damaged RBCs, while most populations of young and middle-aged cells are more resistant and remain “healthy” despite the constant destructive impact [88].
What are the defense mechanisms that ensure the protection of circulating RBCs and support their viability and gas-transport function?
Primarily, glycolysis and salvage pathways provide energy in the form of ATP [89]. The pentose phosphate shunt is required for NADPH production, which together with antioxidant enzymes, i.e., glutathione reductase and glutathione peroxidase, converts oxidized glutathione to reduced glutathione, a major RBC antioxidant which is important for protection against oxidative damage and maintains hemoglobin in a reduced, active form [90]. RBCs also exhibit fairly high superoxide dismutase activity, which maintains a safe level of the superoxide radical (O2•−) that is constantly formed by Hb autoxidation during oxygen binding to hemoglobin in the lungs [84,86]. Catalase and peroxiredoxin-2-peroxidase are the other antioxidant enzymes that collectively destroy hydrogen peroxide, preventing vital biomolecules from its damaging effect and, in particular, protecting Hb from oxidative destruction [88]. In addition, important supporting processes in RBCs include the NADH-cytochrome b5 reductase pathway reduces methemoglobin (metHb) that is incapable of transporting oxygen [91], and also xenobiotic detoxification by the enzyme glutathione transferase [92]. Furthermore, the proteasome machinery plays an important role in protecting the erythrocyte membrane and is involved in the degradation of oxidized Hb during membrane damage [93,94]. In the Rapoport–Luebering shunt [77], 2,3-diphosphoglycerate (2,3-DPG) is produced, which is the main allosteric modifier of Hb-O2 affinity [95]. The shunt activity is regulated by many factors, especially by the rate of glycolysis and the activities of two enzymes of non-glycolytic pathways, diphosphoglycerate mutase (DPGM) and diphosphoglycerate phosphatase (DPGP) [28]. Thus, the metabolism of RBCs is represented by multiple, interconnected systems consisting of soluble, structural proteins, carbohydrates, lipids, anions, cations, cofactors, metabolites, and antioxidants in particular amounts. If a change of at least one component in the integrated system of metabolic pathways occurs under pathological conditions, this leads at once to the loss of RBC’s integrity and functional capacity [77], and even to cell lysis directly into the bloodstream. This is extremely dangerous, since it may lead to hypoxia [96,97] and adverse outcomes following free Hb-iron-dependent cellular oxidative damage and progression of multiple organ failure, which are included in the etiology and pathophysiology of various diseases, including Alzheimer’s disease [98,99,100,101,102,103].
Therefore, in order to assess the efficiency of oxygen transfer from the lungs to the tissue of coronavirus-infected patients with hypoxemic respiratory failure [104], it is important to calculate not only how much oxygen is available in the blood in total (arterial oxygen saturation—SaO2, and partial pressure of oxygen—PaO2), but also to understand whether hemoglobin can bind the maximum possible amount of oxygen in the lungs and release the required amount of O2 to the tissue, which is determined by the Hb-O2 affinity [21,22,24,36,43,105].
Furthermore, according to the research data nearly 5% of the world’s population are hemoglobinopathy carriers [106] and to date more than 1000 Hb variants characterized by increased or decreased oxygen affinity have been identified in people from diverse ethnic groups [29,107,108]. Although the consequences of many abnormal hemoglobin variants are well known, for instance, anemia, reduced membrane deformability, premature aging and destruction of circulating red blood cells, impaired gas exchange, etc. [108,109,110], these abnormalities have limited clinical applications making it difficult to predict the severity and nature of oxygen deficiency at the tissue level. This can lead to errors when planning therapy, especially mechanical ventilation [110,111] for critically ill patients in whom Hb variants appear in indefinite proportions during infection [109]. Thus, in the presence of hemoglobin with a low affinity for O2, leading to a decrease in oxygen–hemoglobin binding in the lungs, it can be expected that prolonged use of mechanical ventilation for critically ill patients with COVID-19 can cause not only worse pulmonary outcomes [112] and respiratory failure [113], but also vasoconstriction, impaired blood flow, and limited O2 delivery to organs such as the brain, heart, skeletal muscle, and others [114,115,116]. This may explain the high incidence of multiple organ failure [117] and greater risk of poor outcomes in these patients [118,119,120]. This issue is further complicated by the fact that even patients with normal PaO2 or SaO2 but in the presence of Hb with high oxygen affinity may still present tissue hypoxia, stemming from a reduced ability to release O2 in tissues, and supplemental oxygen may be warranted irrespective of oxygen saturation or arterial oxygen partial pressure [26,109,110,121,122].
Thus, it is clear that there is a need to establish and implement in clinical laboratories a two-sided reference interval of erythrocyte indicators associated with regulation of the oxygen binding capacity of Hb [109,123], depending on gender, genetics, ethnicity, and other biological characteristics [116,124]. The use of these indicators will enable the most appropriate treatment to be selected for each patient.
3. Peculiarities of Metabolic Abnormalities of Erythrocytes in Elderly Patients: Forgotten Prognostic Factors for Severity of COVID-19?
Although all age groups are at risk for SARS-CoV-2 infection, a major feature of the infection is increased mortality among older people [125,126,127,128,129,130]. Unfortunately, the underlying mechanisms leading to worse age-related outcomes and increased mortality risk in the older age group are not fully understood. Numerous factors have been identified as predictors of poor outcomes, such as chronic comorbidities (hypertension, cardiovascular disease, diabetes [131,132,133], inadequate immune system response, mitochondrial dysfunction [134], faster virus replication and reproduction [135], Alzheimer’s disease and related dementia [136], and so forth [137]).
Moreover, it should be kept in mind that the intracellular metabolism of RBCs undergoes stable and permanent changes in older individuals [61,72,138,139,140], which under the influence of pathological factors may constitute an additional risk factor for RBC dysfunction [33]. Indeed, the RBCs of the aged persons are characterized by a reduced rate of glycolysis, pentose phosphate pathway, and cation transport, confirmed by reduced activity of all glycolytic enzymes, reduced glucose uptake rate [61,72,141,142], the formation of pyruvate and lactate, ATP concentration [143], and sharply reduced Na+,K+-ATP-ase [144], compared with younger individuals [145]. Additionally, RBC abnormalities in older subjects were associated with disturbed calcium homeostasis, higher levels of calpains, caspases-3, increased proteolysis [139,146], hydroperoxide generation, and decreased antioxidant enzyme activity [61,142,147,148,149] and circulatory lifespan [88,139,150,151]. The enzyme activity of the Rapoport–Luebering shunt, DPGM, and DPGP, which maintain stable levels of 2,3-DPG, the main regulator of Hb-O2 affinity, decreases simultaneously with a significant decrease in 2,3-DPG concentration in RBCs in the elderly [142,152,153], resulting in violation of the allosteric properties of hemoglobin [154]. Together with the currently available information, these results indicate that oxidative stress is already present to some extent in RBCs in the elderly [88,155,156,157].
Increased blood viscosity, RBC aggregation [158], modification of the band 3 protein [147], impaired blood rheology [159], and disruption of the RBC transport function [22,58,88,160] are common signs of physiological aging. It is clear that with the appearance of additional adverse factors that can interact with RBCs in the bloodstream, the propensity of already compromised cells in older subjects for disease and life-threatening hypoxemia/hypoxia increases significantly [33,61,72,142,145,160,161,162].
Thus, assessment of the biochemical diversity of RBCs in patients of different age groups, inevitably leading to different levels of adaptive response to hypoxic conditions [163,164], is useful for predicting the severity of COVID-19 and selecting individual therapies. Moreover, there is a need to identify potential reasons for inconsistent experimental results and the existing opposing views about the role of RBCs in disrupting convective oxygen transport in SARS-CoV-2 infection [20,40,165], associated not only with the patient’s age but also with the use of nonunified experimental design that does not permit the researcher to consider a myriad of variables affecting the results.
4. Potential Causes of Discrepancies in Available Data and Challenges in Analyzing Changes in the Affinity of O2 for Hb in Laboratory and Clinical Settings
It is common knowledge that internal Hb-O2 affinity in vivo is very high [166], meaning that allosteric effectors reducing the affinity of Hb-O2 must be continuously formed in RBCs. In the absence of these effectors, high Hb-O2 affinity improves oxygen loading in the lungs but impairs oxygen unloading at the tissue level. As mentioned above, the glycolytic intermediate 2,3-DPG, quantitatively the most important organic phosphate produced within RBCs [167,168], is one of the main allosteric ligands promoting oxygen offloading in the tissues by lowering Hb-O2 affinity [22] and plays a prime role in modulating the ability of the whole blood to transport oxygen. The reduction in Hb-O2 affinity, associated with RBC metabolism through 2,3-DPG with an oxygen-transport function, is a complex and tightly regulated process [123] where the quantitative relationship between the concentration of 2,3-DPG and oxygen release is very precise [123,169,170,171,172]. Its violation leads to increased risk of adverse outcomes in many conditions associated with hypoxemia/hypoxia, as has been repeatedly proven [77,173,174,175,176,177,178,179,180,181].
In clinical settings, however, Hb-O2 affinity is routinely measured by the oxygen dissociation curve (ODC), a graphical representation of the relationship between oxygen saturation and oxygen partial pressure, where p50 (the oxygen tension when Hb is 50% saturated with oxygen) is used as the sole descriptor of Hb-O2 affinity, usually calculated under standard conditions (pH: 7.4, PCO2: 40 mmHg, temperature: 37 °C) [36,41,182,183], i.e., in the “artificial normalization” of a potentially very abnormal blood-flow environment in a seriously ill patient, leading to the elimination of many influencing factors that can change the ODC unpredictably [43], thereby leading to conflicting results [184,185]. Another bottleneck in most previous investigations of Hb-O2 affinity was that the concentration of 2,3-DPG in RBCs of patients with SARS-CoV-2 infection was not usually measured [41], and the concentrations of Hb, phosphate, age (aging red cells or patients), history of heart failure [39], or a fixed value of 2,3-DPG [186,187] were used as a sensitive index in predicting 2,3-DPG concentration. However, the steady-state concentration of 2,3-DPG in RBCs varies widely among patients with the same disease, and does not always correlate with P50, Hb concentration, or plasma phosphate [188,189,190,191]. Therefore, in routine clinical practice it would be desirable to add the measurement of RBC 2,3-DPG concentration, together with identification of the endogenous factors influencing its concentration and thus influencing ODC/Hb-O2 affinity [36].
Furthermore, some patients may have chronic diseases [192,193] and permanently receive drugs such as different types of hormones [170,194,195,196], β-adrenoblockers [197], anesthetics [198], hydroxychloroquine [39,199], salicylates [200], and others [192] which might influence the ODC. Surprisingly, articles on the measurement of ODC/Hb-O2 affinity in patients with COVID-19 have been published without this information [41]. Therefore, it can be assumed that the absence of a more complete set of factors affecting the position of the ODC can lead to either under- or over-estimation of true Hb-O2 affinity, particularly considering the relatively small numbers of patients in each study and poor matches between patients and controls in terms of gender, age, etc., and the lack of a unified experimental design for evaluating the ODC and calculating P50 [41,43]. This is especially the case in COVID-19 patients with severe anemia and hypoxia, reflecting to a limited extent (if at all) the impaired oxygen-transport function of RBCs [201].
In practice, mixed findings for Hb-O2 affinity have been obtained in COVID-19 patients. Thus, no change of Hb-O2 affinity was revealed in some investigations [37,38,39,40,202], while in others the researchers reported increases [36,186] or decreases in this index [20], suggesting the need to measure additional markers relating to tissue oxygen perfusion. For example, in some COVID-19 patients, the appearance of rigid RBCs, including mushroom-shaped RBCs [203], schizocytes [204], and sphero-echinocytes with irreversible metabolic, structural changes [205] and phosphatidylserine inversion [206] in the bloodstream points to an alteration in membrane charge [207], structural membrane damage [20], and decreased turnover rate [208]. Altogether, this can dramatically increase aggregation [209] and reduce microvascular perfusion [210,211], eventually affecting the efficiency of oxygen transport [212,213]. Therefore, the analysis of changes in the morphology and appearance of rigid cells in the bloodstream of the COVID-19 patient can enrich knowledge of the progression of impaired tissue perfusion. However, the metabolic and morphometric parameters of RBCs can vary significantly depending on the experimental conditions [207,214], and the use of a wide range of methods may often lead to inconsistency between measured indicators and those obtained in vivo, leading to difficulties in their interpretation.
According to the recommendations of the International Committee for Standardization in Haematology [215], structural and functional RBC characteristics should be studied using highly purified cells obtained by the fractionating method [216] from blood collected in a uniform manner and previously purified from leukocytes and platelets [217], using sensitive, quantitative methods that are specific for RBCs [215,218,219]. Concurrently, consideration should also be given to the plausible effects of circadian rhythm [220], patient hydration status [221], physical activity [222], recent fluid and food intake [223], recent blood transfusions [224], drugs [225], ambient temperature [226], humidity, sample collection and storage procedures [227,228], selection of an anticoagulant [229], cell washing, resuspension, and lysis buffering [230], and many other factors described in new guidelines for hemorheological laboratory techniques [231] and by experts from the World Health Organization [232]. The crucial importance of such an approach is emphasized by the fact that neither biologists nor clinicians fully take into account the influence of many factors, and therefore sometimes use RBC data that do not necessarily reflect the in vivo state characteristic of a particular pathology [221]. This leads to conflicting results and conclusions, and limits the understanding of the etiological role of RBCs as one of the components of a complex oxygen-transport system involved in the development of life-threatening hypoxemia/hypoxia and multiple organ dysfunction during infection [233]. To avoid this, and to improve the effectiveness of treatment for COVID-19 patients and turn this deadly disease into a well-controlled infection, the current practice should obviously be changed as soon as possible. Understanding the role of RBCs in COVID-19 can assist the choice of treatment and therapies against this infection, and is critical for the success of continued control of infectious diseases worldwide.
5. Erythrocyte Metabolic Disturbances as an Indicator of Mental Disorders: A Possible Link between SARS-CoV-2 Infection and Alzheimer’s Disease
To maintain cell viability and a myriad of highly energy-demanding functions, the brain the highest level of oxidative metabolism compared with the other organs, producing the majority of ATP [57,234,235,236] as a source of energy. Paradoxically, the brain has limited reserves of oxygen [237], and therefore requires a continuous supply of oxygenated blood. As a result, irregular oxygen delivery to the brain for even a short period causes brain damage [235] leading to a loss of consciousness, and chronic oxygen deficiency (regardless of the etiology of the disease) can result in hypoperfusion and irreversible brain damage, thereby provoking encephalopathy with clinical symptoms ranging from minimal changes in intellectual function to dementia and coma, often fatal outcomes [59,238,239,240,241,242,243,244], suggesting some common biochemical background for neurological signs in different diseases.
In recent years, advanced research into RBCs isolated from the blood of patients suffering from various diseases has revealed highly significant positive correlations between numerous biochemical and morphological aberrations in RBCs and the development of various types of dementia syndromes [70,74,75,76,162,245,246,247,248,249]. In particular, it became possible to formulate the vascular/erythrocytic hypothesis of the development of energy crisis in the brain, neurodegeneration, and cognitive deficit at onset of AD. These disorders are considered to be stimulated by hypoperfusion and impaired RBC metabolism and function limiting the continuous supply of oxygen to the brain [33,61,67,162,250,251,252,253]. SARS-CoV-2 infection is no exception. Considering the importance of attitudes towards the development of oxygen deficiency in the brain, the identification and quantification of RBC parameters is instrumental in gaining insight into the reasons for the development of AD-like neurological symptoms in patients with severe COVID-19, at any age [49,54,254,255,256].
Available metabolomics data indicate that RBCs from COVID-19 patients had significantly enhanced glycolysis rates compared with controls, as evidenced by both the accumulation of ATP and many other glycolytic intermediates and by high levels of rate-limiting enzymes of glycolysis. Levels of 2,3-DPG, the main allosteric modifier of Hb-O2 affinity, were also significantly increased in RBCs from COVID-19 patients [20]. The authors concluded that “increases in RBC glycolytic metabolites are consistent with a theoretically improved capacity of hemoglobin to off-load oxygen as a function of allosteric modulation by high-energy phosphate compounds, perhaps to counteract COVID-19-induced hypoxia” [20].
Indeed, there is strong evidence that increases in 2,3-DPG and ATP concentration (decreased Hb-O2 affinity) in circulating RBCs, with simultaneous significantly increased rates of glucose consumption, are among the main compensatory mechanisms that arise sometimes quite rapidly [257] to improve O2 delivery to tissues under grossly inadequate conditions [123], for example, at high altitudes [175] or in patients with hypoxemic/hypoxic diseases such as anemia and cardiopulmonary insufficiency [22,258,259,260,261]. It is apparent that a similar compensatory mechanism can also occur during SARS-CoV-2 infection, the main pathophysiological consequences of which are progressive hypoxia [262] and hypoxia-related multiorgan pathology [263,264].
However, the authors of the above article [20] also found that SARS-CoV-2 infection is associated with impaired structural membrane homeostasis at the level of cytoskeleton proteins and membrane lipids, as well as with a “higher degree of oxidative stress” in patients’ RBCs [20]. It is therefore impossible to assume that COVID-19 patients have completely overcome their difficulties with oxygen transport, or that 2,3-DPG-adaptive response of RBCs would be enough to protect tissues against damage following hypoxia.
It is generally accepted that clear signs indicating disturbances in erythrocyte membranes underlie the loss of specific biconcave shape, deformability, and progressive loss of membrane surface area in cells [219,265,266,267,268,269,270,271,272], which in combination with oxidative stress [273], vesiculation of cells [274], and significant systemic microcirculatory dysfunction [209] may dramatically affect oxygen delivery to tissues in patients with almost any diseases [275].
Given this causal relationship, it can be suggested that the reduced Hb-O2 affinity (increased 2,3-DPG) found by the authors in RBCs of COVID-19 patients [20] could allow oxygen to freely detach from Hb at the site of gas exchange. However, it remains unclear how the non-deformable cells found in COVID-19 patients [209,276], with abnormal morphology [203,277] and oxidative stress [278] leading to capillary occlusion [279] and impaired RBC flow [85,88,278], are able not only to counteract COVID-19-induced hypoxia, but even to reach the sites of gas exchange [280]. Therefore, the question arises as to how nondeformable RBCs overwhelmed by oxidant stress are able to traverse capillary beds that have diameters less than those of RBCs [279], engage in gas exchange, and then restore the normal their shapes for further travel.
In addition, the combined effect of the destruction of membrane structures and the presence of a higher degree of oxidative stress in the RBCs of patients with COVID-19 [20], leads to the formation of metHb [281,282,283,284,285,286], unable to carry oxygen [88,90,287,288] and thereby the initiator of encephalopathy, the clinical manifestations of which (depending on the degree of metHb accumulation) [289] can vary from asymptomatic to severe hypoxia that does not respond to supplemental oxygen therapy, leading to changes in mental status, coma, and death [290,291,292].
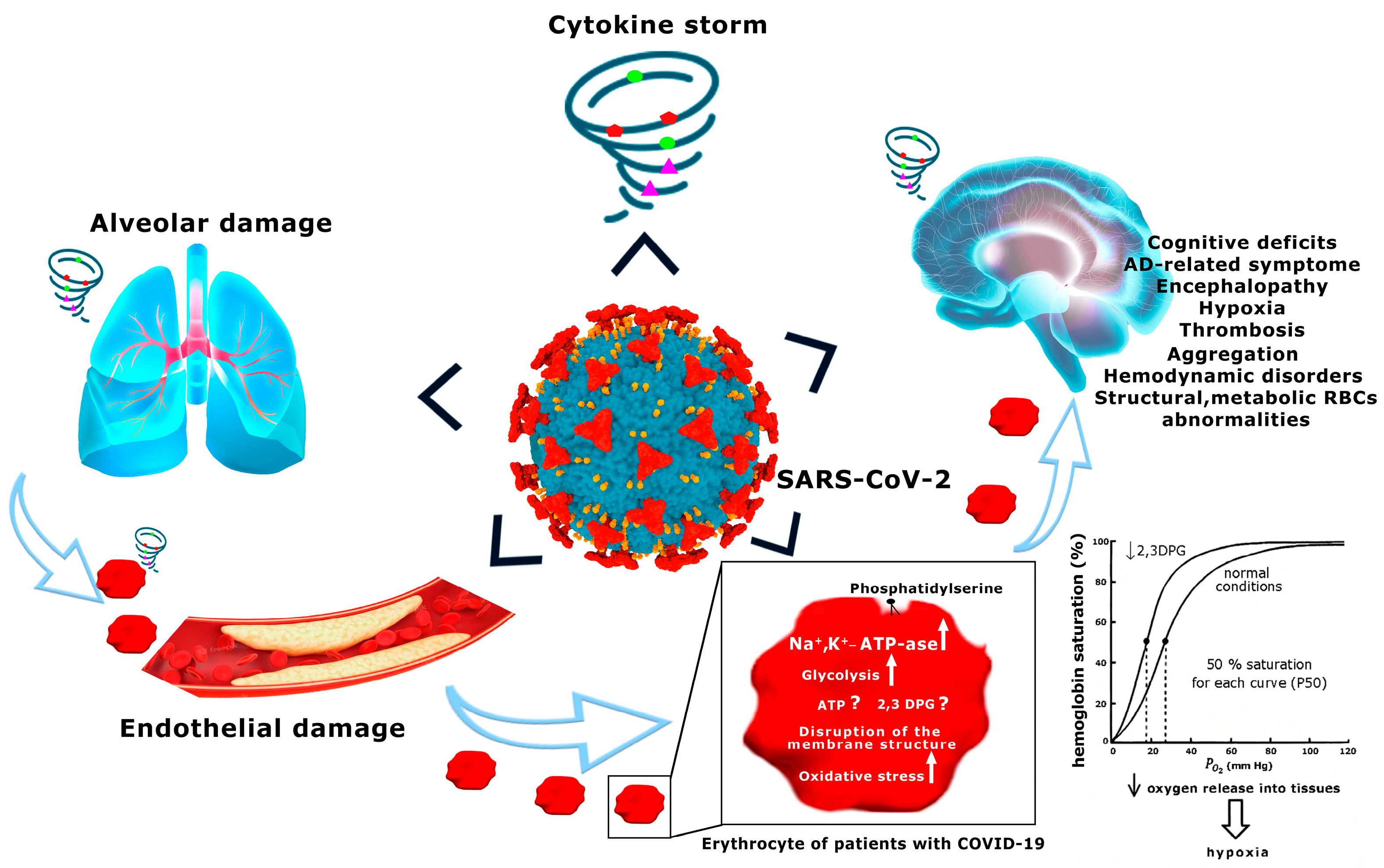
It is quite obvious that the RBCs are highly susceptible to SARS-CoV-2-induced injury [293,294,295,296], and can provide early warning signals of abnormalities of tissue oxygenation [19,297], which in AD can lead to neurological symptoms [33,72,161,162,298,299] and complications of the disease (Figure 1). However, lack of sufficient available data on RBCs of COVID-19 patients, and, moreover, misleading interpretations of these data, do not allow for full-scale comparison of the changes occurring in RBCs in COVID-19 and AD. In this case, it is only possible to point out some similarities.
Firstly, global membrane defects [300,301], proteolysis [302], strong impairment of blood flow, deformability, and shape [248], and oxidative stress [145,303,304] are the main pathological changes of RBCs in AD patients [145,148,305], which are identical to those found in patients with COVID-19 [203,209,276,293].
Secondly, in addition to these structural and biochemical abnormalities, our recent work also indicates that the values of all parameters of the glycolytic system are significantly higher in RBCs of AD patients than those obtained from age-matched controls, indicating an acceleration of the RBC glycolytic pathway [142,148], as in COVID-19 [20].
The exception was that the concentrations of ATP and 2,3-DPG would be significantly reduced in patients with AD [148,153] compared with the control, while in patients with COVID-19, the levels of these metabolites in RBSs compared with the control would be increased, as stated above [20]. The exact causes of this inconsistency are not clear and may be multifactorial.
One of the factors that very probably contributes to altered energy metabolism in the RBCs of AD patients is the enhanced entry of Na+ into the cells [306,307]. Activation of glycolysis in the RBCs of AD patients can apparently be considered an adaptive process aimed at maintaining the ATP concentration necessary for up-regulation of Na+,K+-ATP-ase, which in turn maintains ionic homeostasis in cells. Indeed, the activity of Na+,K+-ATP-ase in the RBCs of AD patients was increased compared with that found in age-matched controls [61,153,308]. As already noted, this increase in enzyme activity was accompanied by a decrease in ATP concentration and its enhanced breakdown, which was confirmed by the accumulation of ADP and AMP and indicates an imbalance between the rate of ATP formation and the pumping of ions, enhanced by an increase in Na+ influx into the RBCs of AD patients. In general, this means that constant and multiple activation of the enzyme in the RBCs of older AD patients experiencing restrictions in adapting to changes cannot completely compensate for the increased plasma membrane permeability that leads to negative consequences in chronic conditions [33,309], including enhancing the hydrolysis of ATP, 2,3-DPG, oxidative stress, and proteolysis [310]. The effect of this pathology, in combination with changes in the morphological and rheological properties, decreased the ability to deform in elderly RBCs [140,311], promoting dysfunction of RBCs mediated by a prolonged phase of deficient oxygen supply [140,312], thereby lowering the threshold for development of neuropathology [33,61,72,162,313]. Considering that the age of AD [314] and COVID-19 patients [20] was different and was somewhat heterogeneous in the COVID-19 group (mean age 58.4 ± 20.9 and 75 ± 2.6 for COVID-19 and AD patients, respectively), it is likely that the appearance of multidirectional changes in ATP and 2,3-DPG with a simultaneous increase in glycolysis in the RBCs of COVID-19 and AD patients reflects different age-related adaptive responses both to hypoxic conditions [315,316] and to different types of cytotoxic agents, although other factors cannot be excluded [317,318,319,320]. Considering the similarity of disturbances in the structure, morphology, energy metabolism, and antioxidant system of RBCs in these two diseases, it is therefore conceivable that cognitive impairment associated with hypoxia may have similar mechanisms. Further studies are needed to investigate this relationship and to confirm the clinical significance of the biomarkers of “RBC illness” as among the main components of a progressive and irreversible insufficiency of the integrated oxygen-transport system. These data may well hold the keys to understanding the pathogenesis and progression of irreversible brain damage and the permanent impairment of cognition that may significantly aggravate the course of severe systemic disease such as COVID-19 [321,322].
6. The Underappreciated Role of Erythrocytes in COVID-19
SARS-CoV-2 infection has spread rapidly around the world, but despite the collaborative efforts of the scientific community and clinicians there is to date no consensus regarding the pathogenesis of the disease, and there remains no effective antiviral therapy for COVID-19. A meta-analysis based on the findings of fundamental and clinical research provided evidence of strong correlation between numerous inflammatory markers, especially C-reactive protein (CRP), interleukin-6 (IL-6), ESR, and severity of COVID-19 [323,324,325,326]. This confirmed a suggestion that hyperinflammation (cytokine storm) is a basic pathologic mechanism of life-threatening multiple organ dysfunction caused by direct cytokine-induced tissue damage [8,9,10,11,12,13,14].
Surprisingly, RBCs are not included in the list of “affected” areas although they are also damaged during hyperinflammation [288,327,328,329], and lose their function [327,328] as oxygen carriers [330] and as “organs” impacting systemic metabolic homeostasis [81], and this predictable pathological relationship is not a tool for diagnosing “disease” relating to the RBCs themselves. It is noteworthy that IL-6 acts on a large number of cells and tissues [331] and, especially importantly, interacts with the RBCs [332] causing redistribution of membrane proteins and phospholipids [75], morphological changes, impaired deformability, increased oxidative stress [333], and accelerated cell aging [334], which altogether might be an important driving force in the development and progression of oxygen-transport dysfunction [322,324,331,335,336,337]. In light of this, we believe that underestimated metabolic and structural abnormalities of RBCs in COVID-19 patients should be an important area for future systematic research.
In addition, an increased ESR value obtained as a result of neutralization of negative charges on the surface of RBCs by acute phase proteins circulating in the blood during the inflammatory process [338] is exclusively perceived as an inflammation marker (although non-specific) [339]. The abnormalities of the RBCs themselves are not considered in clinical practice, although these may be associated with the harmful consequences of a decrease in charge on their surface [207,340], affecting not only their functional abilities but also the rate of capillary blood flow [341], the state of endothelial surface of blood vessels (negatively charged) [338,342,343], and in general the rheological properties of the blood.
It is obvious that the study of the state of RBCs and early diagnosis of their changes during the inflammatory process is very important for analyzing the state of blood flow in the microcirculation, which in turn largely depends on the ability of RBCs to deform [344]. It is essential to understand these data, especially considering that measurement of arterial oxygen saturation (SpO2) by the most common method of pulse oximetry under conditions of low blood flow and elevated carboxyhemoglobin and metHb levels [345] can lead to misinterpretation of gas exchange between the blood and tissues, delaying correct diagnosis and timely initiation of therapeutic interventions [346,347]. Therefore, we suggest that if the erythrocytes of COVID-19 patients contain increased amounts of abnormal forms of hemoglobin [281,348], these patients should be monitored using alternative indicators to avoid false results. Keeping in mind that the relationship between RBCs’ biochemical aberrations and their transport efficacy is a critical missing link in the understanding of life-threatening hypoxemia/hypoxia and multiple organ dysfunction during infection [233], we believe that the biomarkers of metabolic, antioxidant status disorders must be evaluated, including morphological and structural changes of the RBCs that form part of the integrated oxygen-transport system [18,19,20,21].
Finally, functional neuro-imaging studies of patients in different COVID-19 phases have shown that oxygen desaturation significantly influences the degree of brain hypometabolism and related cognitive deterioration [349,350]. Just as importantly, similar patterns of brain metabolic dysfunction have been identified in pediatric patients [351], supporting the accepted contemporary view that cerebral hypoperfusion and disturbed RBC metabolism and function are independent of the specific cause of the disease, and the patient’s age is the main trigger leading to the development of an energy crisis in the brain, neurodegeneration, and cognitive deficit [244,352,353], similar to those found in Alzheimer’s disease.
7. Conclusions
SARS-CoV-2 infection has spread rapidly around the world, but despite collaborative efforts of the scientific community and clinicians there is no consensus to date regarding the pathogenesis of the disease and remains no effective antiviral therapy for COVID-19. In addition to attacks on the respiratory system, Alzheimer’s-related symptoms have been widely reported in patients with COVID-19 and in those who have recovered from it. The mechanisms of brain pathology leading to cognitive dysfunction in COVID-19 patients are still not fully understood, and are multiple, but hypoxia-related brain pathology appears to be the main trigger involved in this disease. RBCs comprise one of the components in the integrated gas-transport system and are the only cells that carry oxygen to tissues, thus ensuring the vital activity of the whole organism. While researchers have long recognized the potential links between metabolic RBC aberration and hypoxia-related multiorgan failure, including the brain, the efficacy of oxygen delivery by RBCs is rarely assessed in combination with the other indicators usually measured in clinical settings to detect limited O2 transport.
Given the importance of addressing the development of functional disturbance of oxygen transport in RBCs, we suggest that a thorough and detailed study of erythrocyte metabolism and morphology is needed, not only to identify potential risk factors but also to shed light on the molecular mechanisms that limit oxygen supply to the tissues. This knowledge will be important to prevent the development of hypoxia-induced multiorgan failure and neurodegenerative processes, typical not only of AD but also of many other diseases with different etiologies, and characterized by varying degrees of cognitive dysfunctions, especially in COVID-19.
The development of technologies to assist in revascularization and restoration of erythrocyte metabolism must form an integral part of new therapeutic strategies in the treatment of a great variety of disorders associated with inadequate oxygen delivery, including devastating illnesses of human beings such as COVID-19.
Author Contributions
E.K. and C.M. provided the original conception and designed the study; E.K., L.T., G.A. and C.M. analyzed the data and discussed the results. The manuscript was written by E.K. and approved by all authors. All authors have read and agreed to the published version of the manuscript.
Funding
This study has been conducted in accordance with the Government Contract N° 075-01025-23-00 ITEB RAS assigned for the year 2023 and 2024–2025 planning period and supported by Agencia Valenciana de Innovación, Generalitat Valenciana (Consolidacio Cadena Valor); Fundación Ramón Areces; Consellería Educación, Generalitat Valenciana (CIPROM2021/082) co-funded with European Regional Development Funds (ERDF); donation by F. Sarabia (PRV00225) to C.M.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within this article.
Conflicts of Interest
The authors declare no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Oran, D.P.; Topol, E.J. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Ann. Intern. Med. 2020, 173, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Du, Z.; Zhu, F.; Cao, Z.; An, Y.; Gao, Y.; Jiang, B. Comorbidities and Multi-Organ Injuries in the Treatment of COVID-19. Lancet 2020, 395, e52. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, T.; Hassani, F.; Ghaffari, N.; Ebrahimi, B.; Yarahmadi, A.; Hassanzadeh, G. COVID-19 and Multiorgan Failure: A Narrative Review on Potential Mechanisms. J. Mol. Histol. 2020, 51, 613–628. [Google Scholar] [CrossRef]
- Zhao, H.-L.; Huang, Y.-M.; Huang, Y. Mortality in Older Patients with COVID-19. J. Am. Geriatr. Soc. 2020, 68, 1685–1687. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.C.; Lely, A.T.; Navis, G.J.; van Goor, H. Tissue Distribution of ACE2 Protein, the Functional Receptor for SARS Coronavirus. A First Step in Understanding SARS Pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Loganathan, S.; Kuppusamy, M.; Wankhar, W.; Gurugubelli, K.R.; Mahadevappa, V.H.; Lepcha, L.; Choudhary, A.K. Angiotensin-Converting Enzyme 2 (ACE2): COVID 19 Gate Way to Multiple Organ Failure Syndromes. Respir. Physiol. Neurobiol. 2021, 283, 103548. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, X.; Zheng, X.; Luo, S.; Xu, S.; Weng, J. Targeting Inflammation and Cytokine Storm in COVID-19. Pharmacol. Res. 2020, 159, 105051. [Google Scholar] [CrossRef]
- Henderson, L.A.; Canna, S.W.; Schulert, G.S.; Volpi, S.; Lee, P.Y.; Kernan, K.F.; Caricchio, R.; Mahmud, S.; Hazen, M.M.; Halyabar, O.; et al. On the Alert for Cytokine Storm: Immunopathology in COVID-19. Arthritis Rheumatol. 2020, 72, 1059–1063. [Google Scholar] [CrossRef] [Green Version]
- Castanares-Zapatero, D.; Chalon, P.; Kohn, L.; Dauvrin, M.; Detollenaere, J.; Maertens de Noordhout, C.; Primus-de Jong, C.; Cleemput, I.; Van den Heede, K. Pathophysiology and Mechanism of Long COVID: A Comprehensive Review. Ann. Med. 2022, 54, 1473–1487. [Google Scholar] [CrossRef]
- Galeotti, C.; Bayry, J. Autoimmune and Inflammatory Diseases Following COVID-19. Nat. Rev. Rheumatol. 2020, 16, 413–414. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, P.; Fallahi, M.S.; Erabi, G.; Pakdin, M.; Zarezadeh, S.M.; Faridzadeh, A.; Entezari, S.; Ansari, A.; Poudineh, M.; Deravi, N. Multisystem Inflammatory Syndrome and Autoimmune Diseases Following COVID-19: Molecular Mechanisms and Therapeutic Opportunities. Front. Mol. Biosci. 2022, 9, 804109. [Google Scholar] [CrossRef] [PubMed]
- Borczuk, A.C.; Yantiss, R.K. The Pathogenesis of Coronavirus-19 Disease. J. Biomed. Sci. 2022, 29, 87. [Google Scholar] [CrossRef] [PubMed]
- Zaim, S.; Chong, J.H.; Sankaranarayanan, V.; Harky, A. COVID-19 and Multiorgan Response. Curr. Probl. Cardiol. 2020, 45, 100618. [Google Scholar] [CrossRef] [PubMed]
- Cosic, I.; Cosic, D.; Loncarevic, I. RRM Prediction of Erythrocyte Band3 Protein as Alternative Receptor for SARS-CoV-2 Virus. Appl. Sci. 2020, 10, 4053. [Google Scholar] [CrossRef]
- Liu, W.; Li, H. COVID-19: Attacks the 1-Beta Chain of Hemoglobin to Disrupt Respiratory Function and Escape Immunity. ChemRxiv 2022. [Google Scholar] [CrossRef]
- Cavezzi, A.; Troiani, E.; Corrao, S. COVID-19: Hemoglobin, Iron, and Hypoxia beyond Inflammation. A Narrative Review. Clin. Pract. 2020, 10, 1271. [Google Scholar] [CrossRef]
- Hacking, S.M. Red Blood Cell Exchange for SARS-CoV-2: A Gemini of Therapeutic Opportunities. Med. Hypotheses 2020, 144, 110227. [Google Scholar] [CrossRef]
- Thomas, T.; Stefanoni, D.; Dzieciatkowska, M.; Issaian, A.; Nemkov, T.; Hill, R.C.; Francis, R.O.; Hudson, K.E.; Buehler, P.W.; Zimring, J.C.; et al. Evidence of Structural Protein Damage and Membrane Lipid Remodeling in Red Blood Cells from COVID-19 Patients. J. Proteome Res. 2020, 19, 4455–4469. [Google Scholar] [CrossRef]
- Woyke, S.; Rauch, S.; Ströhle, M.; Gatterer, H. Modulation of Hb-O2 Affinity to Improve Hypoxemia in COVID-19 Patients. Clin. Nutr. 2021, 40, 38–39. [Google Scholar] [CrossRef]
- Brewer, G.J.; Eaton, J.W. Erythrocyte Metabolism: Interaction with Oxygen Transport. Science 1971, 171, 1205–1211. [Google Scholar] [CrossRef]
- Brewer, G.J.; Oelshlegel, F.J.; Moore, L.G.; Noble, N.A. In Vivo Red Cell Glycolytic Control and DPG-ATP Levels. Ann. N. Y. Acad. Sci. 1974, 241, 513–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelsalam, M. Permissive Hypoxemia: Is It Time to Change Our Approach? Chest 2006, 129, 210–211. [Google Scholar] [CrossRef] [Green Version]
- Verhovsek, M.; Henderson, M.P.A.; Cox, G.; Luo, H.; Steinberg, M.H.; Chui, D.H.K. Unexpectedly Low Pulse Oximetry Measurements Associated with Variant Hemoglobins: A Systematic Review. Am. J. Hematol. 2010, 85, 882–885. [Google Scholar] [CrossRef]
- Zur, B.; Bagci, S.; Ludwig, M.; Stoffel-Wagner, B. Oxygen Saturation in Pulse Oximetry in Hemoglobin Anomalies. Klin. Padiatr. 2012, 224, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Henig, N.R.; Pierson, D.J. Mechanisms of Hypoxemia. Respir. Care Clin. N. Am. 2000, 6, 501–521. [Google Scholar] [CrossRef]
- Balcerek, B.; Steinach, M.; Lichti, J.; Maggioni, M.A.; Becker, P.N.; Labes, R.; Gunga, H.-C.; Persson, P.B.; Fähling, M. A Broad Diversity in Oxygen Affinity to Haemoglobin. Sci. Rep. 2020, 10, 16920. [Google Scholar] [CrossRef] [PubMed]
- Deyell, R.; Jackson, S.; Spier, S.; Le, D.; Poon, M.-C. Low Oxygen Saturation by Pulse Oximetry May Be Associated with a Low Oxygen Affinity Hemoglobin Variant, Hemoglobin Titusville. J. Pediatr. Hematol. Oncol. 2006, 28, 100–102. [Google Scholar] [CrossRef]
- Pittman, R.N. Regulation of Tissue Oxygenation; Integrated Systems Physiology: From Molecule to Function to Disease; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2011. [Google Scholar]
- MacIntyre, N.R. Tissue Hypoxia: Implications for the Respiratory Clinician. Respir. Care 2014, 59, 1590–1596. [Google Scholar] [CrossRef]
- Zubieta-Calleja, G.; Zubieta-DeUrioste, N.; Venkatesh, T.; Das, K.K.; Soliz, J. COVID-19 and Pneumolysis Simulating Extreme High-Altitude Exposure with Altered Oxygen Transport Physiology; Multiple Diseases, and Scarce Need of Ventilators: Andean Condor’s-Eye-View. Rev. Recent. Clin. Trials 2020, 15, 347–359. [Google Scholar] [CrossRef]
- Bosman, G.J.C.G.M. Disturbed Red Blood Cell Structure and Function: An Exploration of the Role of Red Blood Cells in Neurodegeneration. Front. Med. 2018, 5, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misiti, F. SARS-CoV-2 Infection and Red Blood Cells: Implications for Long Term Symptoms during Exercise. Sports Med. Health Sci. 2021, 3, 181–182. [Google Scholar] [CrossRef] [PubMed]
- Wagner, P.D. Oxygen Transport from Air to Tissues as an Integrated System: What Limits Maximal O2 Consumption in Health and Disease? In Structure-Function Relationships in Various Respiratory Systems: Connecting to the Next Generation; Yamaguchi, K., Ed.; Respiratory Disease Series: Diagnostic Tools and Disease Managements; Springer: Singapore, 2020; pp. 191–217. ISBN 9789811555961. [Google Scholar]
- Vogel, D.J.; Formenti, F.; Retter, A.J.; Vasques, F.; Camporota, L. A Left Shift in the Oxyhaemoglobin Dissociation Curve in Patients with Severe Coronavirus Disease 2019 (COVID-19). Br. J. Haematol. 2020, 191, 390–393. [Google Scholar] [CrossRef]
- Daniel, Y.; Hunt, B.J.; Retter, A.; Henderson, K.; Wilson, S.; Sharpe, C.C.; Shattock, M.J. Haemoglobin Oxygen Affinity in Patients with Severe COVID-19 Infection. Br. J. Haematol. 2020, 190, e126–e127. [Google Scholar] [CrossRef] [PubMed]
- Laredo, M.; Curis, E.; Masson-Fron, E.; Voicu, S.; Mégarbane, B. Does COVID-19 Alter the Oxyhemoglobin Dissociation Curve?—An Observational Cohort Study Using a Mixed-Effect Modelling. Clin. Chem. Lab. Med. 2021, 59, e416–e419. [Google Scholar] [CrossRef] [PubMed]
- Gille, T.; Sesé, L.; Aubourg, E.; Fabre, E.E.; Cymbalista, F.; Ratnam, K.C.; Valeyre, D.; Nunes, H.; Richalet, J.-P.; Planès, C. The Affinity of Hemoglobin for Oxygen Is Not Altered During COVID-19. Front. Physiol. 2021, 12, 578708. [Google Scholar] [CrossRef]
- Park, K.C.; Donovan, K.; McKechnie, S.; Ramamurthy, N.; Klenerman, P.; Swietach, P. Single-Cell Oxygen Saturation Imaging Shows That Gas Exchange by Red Blood Cells Is Not Impaired in COVID-19 Patients. Br. J. Haematol. 2020, 190, e229–e232. [Google Scholar] [CrossRef] [PubMed]
- Böning, D.; Kuebler, W.M.; Bloch, W. The Oxygen Dissociation Curve of Blood in COVID-19. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321, L349–L357. [Google Scholar] [CrossRef]
- Shattock, M.J.; Daniel, Y.; Hunt, B.J.; Retter, A.; Henderson, K.; Wilson, S.; Sharpe, C.C. COVID-19 and Haemoglobin Oxygen Affinity: Some Clarity? Br. J. Haematol. 2020, 190, 723–724. [Google Scholar] [CrossRef]
- Harutyunyan, G.; Harutyunyan, G.; Mkhoyan, G.; Harutyunyan, V.; Soghomonyan, S. Haemoglobin Oxygen Affinity in Patients with Severe COVID-19 Infection: Still Unclear. Br. J. Haematol. 2020, 190, 725–726. [Google Scholar] [CrossRef]
- Hladik, A.; Lynshue, K. Dyshemoglobinemias and Pulse Oximetry: A Therapeutic Challenge. J. Pediatr. Hematol. Oncol. 2008, 30, 850–852. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.M.; Mortier, N.A.; Hyde, K.S.; Hankins, J.S.; Ware, R.E. The Diagnostic Dilemma of Congenital Unstable Hemoglobinopathies. Pediatr. Blood Cancer 2010, 55, 1393–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangon, C.-M.; Krantic, S.; Moyse, E.; Fougère, B. The Vagal Autonomic Pathway of COVID-19 at the Crossroad of Alzheimer’s Disease and Aging: A Review of Knowledge. J. Alzheimers Dis. Rep. 2020, 4, 537–551. [Google Scholar] [CrossRef] [PubMed]
- Roy, D.; Ghosh, R.; Dubey, S.; Dubey, M.J.; Benito-León, J.; Kanti Ray, B. Neurological and Neuropsychiatric Impacts of COVID-19 Pandemic. Can. J. Neurol. Sci. 2021, 48, 9–24. [Google Scholar] [CrossRef] [PubMed]
- Collantes, M.E.V.; Espiritu, A.I.; Sy, M.C.C.; Anlacan, V.M.M.; Jamora, R.D.G. Neurological Manifestations in COVID-19 Infection: A Systematic Review and Meta-Analysis. Can. J. Neurol. Sci. 2021, 48, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Maury, A.; Lyoubi, A.; Peiffer-Smadja, N.; de Broucker, T.; Meppiel, E. Neurological Manifestations Associated with SARS-CoV-2 and Other Coronaviruses: A Narrative Review for Clinicians. Rev. Neurol. 2021, 177, 51–64. [Google Scholar] [CrossRef]
- Garg, R.K. Spectrum of Neurological Manifestations in Covid-19: A Review. Neurol. India 2020, 68, 560–572. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-Acute COVID-19 Syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Sullivan, B.N.; Fischer, T. Age-Associated Neurological Complications of COVID-19: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2021, 13, 653694. [Google Scholar] [CrossRef]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, A.A.; Shetty, A.K.; Sprigg, N.; Auer, D.P.; Constantinescu, C.S. Delirium as a Presenting Feature in COVID-19: Neuroinvasive Infection or Autoimmune Encephalopathy? Brain Behav. Immun. 2020, 88, 68–70. [Google Scholar] [CrossRef]
- Royall, D.R. Alzheimer Disease as a Vascular Disorder: Nosological Evidence. Stroke 2002, 33, 2147–2148. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, J.C. Are Major Dementias Triggered by Poor Blood Flow to the Brain? Theoretical Considerations. J. Alzheimers Dis. 2017, 57, 353–371. [Google Scholar] [CrossRef] [PubMed]
- Blass, J.P. Glucose/Mitochondria in Neurological Conditions. Int. Rev. Neurobiol. 2002, 51, 325–376. [Google Scholar] [CrossRef] [PubMed]
- Blass, J.P.; Gibson, G.E. Cerebrometabolic Aspects of Delirium in Relationship to Dementia. Dement. Geriatr. Cogn. Disord. 1999, 10, 335–338. [Google Scholar] [CrossRef]
- de la Torre, J.C. Critically Attained Threshold of Cerebral Hypoperfusion: The CATCH Hypothesis of Alzheimer’s Pathogenesis. Neurobiol. Aging 2000, 21, 331–342. [Google Scholar] [CrossRef]
- de la Torre, J. The Vascular Hypothesis of Alzheimer’s Disease: A Key to Preclinical Prediction of Dementia Using Neuroimaging. J. Alzheimers Dis. 2018, 63, 35–52. [Google Scholar] [CrossRef]
- Kosenko, E.; Tikhonova, L.; Alilova, G.; Urios, A.; Montoliu, C. The Erythrocytic Hypothesis of Brain Energy Crisis in Sporadic Alzheimer Disease: Possible Consequences and Supporting Evidence. J. Clin. Med. 2020, 9, 206. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, J.C. Pathophysiology of Neuronal Energy Crisis in Alzheimer’s Disease. Neurodegener. Dis. 2008, 5, 126–132. [Google Scholar] [CrossRef]
- Perry, G.; Nunomura, A.; Raina, A.K.; Aliev, G.; Siedlak, S.L.; Harris, P.L.R.; Casadesus, G.; Petersen, R.B.; Bligh-Glover, W.; Balraj, E.; et al. A Metabolic Basis for Alzheimer Disease. Neurochem. Res. 2003, 28, 1549–1552. [Google Scholar] [CrossRef] [PubMed]
- De Felice, F.G.; Lourenco, M.V. Brain Metabolic Stress and Neuroinflammation at the Basis of Cognitive Impairment in Alzheimer’s Disease. Front. Aging Neurosci. 2015, 7, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blass, J.P. Cerebrometabolic Abnormalities in Alzheimer’s Disease. Neurol. Res. 2003, 25, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Mergenthaler, P.; Lindauer, U.; Dienel, G.A.; Meisel, A. Sugar for the Brain: The Role of Glucose in Physiological and Pathological Brain Function. Trends. Neurosci. 2013, 36, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevenson, A.; Lopez, D.; Khoo, P.; Kalaria, R.N.; Mukaetova-Ladinska, E.B. Exploring Erythrocytes as Blood Biomarkers for Alzheimer’s Disease. J. Alzheimers Dis. 2017, 60, 845–857. [Google Scholar] [CrossRef]
- Carelli-Alinovi, C.; Dinarelli, S.; Sampaolese, B.; Misiti, F.; Girasole, M. Morphological Changes Induced in Erythrocyte by Amyloid Beta Peptide and Glucose Depletion: A Combined Atomic Force Microscopy and Biochemical Study. Biochim. Biophys. Acta Biomembr. 2019, 1861, 236–244. [Google Scholar] [CrossRef]
- Clementi, M.E.; Giardina, B.; Colucci, D.; Galtieri, A.; Misiti, F. Amyloid-Beta Peptide Affects the Oxygen Dependence of Erythrocyte Metabolism: A Role for Caspase 3. Int. J. Biochem. Cell Biol. 2007, 39, 727–735. [Google Scholar] [CrossRef]
- Bosman, G.J.; Bartholomeus, I.G.; de Man, A.J.; van Kalmthout, P.J.; de Grip, W.J. Erythrocyte Membrane Characteristics Indicate Abnormal Cellular Aging in Patients with Alzheimer’s Disease. Neurobiol. Aging 1991, 12, 13–18. [Google Scholar] [CrossRef]
- Bourdel-Marchasson, I.; Delmas-Beauvieux, M.C.; Peuchant, E.; Richard-Harston, S.; Decamps, A.; Reignier, B.; Emeriau, J.P.; Rainfray, M. Antioxidant Defences and Oxidative Stress Markers in Erythrocytes and Plasma from Normally Nourished Elderly Alzheimer Patients. Age Ageing 2001, 30, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Kosenko, E.A.; Tikhonova, L.A.; Montoliu, C.; Barreto, G.E.; Aliev, G.; Kaminsky, Y.G. Metabolic Abnormalities of Erythrocytes as a Risk Factor for Alzheimer’s Disease. Front. Neurosci. 2018, 11, 728. [Google Scholar] [CrossRef] [Green Version]
- Graham, C.; Santiago-Mugica, E.; Abdel-All, Z.; Li, M.; McNally, R.; Kalaria, R.N.; Mukaetova-Ladinska, E.B. Erythrocytes as Biomarkers for Dementia: Analysis of Protein Content and Alpha-Synuclein. J. Alzheimers Dis. 2019, 71, 569–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pretorius, E.; Olumuyiwa-Akeredolu, O.-O.O.; Mbotwe, S.; Bester, J. Erythrocytes and Their Role as Health Indicator: Using Structure in a Patient-Orientated Precision Medicine Approach. Blood Rev. 2016, 30, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, E.; Swanepoel, A.C.; Buys, A.V.; Vermeulen, N.; Duim, W.; Kell, D.B. Eryptosis as a Marker of Parkinson’s Disease. Aging 2014, 6, 788–819. [Google Scholar] [CrossRef] [Green Version]
- Bester, J.; Soma, P.; Kell, D.B.; Pretorius, E. Viscoelastic and Ultrastructural Characteristics of Whole Blood and Plasma in Alzheimer-Type Dementia, and the Possible Role of Bacterial Lipopolysaccharides (LPS). Oncotarget 2015, 6, 35284–35303. [Google Scholar] [CrossRef] [Green Version]
- van Wijk, R.; van Solinge, W.W. The Energy-Less Red Blood Cell Is Lost: Erythrocyte Enzyme Abnormalities of Glycolysis. Blood 2005, 106, 4034–4042. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M.; Lancaster, J.R. Hemoglobin-Mediated, Hypoxia-Induced Vasodilation via Nitric Oxide: Mechanism(s) and Physiologic versus Pathophysiologic Relevance. Am. J. Respir. Cell. Mol. Biol. 2005, 32, 257–261. [Google Scholar] [CrossRef]
- Crawford, J.H.; Isbell, T.S.; Huang, Z.; Shiva, S.; Chacko, B.K.; Schechter, A.N.; Darley-Usmar, V.M.; Kerby, J.D.; Lang, J.D.; Kraus, D.; et al. Hypoxia, Red Blood Cells, and Nitrite Regulate NO-Dependent Hypoxic Vasodilation. Blood 2006, 107, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Mairbäurl, H. Red Blood Cells in Sports: Effects of Exercise and Training on Oxygen Supply by Red Blood Cells. Front. Physiol. 2013, 4, 332. [Google Scholar] [CrossRef] [Green Version]
- Nemkov, T.; Reisz, J.A.; Xia, Y.; Zimring, J.C.; D’Alessandro, A. Red Blood Cells as an Organ? How Deep Omics Characterization of the Most Abundant Cell in the Human Body Highlights Other Systemic Metabolic Functions beyond Oxygen Transport. Expert Rev. Proteom. 2018, 15, 855–864. [Google Scholar] [CrossRef]
- Katayev, A.; Balciza, C.; Seccombe, D.W. Establishing Reference Intervals for Clinical Laboratory Test Results: Is There a Better Way? Am. J. Clin. Pathol. 2010, 133, 180–186. [Google Scholar] [CrossRef] [Green Version]
- Ozarda, Y. Reference Intervals: Current Status, Recent Developments and Future Considerations. Biochem. Med. 2016, 26, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, V.; Diederich, L.; Keller, T.C.S.; Kramer, C.M.; Lückstädt, W.; Panknin, C.; Suvorava, T.; Isakson, B.E.; Kelm, M.; Cortese-Krott, M.M. Red Blood Cell Function and Dysfunction: Redox Regulation, Nitric Oxide Metabolism, Anemia. Antioxid. Redox Signal. 2017, 26, 718–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Rosa, M.C.; Carelli Alinovi, C.; Galtieri, A.; Scatena, R.; Giardina, B. The Plasma Membrane of Erythrocytes Plays a Fundamental Role in the Transport of Oxygen, Carbon Dioxide and Nitric Oxide and in the Maintenance of the Reduced State of the Heme Iron. Gene 2007, 398, 162–171. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide Dismutase. An Enzymic Function for Erythrocuprein (Hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Evans, E.A. Structure and Deformation Properties of Red Blood Cells: Concepts and Quantitative Methods. Methods Enzymol. 1989, 173, 3–35. [Google Scholar] [CrossRef]
- Mohanty, J.G.; Nagababu, E.; Rifkind, J.M. Red Blood Cell Oxidative Stress Impairs Oxygen Delivery and Induces Red Blood Cell Aging. Front. Physiol. 2014, 5, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, S.; Kenanov, D. Adenine and Adenosine Salvage Pathways in Erythrocytes and the Role of S-Adenosylhomocysteine Hydrolase. A Theoretical Study Using Elementary Flux Modes. FEBS J. 2005, 272, 5278–5290. [Google Scholar] [CrossRef]
- Stromme, J.H.; Eldjarn, L. The Role of the Pentose Phosphate Pathway in the Reduction of Methaemoglobin in Human Erythrocytes. Biochem. J. 1962, 84, 406–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bando, S.; Takano, T.; Yubisui, T.; Shirabe, K.; Takeshita, M.; Nakagawa, A. Structure of Human Erythrocyte NADH-Cytochrome B5 Reductase. Acta. Crystallogr. D Biol. Crystallogr. 2004, 60, 1929–1934. [Google Scholar] [CrossRef]
- Ansari, G.A.; Singh, S.V.; Gan, J.C.; Awasthi, Y.C. Human Erythrocyte Glutathione S-Transferase: A Possible Marker of Chemical Exposure. Toxicol. Lett. 1987, 37, 57–62. [Google Scholar] [CrossRef]
- Sae-Lee, W.; McCafferty, C.L.; Verbeke, E.J.; Havugimana, P.C.; Papoulas, O.; McWhite, C.D.; Houser, J.R.; Vanuytsel, K.; Murphy, G.J.; Drew, K.; et al. The Protein Organization of a Red Blood Cell. Cell. Rep. 2022, 40, 111103. [Google Scholar] [CrossRef] [PubMed]
- Anastasiadi, A.T.; Tzounakas, V.L.; Dzieciatkowska, M.; Arvaniti, V.-Z.; Papageorgiou, E.G.; Papassideri, I.S.; Stamoulis, K.; D’Alessandro, A.; Kriebardis, A.G.; Antonelou, M.H. Innate Variability in Physiological and Omics Aspects of the Beta Thalassemia Trait-Specific Donor Variation Effects. Front. Physiol. 2022, 13, 907444. [Google Scholar] [CrossRef] [PubMed]
- Benesch, R.; Benesch, R.E. The Effect of Organic Phosphates from the Human Erythrocyte on the Allosteric Properties of Hemoglobin. Biochem. Biophys. Res. Commun. 1967, 26, 162–167. [Google Scholar] [CrossRef]
- MacDonald, R. Red Cell 2,3-Diphosphoglycerate and Oxygen Affinity. Anaesthesia 1977, 32, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, J.; Koh, N.; Sakakibara, F.; Hamada, Y.; Wakao, T.; Hara, T.; Mori, K.; Nakashima, E.; Naruse, K.; Hotta, N. Polyol Pathway, 2,3-Diphosphoglycerate in Erythrocytes and Diabetic Neuropathy in Rats. Eur. J. Pharmacol. 1995, 294, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Leaf, D.E.; Rajapurkar, M.; Lele, S.S.; Mukhopadhyay, B.; Rawn, J.D.; Frendl, G.; Waikar, S.S. Increased Plasma Catalytic Iron in Patients May Mediate Acute Kidney Injury and Death Following Cardiac Surgery. Kidney Int. 2015, 87, 1046–1054. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.-P.; Xi, G.; Keep, R.F.; Hua, Y.; Nemoianu, A.; Hoff, J.T. Brain Edema after Experimental Intracerebral Hemorrhage: Role of Hemoglobin Degradation Products. J. Neurosurg. 2002, 96, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.F.; Loftspring, M.; Wurster, W.L.; Beiler, S.; Beiler, C.; Wagner, K.R.; Pyne-Geithman, G.J. Bilirubin Oxidation Products, Oxidative Stress, and Intracerebral Hemorrhage. Acta Neurochir. Suppl. 2008, 105, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Shaver, C.M.; Upchurch, C.P.; Janz, D.R.; Grove, B.S.; Putz, N.D.; Wickersham, N.E.; Dikalov, S.I.; Ware, L.B.; Bastarache, J.A. Cell-Free Hemoglobin: A Novel Mediator of Acute Lung Injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L532–L541. [Google Scholar] [CrossRef] [Green Version]
- Drvenica, I.T.; Stančić, A.Z.; Maslovarić, I.S.; Trivanović, D.I.; Ilić, V.L. Extracellular Hemoglobin: Modulation of Cellular Functions and Pathophysiological Effects. Biomolecules 2022, 12, 1708. [Google Scholar] [CrossRef]
- Rapozzi, V.; Juarranz, A.; Habib, A.; Ihan, A.; Strgar, R. Is Haem the Real Target of COVID-19? Photodiagn. Photodyn. Ther. 2021, 35, 102381. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, J.L. Respiratory Failure. Blood Purif. 2002, 20, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Morgan, T.J. The Oxyhaemoglobin Dissociation Curve in Critical Illness. Crit. Care Resusc. 1999, 1, 93–100. [Google Scholar] [PubMed]
- Angastiniotis, M.; Modell, B. Global Epidemiology of Hemoglobin Disorders. Ann. N. Y. Acad. Sci. 1998, 850, 251–269. [Google Scholar] [CrossRef] [PubMed]
- Chui, D.H.; Hardison, R.; Riemer, C.; Miller, W.; Carver, M.F.; Molchanova, T.P.; Efremov, G.D.; Huisman, T.H. An Electronic Database of Human Hemoglobin Variants on the World Wide Web. Blood 1998, 91, 2643–2644. [Google Scholar]
- Yasar, S.J.; Ravn Berg, V.I.; Ahmad, A.; Doll, D. Hemoglobin Titusville: A Rare Low Oxygen Affinity Hemoglobinopathy. Clin. Case Rep. 2017, 5, 1011–1012. [Google Scholar] [CrossRef] [Green Version]
- Schechter, A.N. Hemoglobin Research and the Origins of Molecular Medicine. Blood 2008, 112, 3927–3938. [Google Scholar] [CrossRef] [Green Version]
- Mangin, O. High Oxygen Affinity Hemoglobins. Rev. Med. Int. 2017, 38, 106–112. [Google Scholar] [CrossRef]
- Wunsch, H. Mechanical Ventilation in COVID-19: Interpreting the Current Epidemiology. Am. J. Respir. Crit. Care Med. 2020, 202, 1–4. [Google Scholar] [CrossRef]
- Sharma, S.; Kelly, Y.P.; Palevsky, P.M.; Waikar, S.S. Intensity of Renal Replacement Therapy and Duration of Mechanical Ventilation: Secondary Analysis of the Acute Renal Failure Trial Network Study. Chest 2020, 158, 1473–1481. [Google Scholar] [CrossRef]
- Alsumrain, M.H.; Jawad, S.A.; Imran, N.B.; Riar, S.; DeBari, V.A.; Adelman, M. Association of Hypophosphatemia with Failure-to-Wean from Mechanical Ventilation. Ann. Clin. Lab. Sci. 2010, 40, 144–148. [Google Scholar]
- Sjöberg, F.; Singer, M. The Medical Use of Oxygen: A Time for Critical Reappraisal. J. Intern. Med. 2013, 274, 505–528. [Google Scholar] [CrossRef]
- Broccard, A. Prone Position, High-Frequency Oscillatory Ventilation, and Hippocrates in Acute Respiratory Distress Syndrome. Crit. Care Med. 2005, 33, 2407–2408. [Google Scholar] [CrossRef]
- Tu, Y.; Yang, P.; Zhou, Y.; Wen, X.; Li, Q.; Zhou, J.; Wang, J.; Hu, J.; He, N.; Wang, K.; et al. Risk Factors for Mortality of Critically Ill Patients with COVID-19 Receiving Invasive Ventilation. Int. J. Med. Sci. 2021, 18, 1198–1206. [Google Scholar] [CrossRef]
- Slutsky, A.S.; Tremblay, L.N. Multiple System Organ Failure. Is Mechanical Ventilation a Contributing Factor? Am. J. Respir. Crit. Care Med. 1998, 157, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Bhatraju, P.K.; Ghassemieh, B.J.; Nichols, M.; Kim, R.; Jerome, K.R.; Nalla, A.K.; Greninger, A.L.; Pipavath, S.; Wurfel, M.M.; Evans, L.; et al. Covid-19 in Critically Ill Patients in the Seattle Region—Case Series. N. Engl. J. Med. 2020, 382, 2012–2022. [Google Scholar] [CrossRef]
- Telle, K.E.; Grøsland, M.; Helgeland, J.; Håberg, S.E. Factors Associated with Hospitalization, Invasive Mechanical Ventilation Treatment and Death among All Confirmed COVID-19 Cases in Norway: Prospective Cohort Study. Scand. J. Public Health 2021, 49, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Nevins, M.L.; Epstein, S.K. Predictors of Outcome for Patients with COPD Requiring Invasive Mechanical Ventilation. Chest 2001, 119, 1840–1849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsia, C.C. Respiratory Function of Hemoglobin. N. Engl. J. Med. 1998, 338, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Ryerson, G.; Block, E. Safe Use of Oxygen Therapy: A Physiologic Approach. Part 2. Respir. Ther. 1983, 13, 25–30. [Google Scholar]
- Mairbäurl, H.; Weber, R.E. Oxygen Transport by Hemoglobin. Compr. Physiol. 2012, 2, 1463–1489. [Google Scholar] [CrossRef]
- Horn, P.S.; Pesce, A.J. Reference Intervals: An Update. Clin. Chim. Acta 2003, 334, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical Course and Risk Factors for Mortality of Adult Inpatients with COVID-19 in Wuhan, China: A Retrospective Cohort Study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Kang, S.J.; Jung, S.I. Age-Related Morbidity and Mortality among Patients with COVID-19. Infect. Chemother. 2020, 52, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.G.; Jarvis, C.I.; CMMID COVID-19 Working Group; Edmunds, W.J.; Jewell, N.P.; Diaz-Ordaz, K.; Keogh, R.H. Increased Mortality in Community-Tested Cases of SARS-CoV-2 Lineage, B.1.1.7. Nature 2021, 593, 270–274. [Google Scholar] [CrossRef]
- Yanez, N.D.; Weiss, N.S.; Romand, J.-A.; Treggiari, M.M. COVID-19 Mortality Risk for Older Men and Women. BMC Public Health 2020, 20, 1742. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Ning, R.; Tao, Y.; Yu, C.; Deng, X.; Zhao, C.; Meng, S.; Tang, F.; Xu, D. Risk Factors for Mortality in 244 Older Adults With COVID-19 in Wuhan, China: A Retrospective Study. J. Am. Geriatr. Soc. 2020, 68, E19–E23. [Google Scholar] [CrossRef]
- Poloni, T.E.; Carlos, A.F.; Cairati, M.; Cutaia, C.; Medici, V.; Marelli, E.; Ferrari, D.; Galli, A.; Bognetti, P.; Davin, A.; et al. Prevalence and Prognostic Value of Delirium as the Initial Presentation of COVID-19 in the Elderly with Dementia: An Italian Retrospective Study. EClinicalMedicine 2020, 26, 100490. [Google Scholar] [CrossRef] [PubMed]
- de Erausquin, G.A.; Snyder, H.; Carrillo, M.; Hosseini, A.A.; Brugha, T.S.; Seshadri, S. CNS SARS-CoV-2 Consortium The Chronic Neuropsychiatric Sequelae of COVID-19: The Need for a Prospective Study of Viral Impact on Brain Functioning. Alzheimers Dement. 2021, 17, 1056–1065. [Google Scholar] [CrossRef]
- Goh, K.J.; Choong, M.C.; Cheong, E.H.; Kalimuddin, S.; Duu Wen, S.; Phua, G.C.; Chan, K.S.; Haja Mohideen, S. Rapid Progression to Acute Respiratory Distress Syndrome: Review of Current Understanding of Critical Illness from Coronavirus Disease 2019 (COVID-19) Infection. Ann. Acad. Med. Singap. 2020, 49, 108–118. [Google Scholar] [CrossRef]
- Wang, L.; He, W.; Yu, X.; Hu, D.; Bao, M.; Liu, H.; Zhou, J.; Jiang, H. Coronavirus Disease 2019 in Elderly Patients: Characteristics and Prognostic Factors Based on 4-Week Follow-Up. J. Infect. 2020, 80, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Taghizadeh-Hesary, F.; Akbari, H. The Powerful Immune System against Powerful COVID-19: A Hypothesis. Med. Hypotheses 2020, 140, 109762. [Google Scholar] [CrossRef]
- Jin, R.; Niu, C.; Wu, F.; Zhou, S.; Han, T.; Zhang, Z.; Li, E.; Zhang, X.; Xu, S.; Wang, J.; et al. DNA Damage Contributes to Age-Associated Differences in SARS-CoV-2 Infection. Aging Cell 2022, 21, e13729. [Google Scholar] [CrossRef] [PubMed]
- Portacolone, E.; Byers, A.L.; Halpern, J.; Barnes, D.E. Addressing Suicide Risk in Patients Living With Dementia During the COVID-19 Pandemic and Beyond. Gerontologist 2022, 62, 956–963. [Google Scholar] [CrossRef]
- Cooper, I.D.; Crofts, C.A.P.; DiNicolantonio, J.J.; Malhotra, A.; Elliott, B.; Kyriakidou, Y.; Brookler, K.H. Relationships between Hyperinsulinaemia, Magnesium, Vitamin D, Thrombosis and COVID-19: Rationale for Clinical Management. Open Heart 2020, 7, e001356. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Picot, I.; Trivier, J.M.; Nicole, A.; Sinet, P.M.; Thevenin, M. Age-Correlated Modifications of Copper-Zinc Superoxide Dismutase and Glutathione-Related Enzyme Activities in Human Erythrocytes. Clin. Chem. 1992, 38, 66–70. [Google Scholar] [CrossRef]
- Kosower, N.S. Altered Properties of Erythrocytes in the Aged. Am. J. Hematol. 1993, 42, 241–247. [Google Scholar] [CrossRef]
- Ajmani, R.S.; Rifkind, J.M. Hemorheological Changes during Human Aging. Gerontology 1998, 44, 111–120. [Google Scholar] [CrossRef]
- Güven, M.; Ozkiliç, A.; Kanigür-Sultuybek, G.; Ulutin, T. Age-Related Changes on Glucose Transport and Utilization of Human Erythrocytes: Effect of Oxidative Stress. Gerontology 1999, 45, 79–82. [Google Scholar] [CrossRef]
- Kosenko, E.A.; Aliev, G.; Kaminsky, Y.G. Relationship between Chronic Disturbance of 2,3-Diphosphoglycerate Metabolism in Erythrocytes and Alzheimer Disease. CNS Neurol. Disord. Drug Targets 2016, 15, 113–123. [Google Scholar] [CrossRef]
- Rodgers, G.P.; Lichtman, H.C.; Sheff, M.F. Red Blood Cell Glucose-6-Phosphate Dehydrogenase Activity in Aged Humans. J. Am. Geriatr. Soc. 1983, 31, 8–11. [Google Scholar] [CrossRef]
- Rabini, R.A.; Petruzzi, E.; Staffolani, R.; Tesei, M.; Fumelli, P.; Pazzagli, M.; Mazzanti, L. Diabetes Mellitus and Subjects’ Ageing: A Study on the ATP Content and ATP-Related Enzyme Activities in Human Erythrocytes. Eur. J. Clin. Investig. 1997, 27, 327–332. [Google Scholar] [CrossRef]
- Kosenko, E.A.; Solomadin, I.N.; Tikhonova, L.A.; Reddy, V.P.; Aliev, G.; Kaminsky, Y.G. Pathogenesis of Alzheimer Disease: Role of Oxidative Stress, Amyloid-β Peptides, Systemic Ammonia and Erythrocyte Energy Metabolism. CNS Neurol. Disord. Drug Targets 2014, 13, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Glaser, T.; Schwarz-Benmeir, N.; Barnoy, S.; Barak, S.; Eshhar, Z.; Kosower, N.S. Calpain (Ca2+-Dependent Thiol Protease) in Erythrocytes of Young and Old Individuals. Proc. Natl. Acad. Sci. USA 1994, 91, 7879–7883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz-Ben Meir, N.; Glaser, T.; Kosower, N.S. Band 3 Protein Degradation by Calpain Is Enhanced in Erythrocytes of Old People. Biochem. J. 1991, 275 Pt 1, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosenko, E.A.; Aliev, G.; Tikhonova, L.A.; Li, Y.; Poghosyan, A.C.; Kaminsky, Y.G. Antioxidant Status and Energy State of Erythrocytes in Alzheimer Dementia: Probing for Markers. CNS Neurol. Disord. Drug Targets 2012, 11, 926–932. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Markers of Oxidative Stress in Erythrocytes and Plasma during Aging in Humans. Oxid. Med. Cell. Longev. 2010, 3, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, M.J.; Meiselman, H.J.; Baskurt, O.K. Blood Rheology and Aging. J. Geriatr. Cardiol. 2013, 10, 291–301. [Google Scholar] [CrossRef]
- Raberin, A.; Burtscher, J.; Connes, P.; Millet, G.P. Hypoxia and Hemorheological Properties in Older Individuals. Ageing Res. Rev. 2022, 79, 101650. [Google Scholar] [CrossRef]
- Purcell, Y.; Brozović, B. Red Cell 2,3-Diphosphoglycerate Concentration in Man Decreases with Age. Nature 1974, 251, 511–512. [Google Scholar] [CrossRef]
- Kaminsky, Y.G.; Reddy, V.P.; Ashraf, G.M.; Ahmad, A.; Benberin, V.V.; Kosenko, E.A.; Aliev, G. Age-Related Defects in Erythrocyte 2,3-Diphosphoglycerate Metabolism in Dementia. Aging Dis. 2013, 4, 244–255. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.C.; Carelli Alinovi, C.; Galtieri, A.; Russo, A.; Giardina, B. Allosteric Properties of Hemoglobin and the Plasma Membrane of the Erythrocyte: New Insights in Gas Transport and Metabolic Modulation. IUBMB Life 2008, 60, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Resveratrol Up-Regulates the Erythrocyte Plasma Membrane Redox System and Mitigates Oxidation-Induced Alterations in Erythrocytes during Aging in Humans. Rejuvenation Res. 2013, 16, 232–240. [Google Scholar] [CrossRef]
- Maurya, P.K.; Kumar, P.; Chandra, P. Biomarkers of Oxidative Stress in Erythrocytes as a Function of Human Age. World J. Methodol. 2015, 5, 216–222. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.I.; Maurya, P.K. Markers of Oxidative Stress in Erythrocytes during Aging in Humans. Ann. N. Y. Acad. Sci. 2007, 1100, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Christy, R.M.; Baskurt, O.K.; Gass, G.C.; Gray, A.B.; Marshall-Gradisnik, S.M. Erythrocyte Aggregation and Neutrophil Function in an Aging Population. Gerontology 2010, 56, 175–180. [Google Scholar] [CrossRef]
- Baskurt, O.K.; Meiselman, H.J. Blood Rheology and Hemodynamics. Semin. Thromb. Hemost. 2003, 29, 435–450. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.; Doctor, A. Red Blood Cell Dysfunction in Critical Illness. Crit. Care Clin. 2020, 36, 267–292. [Google Scholar] [CrossRef]
- Meltzer, H.L. Is There a Specific Membrane Defect in Bipolar Disorders? Biol. Psychiatry 1991, 30, 1071–1074. [Google Scholar] [CrossRef]
- Ponizovsky, A.M.; Barshtein, G.; Bergelson, L.D. Biochemical Alterations of Erythrocytes as an Indicator of Mental Disorders: An Overview. Harv. Rev. Psychiatry 2003, 11, 317–332. [Google Scholar] [CrossRef]
- Chaleckis, R.; Murakami, I.; Takada, J.; Kondoh, H.; Yanagida, M. Individual Variability in Human Blood Metabolites Identifies Age-Related Differences. Proc. Natl. Acad. Sci. USA 2016, 113, 4252–4259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racine, M.L.; Dinenno, F.A. Reduced Deformability Contributes to Impaired Deoxygenation-Induced ATP Release from Red Blood Cells of Older Adult Humans. J. Physiol. 2019, 597, 4503–4519. [Google Scholar] [CrossRef]
- Reva, I.; Yamamoto, T.; Rasskazova, M.; Lemeshko, T.; Usov, V.; Krasnikov, Y.; Fisenko, A.; Kotsyurbiy, E.; Tudakov, V.; Tsegolnik, E.; et al. Erythrocytes as a Target of SARS COV-2 in Pathogenesis of COVID-19. Archiv. Euromedica 2020, 10, 5–11. [Google Scholar] [CrossRef]
- Weber, R.E.; Fago, A. Functional Adaptation and Its Molecular Basis in Vertebrate Hemoglobins, Neuroglobins and Cytoglobins. Respir. Physiol. Neurobiol. 2004, 144, 141–159. [Google Scholar] [CrossRef] [PubMed]
- Benesch, R.; Benesch, R.E. Intracellular Organic Phosphates as Regulators of Oxygen Release by Haemoglobin. Nature 1969, 221, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Klocke, R.A. Oxygen Transport and 2,3-Diphosphoglycerate (DPG). Chest 1972, 62, 79S–85S. [Google Scholar] [CrossRef] [Green Version]
- Ditzel, J.; Standl, E. The Problem of Tissue Oxygenation in Diabetes Mellitus: Evidence of disordered oxygen release from the erythrocytes of diabetics in various conditions of metabolic control. Acta Med. Scand. Suppl. 1975, 578, 59–68. [Google Scholar] [CrossRef]
- Ditzel, J.; Jaeger, P.; Standl, E. An Adverse Effect of Insulin on the Oxygen-Release Capacity of Red Blood Cells in Nonacidotic Diabetics. Metabolism 1978, 27, 927–934. [Google Scholar] [CrossRef]
- Duhm, J. Effects of 2,3-Diphosphoglycerate and Other Organic Phosphate Compounds on Oxygen Affinity and Intracellular PH of Human Erythrocytes. Pflug. Arch. 1971, 326, 341–356. [Google Scholar] [CrossRef]
- Böning, D.; Bloch, W.; Kuebler, W.M. Comment on Ceruti et al. Temporal Changes in the Oxyhemoglobin Dissociation Curve of Critically Ill COVID-19 Patients. J. Clin. Med. 2022, 11, 788. J. Clin. Med. 2022, 11, 3376. [Google Scholar] [CrossRef]
- Sharma, S.; Brugnara, C.; Betensky, R.A.; Waikar, S.S. Reductions in Red Blood Cell 2,3-Diphosphoglycerate Concentration during Continuous Renal Replacment Therapy. Clin. J. Am. Soc. Nephrol. 2015, 10, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohmer, P.R.; Dawson, R.B. The Significance of 2,3-DPG in Red Blood Cell Transfusions. CRC Crit. Rev. Clin. Lab. Sci. 1979, 11, 107–174. [Google Scholar] [CrossRef] [PubMed]
- Lenfant, C.; Torrance, J.; English, E.; Finch, C.A.; Reynafarje, C.; Ramos, J.; Faura, J. Effect of Altitude on Oxygen Binding by Hemoglobin and on Organic Phosphate Levels. J. Clin. Investig. 1968, 47, 2652–2656. [Google Scholar] [CrossRef] [Green Version]
- Lenfant, C.; Torrance, J.D.; Reynafarje, C. Shift of the O2-Hb Dissociation Curve at Altitude: Mechanism and Effect. J. Appl. Physiol. 1971, 30, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Oski, F.A.; Gottlieb, A.J.; Delivoria-Papadopoulos, M.; Miller, W.W. Red-Cell 2,3-Diphosphoglycerate Levels in Subjects with Chronic Hypoxemia. N. Engl. J. Med. 1969, 280, 1165–1166. [Google Scholar] [CrossRef]
- Bunn, H.F.; Jandl, J.H. Control of Hemoglobin Function within the Red Cell. N. Engl. J. Med. 1970, 282, 1414–1421. [Google Scholar] [CrossRef]
- Valentine, W.N.; Tanaka, K.R.; Paglia, D.E. Hemolytic Anemias and Erythrocyte Enzymopathies. Ann. Intern. Med. 1985, 103, 245–257. [Google Scholar] [CrossRef]
- Hodgkin, J.E. The Importance of 2,3-Diphosphoglycerate in Regulation of Tissue Oxygenation. West. J. Med. 1975, 123, 44. [Google Scholar]
- Ditzel, J.; Standl, E. The Problem of Tissue Oxygenation in Diabetes Mellitus. I. Its Relation to the Early Functional Changes in the Microcirculation of Diabetic Subjects. Acta Med. Scand. Suppl. 1975, 578, 49–58. [Google Scholar]
- Collins, J.-A.; Rudenski, A.; Gibson, J.; Howard, L.; O’Driscoll, R. Relating Oxygen Partial Pressure, Saturation and Content: The Haemoglobin-Oxygen Dissociation Curve. Breathe 2015, 11, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Severinghaus, J.W. Simple, Accurate Equations for Human Blood O2 Dissociation Computations. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1979, 46, 599–602. [Google Scholar] [CrossRef] [Green Version]
- Gille, T.; Sesé, L.; Aubourg, E.; Bernaudin, J.-F.; Richalet, J.-P.; Planès, C. Is There a Shift of the Oxygen-Hemoglobin Dissociation Curve in COVID-19? Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 322, L174–L175. [Google Scholar] [CrossRef] [PubMed]
- Eytan, D.; Mazwi, M.L.; Goodwin, A.J.; Goodfellow, S.D.; Assadi, A.; Greer, R.W.; Jegatheeswaran, A.; Laussen, P.C. Revisiting Oxygen Dissociation Curves and Bedside Measured Arterial Saturation in Critically Ill Children. Intensive Care Med. 2019, 45, 1832–1834. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, S.; Minotti, B.; Glotta, A.; Biggiogero, M.; Bona, G.; Marzano, M.; Greco, P.; Spagnoletti, M.; Garzoni, C.; Bendjelid, K. Temporal Changes in the Oxyhemoglobin Dissociation Curve of Critically Ill COVID-19 Patients. J. Clin. Med. 2022, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Ceruti, S.; Minotti, B.; Glotta, A.; Biggiogero, M.; Bona, G.; Marzano, M.; Greco, P.; Spagnoletti, M.; Garzoni, C.; Bendjelid, K. Reply to Böning et al. Comment on “Ceruti et al. Temporal Changes in the Oxyhemoglobin Dissociation Curve of Critically Ill COVID-19 Patients. J. Clin. Med. 2022, 11, 788”. J. Clin. Med. 2022, 11, 4547. [Google Scholar] [CrossRef]
- Ibrahim, E.e.d.S.; McLellan, S.A.; Walsh, T.S. Red Blood Cell 2,3-Diphosphoglycerate Concentration and In Vivo P50 during Early Critical Illness. Crit. Care Med. 2005, 33, 2247–2252. [Google Scholar] [CrossRef] [PubMed]
- Slawsky, P.; Desforges, J.F. Erythrocyte 2,3-Diphosphoglycerate in Iron Deficiency. Arch. Intern. Med. 1972, 129, 914–917. [Google Scholar] [CrossRef]
- Thomas, H.M.; Lefrak, S.S.; Irwin, R.S.; Fritts, H.W.; Caldwell, P.R. The Oxyhemoglobin Dissociation Curve in Health and Disease. Role of 2,3-Diphosphoglycerate. Am. J. Med. 1974, 57, 331–348. [Google Scholar] [CrossRef]
- Morgan, T.J.; Koch, D.; Morris, D.; Clague, A.; Purdie, D.M. Reduced Red Cell 2,3-Diphosphoglycerate Concentrations in Critical Illness without Decreased in Vivo P50. Anaesth. Intensive Care 2001, 29, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Naeraa, N. The Variation of Blood Oxygen Dissociation Curves in Patients. Scand. J. Clin. Lab. Investig. 1964, 16, 630–634. [Google Scholar] [CrossRef]
- Kang, S.K.; Park, S.K. 2,3-Diphosphoglyceric Acid Changes in Uremia and during Hemodialysis. Korean J. Intern. Med. 1986, 1, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Silken, A.B. Pharmacologic Manipulation of Human Erythrocyte 2,3-Diphosphoglycerate Levels by Prednisone Administration. Pediatr. Res. 1975, 9, 61–64. [Google Scholar] [CrossRef]
- Parker, J.P.; Beirne, G.J.; Desai, J.N.; Raich, P.C.; Shahidi, N.T. Androgen-Induced Increase in Red-Cell 2,3-Diphosphoglycerate. N. Engl. J. Med. 1972, 287, 381–383. [Google Scholar] [CrossRef]
- Molinari, P.F.; Chung, S.K.; Snyder, L.M. Variations of Erythrocyte Glycolysis Following Androgens. J. Lab. Clin. Med. 1973, 81, 443–446. [Google Scholar]
- Schrumpf, J.D.; Sheps, D.S.; Wolfson, S.; Aronson, A.L.; Cohen, L.S. Altered Hemoglobin-Oxygen Affinity with Long-Term Propranolol Therapy in Patients with Coronary Artery Disease. Am. J. Cardiol. 1977, 40, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.C.; Colton, E.T.; Behar, M.G. Does Anesthesia Alter Hemoglobin Dissociation? Anesthesiology 1970, 32, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.H.; Kulig, K.W.; Rumack, B.H. Drug- and Chemical-Induced Methaemoglobinaemia. Clinical Features and Management. Med. Toxicol. 1986, 1, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kravath, R.E.; Abel, G.; Colli, A.; McNamara, H.; Cohen, M.I. Salicylate Poisoning—Effect on 2,3-Diphosphoglycerate Levels in the Rat. Biochem. Pharmacol. 1972, 21, 2656–2658. [Google Scholar] [CrossRef]
- Severinghaus, J.W. Oxyhemoglobin Dissociation Curve Correction for Temperature and PH Variation in Human Blood. J. Appl. Physiol. 1958, 12, 485–486. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Guàrdia, S.; Ferrer, A.; Díaz, Ó.; Caguana, A.O.; Tejedor, E.; Bellido-Calduch, S.; Rodríguez-Chiaradia, D.A.; Gea, J. Absence of Relevant Clinical Effects of SARS-COV-2 on the Affinity of Hemoglobin for O2 in Patients with COVID-19. Arch. Bronconeumol. 2021, 57, 757–763. [Google Scholar] [CrossRef]
- Gérard, D.; Ben Brahim, S.; Lesesve, J.F.; Perrin, J. Are Mushroom-Shaped Erythrocytes an Indicator of COVID-19? Br. J. Haematol. 2021, 192, 230. [Google Scholar] [CrossRef]
- Sweeney, J.M.; Barouqa, M.; Krause, G.J.; Gonzalez-Lugo, J.D.; Rahman, S.; Gil, M.R. Evidence for Secondary Thrombotic Microangiopathy in COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Recktenwald, S.M.; Simionato, G.; Lopes, M.G.M.; Gamboni, F.; Dzieciatkowska, M.; Meybohm, P.; Zacharowski, K.; von Knethen, A.; Wagner, C.; Kaestner, L.; et al. Cross-Talk between Red Blood Cells and Plasma Influences Blood Flow and Omics Phenotypes in Severe COVID-19. eLife 2022, 11, e81316. [Google Scholar] [CrossRef] [PubMed]
- Bouchla, A.; Kriebardis, A.G.; Georgatzakou, H.T.; Fortis, S.P.; Thomopoulos, T.P.; Lekkakou, L.; Markakis, K.; Gkotzias, D.; Panagiotou, A.; Papageorgiou, E.G.; et al. Red Blood Cell Abnormalities as the Mirror of SARS-CoV-2 Disease Severity: A Pilot Study. Front. Physiol. 2021, 12, 825055. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-X.; Wu, Z.-J.; Mehrishi, J.; Huang, B.-T.; Chen, X.-Y.; Zheng, X.-J.; Liu, W.-J.; Luo, M. Human Red Blood Cell Aging: Correlative Changes in Surface Charge and Cell Properties. J. Cell. Mol. Med. 2011, 15, 2634–2642. [Google Scholar] [CrossRef] [Green Version]
- Kronstein-Wiedemann, R.; Stadtmüller, M.; Traikov, S.; Georgi, M.; Teichert, M.; Yosef, H.; Wallenborn, J.; Karl, A.; Schütze, K.; Wagner, M.; et al. SARS-CoV-2 Infects Red Blood Cell Progenitors and Dysregulates Hemoglobin and Iron Metabolism. Stem. Cell. Rev. Rep. 2022, 18, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Renoux, C.; Fort, R.; Nader, E.; Boisson, C.; Joly, P.; Stauffer, E.; Robert, M.; Girard, S.; Cibiel, A.; Gauthier, A.; et al. Impact of COVID-19 on Red Blood Cell Rheology. Br. J. Haematol. 2021, 192, e108–e111. [Google Scholar] [CrossRef]
- Cicco, G.; Cicco, S. Hemorheological Aspects in the Microvasculature of Several Pathologies. Adv. Exp. Med. Biol. 2007, 599, 7–15. [Google Scholar] [CrossRef]
- Cabrales, P. Effects of Erythrocyte Flexibility on Microvascular Perfusion and Oxygenation during Acute Anemia. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1206–H1215. [Google Scholar] [CrossRef]
- Santini, M.T.; Straface, E.; Cipri, A.; Peverini, M.; Santulli, M.; Malorni, W. Structural Alterations in Erythrocytes from Patients with Chronic Obstructive Pulmonary Disease. Haemostasis 1997, 27, 201–210. [Google Scholar] [CrossRef]
- Revin, V.; Balykova, L.; Pinyaev, S.; Syusin, I.; Radaeva, O.; Revina, N.; Kostina, Y.; Kozlov, E.; Inchina, V.; Nikitin, I.; et al. Research into Morphofunctional Characteristics of Erythrocytes in COVID-19 Patients. Biomedicines 2022, 10, 553. [Google Scholar] [CrossRef] [PubMed]
- Wong, P. A Hypothesis of the Disc-Sphere Transformation of the Erythrocytes between Glass Surfaces and of Related Observations. J. Theor. Biol. 2005, 233, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Blume, K.G.; Kaplan, J.C.; Löhr, G.W.; Ramot, B.; Valentine, W.N. International Committee for Standardization in Haematology: Recommended Methods for Red-Cell Enzyme Analysis. Br. J. Haematol. 1977, 35, 331–340. [Google Scholar] [CrossRef]
- Werre, J.M.; Willekens, F.L.A.; Bosch, F.H.; de Haans, L.D.; van der Vegt, S.G.L.; van den Bos, A.G.; Bosman, G.J.C.G.M. The Red Cell Revisited—Matters of Life and Death. Cell. Mol. Biol. 2004, 50, 139–145. [Google Scholar]
- Bryk, A.H.; Wiśniewski, J.R. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 2017, 16, 2752–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Recommended Methods for an Additional Red Cell Enzyme (Pyrimidine 5′-Nucleotidase) Assay and the Determination of Red Cell Adenosine-5′-Triphosphate, 2,3-Diphosphoglycerate and Reduced Glutathione. International Committee for Standardization in Haematology. Clin. Lab. Haematol. 1989, 11, 131–138. [CrossRef] [PubMed]
- Bosman, G.J.C.G.M.; Lasonder, E.; Groenen-Döpp, Y.A.M.; Willekens, F.L.A.; Werre, J.M.; Novotný, V.M.J. Comparative Proteomics of Erythrocyte Aging in Vivo and in Vitro. J. Proteom. 2010, 73, 396–402. [Google Scholar] [CrossRef]
- O’Neill, J.S.; Reddy, A.B. Circadian Clocks in Human Red Blood Cells. Nature 2011, 469, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, R.F.; Ledue, T.B.; Craig, W.Y. Patient Hydration: A Major Source of Laboratory Uncertainty. Clin. Chem. Lab. Med. 2007, 45, 158–166. [Google Scholar] [CrossRef]
- Bouix, D.; Peyreigne, C.; Raynaud, E.; Monnier, J.F.; Micallef, J.P.; Brun, J.F. Relationships among Body Composition, Hemorheology and Exercise Performance in Rugbymen. Clin. Hemorheol. Microcirc. 1998, 19, 245–254. [Google Scholar]
- Lima-Oliveira, G.; Salvagno, G.L.; Lippi, G.; Gelati, M.; Montagnana, M.; Danese, E.; Picheth, G.; Guidi, G.C. Influence of a Regular, Standardized Meal on Clinical Chemistry Analytes. Ann. Lab. Med. 2012, 32, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.M.; Abazyan, B.; Ono, M.; Hogue, C.W.; Cohen, D.B.; Berkowitz, D.E.; Ness, P.M.; Barodka, V.M. Decreased Erythrocyte Deformability after Transfusion and the Effects of Erythrocyte Storage Duration. Anesth. Analg. 2013, 116, 975–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muravyov, A.V.; Yakusevich, V.V.; Chuchkanov, F.A.; Maimistova, A.A.; Bulaeva, S.V.; Zaitsev, L.G. Hemorheological Efficiency of Drugs, Targeting on Intracellular Phosphodiesterase Activity: In Vitro Study. Clin. Hemorheol. Microcirc. 2007, 36, 327–334. [Google Scholar] [PubMed]
- Maw, G.J.; Mackenzie, I.L.; Taylor, N.A. Human Body-Fluid Distribution during Exercise in Hot, Temperate and Cool Environments. Acta Physiol. Scand. 1998, 163, 297–304. [Google Scholar] [CrossRef]
- Banfi, G.; Salvagno, G.L.; Lippi, G. The Role of Ethylenediamine Tetraacetic Acid (EDTA) as in Vitro Anticoagulant for Diagnostic Purposes. Clin. Chem. Lab. Med. 2007, 45, 565–576. [Google Scholar] [CrossRef]
- Boyanton, B.L.; Blick, K.E. Stability Studies of Twenty-Four Analytes in Human Plasma and Serum. Clin. Chem. 2002, 48, 2242–2247. [Google Scholar] [CrossRef] [PubMed]
- Mohri, M.; Rezapoor, H. Effects of Heparin, Citrate, and EDTA on Plasma Biochemistry of Sheep: Comparison with Serum. Res. Vet. Sci. 2009, 86, 111–114. [Google Scholar] [CrossRef]
- Torrance, J.D.; Lenfant, C. Methods for Determination of O2 Dissociation Curves, Including Bohr Effect. Respir. Physiol. 1969, 8, 127–136. [Google Scholar] [CrossRef]
- Baskurt, O.K.; Boynard, M.; Cokelet, G.C.; Connes, P.; Cooke, B.M.; Forconi, S.; Liao, F.; Hardeman, M.R.; Jung, F.; Meiselman, H.J.; et al. New Guidelines for Hemorheological Laboratory Techniques. Clin. Hemorheol. Microcirc. 2009, 42, 75–97. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Diagnostic Imaging and Laboratory Technology. In Use of Anticoagulants in Diagnostic Laboratory Investigations. Stability of Blood, Plasma and Serum Samples; World Health Organization: Geneva, Switzerland, 2002; p. 62. [Google Scholar]
- Park, J.-S.; Kim, S.-J.; Lee, S.-W.; Lee, E.-J.; Han, K.-S.; Moon, S.-W.; Hong, Y.-S. Initial Low Oxygen Extraction Ratio Is Related to Severe Organ Dysfunction and High In-Hospital Mortality in Severe Sepsis and Septic Shock Patients. J. Emerg. Med. 2015, 49, 261–267. [Google Scholar] [CrossRef]
- Erecińska, M.; Silver, I.A. ATP and Brain Function. J. Cereb. Blood Flow Metab. 1989, 9, 2–19. [Google Scholar] [CrossRef] [Green Version]
- Leithner, C.; Royl, G. The Oxygen Paradox of Neurovascular Coupling. J. Cereb. Blood Flow Metab. 2014, 34, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, C.N.; Klein-Flügge, M.C.; Howarth, C.; Attwell, D. Oxidative Phosphorylation, Not Glycolysis, Powers Presynaptic and Postsynaptic Mechanisms Underlying Brain Information Processing. J. Neurosci. 2012, 32, 8940–8951. [Google Scholar] [CrossRef]
- Bailey, D.M.; Willie, C.K.; Hoiland, R.L.; Bain, A.R.; MacLeod, D.B.; Santoro, M.A.; DeMasi, D.K.; Andrijanic, A.; Mijacika, T.; Barak, O.F.; et al. Surviving Without Oxygen: How Low Can the Human Brain Go? High Alt. Med. Biol. 2017, 18, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Sanap, M.N.; Worthley, L.I.G. Neurologic Complications of Critical Illness: Part I. Altered States of Consciousness and Metabolic Encephalopathies. Crit. Care Resusc. 2002, 4, 119–132. [Google Scholar] [PubMed]
- Berisavac, I.I.; Jovanović, D.R.; Padjen, V.V.; Ercegovac, M.D.; Stanarčević, P.D.J.; Budimkić-Stefanović, M.S.; Radović, M.M.; Beslać-Bumbaširević, L.G. How to Recognize and Treat Metabolic Encephalopathy in Neurology Intensive Care Unit. Neurol. India 2017, 65, 123–128. [Google Scholar] [CrossRef]
- Moroney, J.T.; Bagiella, E.; Desmond, D.W.; Paik, M.C.; Stern, Y.; Tatemichi, T.K. Risk Factors for Incident Dementia after Stroke. Role of Hypoxic and Ischemic Disorders. Stroke 1996, 27, 1283–1289. [Google Scholar] [CrossRef]
- Raz, L.; Knoefel, J.; Bhaskar, K. The Neuropathology and Cerebrovascular Mechanisms of Dementia. J. Cereb. Blood Flow Metab. 2016, 36, 172–186. [Google Scholar] [CrossRef] [Green Version]
- Peers, C.; Dallas, M.L.; Boycott, H.E.; Scragg, J.L.; Pearson, H.A.; Boyle, J.P. Hypoxia and Neurodegeneration. Ann. N. Y. Acad. Sci. 2009, 1177, 169–177. [Google Scholar] [CrossRef]
- Carlson, B.W.; Neelon, V.J.; Carlson, J.R.; Hartman, M.; Bliwise, D.L. Cerebral Oxygenation in Wake and during Sleep and Its Relationship to Cognitive Function in Community-Dwelling Older Adults without Sleep Disordered Breathing. J. Gerontol. A Biol. Sci. Med. Sci. 2011, 66, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Yaffe, K.; Laffan, A.M.; Harrison, S.L.; Redline, S.; Spira, A.P.; Ensrud, K.E.; Ancoli-Israel, S.; Stone, K.L. Sleep-Disordered Breathing, Hypoxia, and Risk of Mild Cognitive Impairment and Dementia in Older Women. JAMA 2011, 306, 613–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carelli-Alinovi, C.; Misiti, F. Erythrocytes as Potential Link between Diabetes and Alzheimer’s Disease. Front. Aging Neurosci. 2017, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Pańkowska, E.; Szypowska, A.; Wysocka, M.; Lipka, M. The role of 2,3-DPG in nerve conduction of children with type 1 diabetes. Endokrynol. Diabetol. Chor. Przemiany Mater. Wieku Rozw 2005, 11, 207–210. [Google Scholar]
- Barshtein, G.; Ponizovsky, A.M.; Nechamkin, Y.; Ritsner, M.; Yedgar, S.; Bergelson, L.D. Aggregability of Red Blood Cells of Schizophrenia Patients with Negative Syndrome Is Selectively Enhanced. Schizophr. Bull. 2004, 30, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.-Y.; Liang, H.-J.; Chow, S.-Y.; Chen, S.-M.; Liu, D.-Z. Hemorheological Mechanisms in Alzheimer’s Disease. Microcirculation 2007, 14, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Wojsiat, J.; Laskowska-Kaszub, K.; Mietelska-Porowska, A.; Wojda, U. Search for Alzheimer’s Disease Biomarkers in Blood Cells: Hypotheses-Driven Approach. Biomark. Med. 2017, 11, 917–931. [Google Scholar] [CrossRef] [Green Version]
- de la Torre, J.C. Alzheimer Disease as a Vascular Disorder. Nosological Evidence. Stroke 2002, 33, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Prohaska, R.; Sibon, O.C.M.; Rudnicki, D.D.; Danek, A.; Hayflick, S.J.; Verhaag, E.M.; Vonk, J.J.; Margolis, R.L.; Walker, R.H. Brain, Blood, and Iron: Perspectives on the Roles of Erythrocytes and Iron in Neurodegeneration. Neurobiol. Dis. 2012, 46, 607–624. [Google Scholar] [CrossRef] [Green Version]
- Weuve, J.; Mendes de Leon, C.F.; Bennett, D.A.; Dong, X.; Evans, D.A. The Red Cell Distribution Width and Anemia in Association with Prevalent Dementia. Alzheimer Dis. Assoc. Disord. 2014, 28, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, V.L.; Hardie, R.J. Acanthocytosis and Neurological Disorders. J. Neurol. 2001, 248, 87–94. [Google Scholar] [CrossRef]
- Lin, J.E.; Asfour, A.; Sewell, T.B.; Hooe, B.; Pryce, P.; Earley, C.; Shen, M.Y.; Kerner-Rossi, M.; Thakur, K.T.; Vargas, W.S.; et al. Neurological Issues in Children with COVID-19. Neurosci. Lett. 2021, 743, 135567. [Google Scholar] [CrossRef]
- Chen, X.; Laurent, S.; Onur, O.A.; Kleineberg, N.N.; Fink, G.R.; Schweitzer, F.; Warnke, C. A Systematic Review of Neurological Symptoms and Complications of COVID-19. J. Neurol. 2021, 268, 392–402. [Google Scholar] [CrossRef]
- Román, G.C.; Spencer, P.S.; Reis, J.; Buguet, A.; Faris, M.E.A.; Katrak, S.M.; Láinez, M.; Medina, M.T.; Meshram, C.; Mizusawa, H.; et al. The Neurology of COVID-19 Revisited: A Proposal from the Environmental Neurology Specialty Group of the World Federation of Neurology to Implement International Neurological Registries. J. Neurol. Sci. 2020, 414, 116884. [Google Scholar] [CrossRef]
- Shappell, S.D.; Murray, J.A.; Nasser, M.G.; Wills, R.E.; Torrance, J.D.; Lenfant, C.J. Acute Change in Hemoglobin Affinity for Oxygen during Angina Pectoris. N. Engl. J. Med. 1970, 282, 1219–1224. [Google Scholar] [CrossRef]
- Kinoshita, A.; Tsukada, K.; Soga, T.; Hishiki, T.; Ueno, Y.; Nakayama, Y.; Tomita, M.; Suematsu, M. Roles of Hemoglobin Allostery in Hypoxia-Induced Metabolic Alterations in Erythrocytes: Simulation and Its Verification by Metabolome Analysis. J. Biol. Chem. 2007, 282, 10731–10741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böning, D.; Enciso, G. Hemoglobin-Oxygen Affinity in Anemia. Blut 1987, 54, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Torrance, J.; Jacobs, P.; Restrepo, A.; Eschbach, J.; Lenfant, C.; Finch, C.A. Intraerythrocytic Adaptation to Anemia. N. Engl. J. Med. 1970, 283, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Valeri, C.R.; Fortier, N.L. Red-Cell 2,3-Diphosphoglycerate and Creatine Levels in Patients with Red-Cell Mass Deficits or with Cardiopulmonary Insufficiency. N. Engl. J. Med. 1969, 281, 1452–1455. [Google Scholar] [CrossRef]
- Kashani, K.B. Hypoxia in COVID-19: Sign of Severity or Cause for Poor Outcomes. Mayo Clin. Proc. 2020, 95, 1094–1096. [Google Scholar] [CrossRef]
- Ghebrehiwet, B.; Peerschke, E.I. Complement and Coagulation: Key Triggers of COVID-19-Induced Multiorgan Pathology. J. Clin. Investig. 2020, 130, 5674–5676. [Google Scholar] [CrossRef]
- Abboud, H.; Abboud, F.Z.; Kharbouch, H.; Arkha, Y.; El Abbadi, N.; El Ouahabi, A. COVID-19 and SARS-Cov-2 Infection: Pathophysiology and Clinical Effects on the Nervous System. World Neurosurg. 2020, 140, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.M.; Chiu, D.T.; Yee, M.C.; Lubin, B.H. Red Cell Vesiculation—A Common Membrane Physiologic Event. J. Lab. Clin. Med. 1986, 108, 315–324. [Google Scholar] [PubMed]
- Leonard, C.; Conrard, L.; Guthmann, M.; Pollet, H.; Carquin, M.; Vermylen, C.; Gailly, P.; Van Der Smissen, P.; Mingeot-Leclercq, M.P.; Tyteca, D. Contribution of Plasma Membrane Lipid Domains to Red Blood Cell (Re)Shaping. Sci. Rep. 2017, 7, 4264. [Google Scholar] [CrossRef] [PubMed]
- Chasis, J.A.; Mohandas, N. Erythrocyte Membrane Deformability and Stability: Two Distinct Membrane Properties That Are Independently Regulated by Skeletal Protein Associations. J. Cell. Biol. 1986, 103, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Cluitmans, J.C.A.; Hardeman, M.R.; Dinkla, S.; Brock, R.; Bosman, G.J.C.G.M. Red Blood Cell Deformability during Storage: Towards Functional Proteomics and Metabolomics in the Blood Bank. Blood Transfus 2012, 10 (Suppl. S2), s12–s18. [Google Scholar] [CrossRef]
- Grau, M.; Pauly, S.; Ali, J.; Walpurgis, K.; Thevis, M.; Bloch, W.; Suhr, F. RBC-NOS-Dependent S-Nitrosylation of Cytoskeletal Proteins Improves RBC Deformability. PLoS ONE 2013, 8, e56759. [Google Scholar] [CrossRef] [Green Version]
- Kuypers, F.A.; Scott, M.D.; Schott, M.A.; Lubin, B.; Chiu, D.T. Use of Ektacytometry to Determine Red Cell Susceptibility to Oxidative Stress. J. Lab. Clin. Med. 1990, 116, 535–545. [Google Scholar] [PubMed]
- Kim, D.H.; Kim, Y.K.; Won, D.I.; Shin, S.; Suh, J.S. Assessment of hemorheological deformability of human red cells exposed to tert-butyl hydroperoxide, verapamil and ascorbate by ektacytometer. Korean J. Lab. Med. 2008, 28, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, R.J.; Machiedo, G.W.; Rush, B.F.; Dikdan, G. Oxygen Free Radicals: Effect on Red Cell Deformability in Sepsis. Crit. Care Med. 1991, 19, 732–735. [Google Scholar] [CrossRef]
- Sivilotti, M.L.A. Oxidant Stress and Haemolysis of the Human Erythrocyte. Toxicol. Rev. 2004, 23, 169–188. [Google Scholar] [CrossRef]
- Li, H.; Lu, L.; Li, X.; Buffet, P.A.; Dao, M.; Karniadakis, G.E.; Suresh, S. Mechanics of Diseased Red Blood Cells in Human Spleen and Consequences for Hereditary Blood Disorders. Proc. Natl. Acad. Sci. USA 2018, 115, 9574–9579. [Google Scholar] [CrossRef] [Green Version]
- Hébert, P.C.; Hu, L.Q.; Biro, G.P.; Carter, A. Review of Physiologic Mechanisms in Response to Anemia. Can. Med. Assoc. J. 1997, 156, S27–S40. [Google Scholar]
- Kubánková, M.; Hohberger, B.; Hoffmanns, J.; Fürst, J.; Herrmann, M.; Guck, J.; Kräter, M. Physical Phenotype of Blood Cells Is Altered in COVID-19. Biophys. J. 2021, 120, 2838–2847. [Google Scholar] [CrossRef] [PubMed]
- Revin, V.V.; Balykova, L.A.; Radaeva, O.A.; Shchapov, V.V.; Revina, E.S.; Pinyaev, S.I.; Kostina, Y.A.; Kozlov, E.D. Morphofunctional Characteristics of Erythrocytes and Blood Erythropoietin Level in Patients as Predictors of Severe Course of COVID-19. Bull. Exp. Biol. Med. 2022, 173, 46–50. [Google Scholar] [CrossRef]
- Mahdi, A.; Collado, A.; Tengbom, J.; Jiao, T.; Wodaje, T.; Johansson, N.; Farnebo, F.; Färnert, A.; Yang, J.; Lundberg, J.O.; et al. Erythrocytes Induce Vascular Dysfunction in COVID-19. JACC Basic Transl. Sci. 2022, 7, 193–204. [Google Scholar] [CrossRef]
- Yedgar, S.; Koshkaryev, A.; Barshtein, G. The Red Blood Cell in Vascular Occlusion. Pathophysiol. Haemost. Thromb. 2002, 32, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Lipowsky, H.H. Microvascular Rheology and Hemodynamics. Microcirculation 2005, 12, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Scholkmann, F.; Restin, T.; Ferrari, M.; Quaresima, V. The Role of Methemoglobin and Carboxyhemoglobin in COVID-19: A Review. J. Clin. Med. 2020, 10, 50. [Google Scholar] [CrossRef]
- Kashari, O.F.; Alsamiri, S.A.; Zabbani, F.M.; Musalli, D.I.; Ibrahim, A.M. Occurrence of Methemoglobinemia Due to COVID-19: A Case Report. Cureus 2022, 14, e23155. [Google Scholar] [CrossRef]
- Ting, Y.L.; Lim, J.Z.M.; Yeo, P.M.; Sim, W. Methemoglobinemia: A Potential Confounder in COVID-19 Respiratory Failure. Singap. Med. J. 2021, 1–6. [Google Scholar] [CrossRef]
- Sahu, K.K.; Mishra, A.K.; Mishra, K. Methemoglobinemia in COVID-19. Am. J. Med. Sci. 2021, 362, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.V.; Lazar Neto, F.; Marques, L.C.; Lima, R.B.O.; Brandão, A.A.G.S. Methemoglobinemia and Hemolytic Anemia after COVID-19 Infection without Identifiable Eliciting Drug: A Case-Report. IDCases 2021, 23, e01013. [Google Scholar] [CrossRef] [PubMed]
- Naymagon, L.; Berwick, S.; Kessler, A.; Lancman, G.; Gidwani, U.; Troy, K. The Emergence of Methemoglobinemia amidst the COVID-19 Pandemic. Am. J. Hematol. 2020, 95, E196–E197. [Google Scholar] [CrossRef]
- Rifkind, J.M.; Mohanty, J.G.; Nagababu, E. The Pathophysiology of Extracellular Hemoglobin Associated with Enhanced Oxidative Reactions. Front. Physiol. 2014, 5, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, R.M.; Sharpe, M.D.; Singer, M.; Ellis, C.G. The Effect of Sepsis on the Erythrocyte. Int. J. Mol. Sci. 2017, 18, 1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohorović, L.; Lavezzi, A.M.; Jonjić, N.; Štifter, S.; Perry, G.; Malatestinić, Đ.; Mićović, V.; Materljan, E.; Haller, H.; Petrović, O. Methemoglobinemia as Biomarker and Precursor of Brain Capillary Oxidative Damage Link to Ferric Iron Accumulation and Originator of Neurodegenerative Disease. J. Syst. Integr. Neurosci. 2017, 3, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Wright, R.O.; Lewander, W.J.; Woolf, A.D. Methemoglobinemia: Etiology, Pharmacology, and Clinical Management. Ann. Emerg. Med. 1999, 34, 646–656. [Google Scholar] [CrossRef]
- Jaffé, E.R. Methemoglobin Pathophysiology. Prog. Clin. Biol. Res. 1981, 51, 133–151. [Google Scholar]
- Skold, A.; Cosco, D.L.; Klein, R. Methemoglobinemia: Pathogenesis, Diagnosis, and Management. South. Med. J. 2011, 104, 757–761. [Google Scholar] [CrossRef]
- Russo, A.; Tellone, E.; Barreca, D.; Ficarra, S.; Laganà, G. Implication of COVID-19 on Erythrocytes Functionality: Red Blood Cell Biochemical Implications and Morpho-Functional Aspects. Int. J. Mol. Sci. 2022, 23, 2171. [Google Scholar] [CrossRef]
- Khakwani, M.; Horgan, C.; Ewing, J. COVID-19-Associated Oxidative Damage to Red Blood Cells. Br. J. Haematol. 2021, 193, 481. [Google Scholar] [CrossRef] [PubMed]
- Berzuini, A.; Bianco, C.; Migliorini, A.C.; Maggioni, M.; Valenti, L.; Prati, D. Red Blood Cell Morphology in Patients with COVID-19-Related Anaemia. Blood Transfus 2021, 19, 34–36. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, E.; Bagherpour, Z.; Zarei, E.; Omidkhoda, A. Pathological Effects of SARS-CoV-2 on Hematological and Immunological Cells: Alterations in Count, Morphology, and Function. Pathol. Res. Pract. 2022, 231, 153782. [Google Scholar] [CrossRef]
- Janz, D.R.; Ware, L.B. The Role of Red Blood Cells and Cell-Free Hemoglobin in the Pathogenesis of ARDS. J. Intensive Care 2015, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- De Franceschi, L.; Bosman, G.J.C.G.M.; Mohandas, N. Abnormal Red Cell Features Associated with Hereditary Neurodegenerative Disorders: The Neuroacanthocytosis Syndromes. Curr. Opin. Hematol. 2014, 21, 201–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadanand, A.; Janardhanan, A.; Sankaradoss, A.; Vanisree, A.J.; Arulnambi, T.; Bhanu, K. Erythrocyte Membrane in the Evaluation of Neurodegenerative Disorders. Degener. Neurol. Neuromuscul. Dis. 2017, 7, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, D.A.; Markesbery, W.R. Specificity of Biophysical and Biochemical Alterations in Erythrocyte Membranes in Neurological Disorders—Huntington’s Disease, Friedreich’s Ataxia, Alzheimer’s Disease, Amyotrophic Lateral Sclerosis, and Myotonic and Duchenne Muscular Dystrophy. J. Neurol. Sci. 1980, 47, 261–271. [Google Scholar] [CrossRef]
- Bosman, G.J.; Bartholomeus, I.G.; de Grip, W.J. Alzheimer’s Disease and Cellular Aging: Membrane-Related Events as Clues to Primary Mechanisms. Gerontology 1991, 37, 95–112. [Google Scholar] [CrossRef]
- Kaminsky, Y.; Poghosyan, A.; Tikhonova, L.; Palacios, H.H.; Kamal, M.A.; Kosenko, E.; Aliev, G. Glycolytic and Proteolytic Metabolism in Erythrocytes from Elderly and Demented Patients. Am. J. Neuroprotec. Neuroregen. 2012, 4, 73–77. [Google Scholar] [CrossRef]
- Repetto, M.G.; Reides, C.G.; Evelson, P.; Kohan, S.; de Lustig, E.S.; Llesuy, S.F. Peripheral Markers of Oxidative Stress in Probable Alzheimer Patients. Eur. J. Clin. Investig. 1999, 29, 643–649. [Google Scholar] [CrossRef]
- Vural, H.; Demirin, H.; Kara, Y.; Eren, I.; Delibas, N. Alterations of Plasma Magnesium, Copper, Zinc, Iron and Selenium Concentrations and Some Related Erythrocyte Antioxidant Enzyme Activities in Patients with Alzheimer’s Disease. J. Trace. Elem. Med. Biol. 2010, 24, 169–173. [Google Scholar] [CrossRef]
- Gibson, G.E.; Huang, H.-M. Oxidative Processes in the Brain and Non-Neuronal Tissues as Biomarkers of Alzheimer’s Disease. Front. Biosci. 2002, 7, d1007–d1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, J.M.; Matsuyama, S.S.; Meier, K.; Jarvik, L.F. Elevation of Erythrocyte Countertransport Rates in Alzheimer’s Dementia. N. Engl. J. Med. 1983, 309, 1061–1062. [Google Scholar] [PubMed]
- Ronquist, G.; Waldenström, A. Imbalance of Plasma Membrane Ion Leak and Pump Relationship as a New Aetiological Basis of Certain Disease States. J. Intern. Med. 2003, 254, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Kawamoto, E.M.; Munhoz, C.D.; Glezer, I.; Bahia, V.S.; Caramelli, P.; Nitrini, R.; Gorjão, R.; Curi, R.; Scavone, C.; Marcourakis, T. Oxidative State in Platelets and Erythrocytes in Aging and Alzheimer’s Disease. Neurobiol. Aging 2005, 26, 857–864. [Google Scholar] [CrossRef]
- Dinkla, S.; van Eijk, L.T.; Fuchs, B.; Schiller, J.; Joosten, I.; Brock, R.; Pickkers, P.; Bosman, G.J.C.G.M. Inflammation-Associated Changes in Lipid Composition and the Organization of the Erythrocyte Membrane. BBA Clin. 2016, 5, 186–192. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, A.; Nemkov, T.; Sun, K.; Liu, H.; Song, A.; Monte, A.A.; Subudhi, A.W.; Lovering, A.T.; Dvorkin, D.; Julian, C.G.; et al. AltitudeOmics: Red Blood Cell Metabolic Adaptation to High Altitude Hypoxia. J. Proteome Res. 2016, 15, 3883–3895. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, J.G.; Eckley, D.M.; Williamson, J.D.; Launer, L.J.; Rifkind, J.M. Do Red Blood Cell-Beta-Amyloid Interactions Alter Oxygen Delivery in Alzheimer’s Disease? Adv. Exp. Med. Biol. 2008, 614, 29–35. [Google Scholar] [CrossRef]
- Humpeler, E.; Vogel, S.; Schobersberger, W.; Mairbäurl, H. Red Cell Oxygen Transport in Man in Relation to Gender and Age. Mech. Ageing Dev. 1989, 47, 229–239. [Google Scholar] [CrossRef]
- Bosman, G.J.; Janzing, J.G.; Bartholomeus, I.G.; De Man, A.J.; Zitman, F.G.; De Grip, W.J. Erythrocyte Aging Characteristics in Elderly Individuals with Beginning Dementia. Neurobiol. Aging 1997, 18, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Kosenko, E.A.; Tikhonova, L.A.; Pogosyan, A.S.; Kaminsky, Y.G. Erythrocyte Antioxidants in Aging and Dementia. Biochem. Mosc. Suppl. Ser. B 2012, 6, 273–277. [Google Scholar] [CrossRef]
- Rabie, T.; Kunze, R.; Marti, H.H. Impaired Hypoxic Response in Senescent Mouse Brain. Int. J. Dev. Neurosci. 2011, 29, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Ratcliffe, S.J.; Knoll, F.; Wu, J.; Ances, B.; Mardini, W.; Floyd, T.F. Aging: Impact upon Local Cerebral Oxygenation and Blood Flow with Acute Isovolemic Hemodilution. J. Neurosurg. Anesthesiol. 2006, 18, 125–131. [Google Scholar] [CrossRef]
- Anai, M.; Akaike, K.; Iwagoe, H.; Akasaka, T.; Higuchi, T.; Miyazaki, A.; Naito, D.; Tajima, Y.; Takahashi, H.; Komatsu, T.; et al. Decrease in Hemoglobin Level Predicts Increased Risk for Severe Respiratory Failure in COVID-19 Patients with Pneumonia. Respir. Investig. 2021, 59, 187–193. [Google Scholar] [CrossRef]
- Bellmann-Weiler, R.; Lanser, L.; Barket, R.; Rangger, L.; Schapfl, A.; Schaber, M.; Fritsche, G.; Wöll, E.; Weiss, G. Prevalence and Predictive Value of Anemia and Dysregulated Iron Homeostasis in Patients with COVID-19 Infection. J. Clin. Med. 2020, 9, 2429. [Google Scholar] [CrossRef] [PubMed]
- Klok, F.A.; Kruip, M.J.H.A.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.A.M.P.J.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Incidence of Thrombotic Complications in Critically Ill ICU Patients with COVID-19. Thromb. Res. 2020, 191, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Seyhan, E.C.; Özgül, M.A.; Tutar, N.; Ömür, I.; Uysal, A.; Altin, S. Red Blood Cell Distribution and Survival in Patients with Chronic Obstructive Pulmonary Disease. COPD 2013, 10, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.; Saad, K.; Elgohary, G.; AbdElHaffez, A.; El-Aziz, N.A. Is COVID-19 a Systemic Disease? Coronaviruses 2021, 2, 4–8. [Google Scholar] [CrossRef]
- Ponti, G.; Maccaferri, M.; Ruini, C.; Tomasi, A.; Ozben, T. Biomarkers Associated with COVID-19 Disease Progression. Crit. Rev. Clin. Lab. Sci. 2020, 57, 389–399. [Google Scholar] [CrossRef]
- Bonaventura, A.; Vecchié, A.; Dagna, L.; Martinod, K.; Dixon, D.L.; Van Tassell, B.W.; Dentali, F.; Montecucco, F.; Massberg, S.; Levi, M.; et al. Endothelial Dysfunction and Immunothrombosis as Key Pathogenic Mechanisms in COVID-19. Nat. Rev. Immunol. 2021, 21, 319–329. [Google Scholar] [CrossRef]
- Zeng, F.; Huang, Y.; Guo, Y.; Yin, M.; Chen, X.; Xiao, L.; Deng, G. Association of Inflammatory Markers with the Severity of COVID-19: A Meta-Analysis. Int. J. Infect. Dis. 2020, 96, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Caricchio, R.; Gallucci, M.; Dass, C.; Zhang, X.; Gallucci, S.; Fleece, D.; Bromberg, M.; Criner, G.J. Temple University COVID-19 Research Group Preliminary Predictive Criteria for COVID-19 Cytokine Storm. Ann. Rheum. Dis. 2021, 80, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An Inflammatory Cytokine Signature Predicts COVID-19 Severity and Survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; Remigante, A.; Cordaro, M.; Trichilo, V.; Loddo, S.; Dossena, S.; Marino, A. Impact of Acute Inflammation on Band 3 Protein Anion Exchange Capability in Human Erythrocytes. Arch. Physiol. Biochem. 2022, 128, 1242–1248. [Google Scholar] [CrossRef]
- Straat, M.; van Bruggen, R.; de Korte, D.; Juffermans, N.P. Red Blood Cell Clearance in Inflammation. Transfus Med. Hemother. 2012, 39, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Reithmeier, R.A.F.; Casey, J.R.; Kalli, A.C.; Sansom, M.S.P.; Alguel, Y.; Iwata, S. Band 3, the Human Red Cell Chloride/Bicarbonate Anion Exchanger (AE1, SLC4A1), in a Structural Context. Biochim. Biophys. Acta 2016, 1858, 1507–1532. [Google Scholar] [CrossRef]
- Jensen, F.B. The Dual Roles of Red Blood Cells in Tissue Oxygen Delivery: Oxygen Carriers and Regulators of Local Blood Flow. J. Exp. Biol. 2009, 212, 3387–3393. [Google Scholar] [CrossRef] [Green Version]
- Pyle, C.J.; Uwadiae, F.I.; Swieboda, D.P.; Harker, J.A. Early IL-6 Signalling Promotes IL-27 Dependent Maturation of Regulatory T Cells in the Lungs and Resolution of Viral Immunopathology. PLoS Pathog. 2017, 13, e1006640. [Google Scholar] [CrossRef] [Green Version]
- Bester, J.; Pretorius, E. Effects of IL-1β, IL-6 and IL-8 on Erythrocytes, Platelets and Clot Viscoelasticity. Sci. Rep. 2016, 6, 32188. [Google Scholar] [CrossRef] [Green Version]
- Spickett, C.M.; Smith, W.E.; Reglinski, J.; Wilson, R.; Walker, J.J. Oxidation of Erythrocyte Glutathione by Monocytes Stimulated with Interleukin-6. Analysis by 1H Spin Echo NMR. Clin. Chim. Acta 1998, 270, 115–124. [Google Scholar] [CrossRef]
- Santos-Silva, A.; Castro, E.M.; Teixeira, N.A.; Guerra, F.C.; Quintanilha, A. Erythrocyte Membrane Band 3 Profile Imposed by Cellular Aging, by Activated Neutrophils and by Neutrophilic Elastase. Clin. Chim. Acta 1998, 275, 185–196. [Google Scholar] [CrossRef]
- Khan, M.; Shah, N.; Mushtaq, H.; Jehanzeb, V. Profiling Laboratory Biomarkers Associated with COVID-19 Disease Progression: A Single-Center Experience. Int. J. Microbiol. 2021, 2021, 6643333. [Google Scholar] [CrossRef] [PubMed]
- Mahat, R.K.; Panda, S.; Rathore, V.; Swain, S.; Yadav, L.; Sah, S.P. The Dynamics of Inflammatory Markers in Coronavirus Disease-2019 (COVID-19) Patients: A Systematic Review and Meta-Analysis. Clin. Epidemiol. Glob. Health 2021, 11, 100727. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Lu, L.; Cao, W.; Li, T. Hypothesis for Potential Pathogenesis of SARS-CoV-2 Infection-a Review of Immune Changes in Patients with Viral Pneumonia. Emerg. Microbes Infect. 2020, 9, 727–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, C.; Bell, L.N.; Liang, H.; Haykal, R.; Kaiksow, F.; Mazza, J.J.; Yale, S.H. Erythrocyte Sedimentation Rate and C-Reactive Protein Measurements and Their Relevance in Clinical Medicine. WMJ 2016, 115, 317–321. [Google Scholar] [PubMed]
- Alende-Castro, V.; Alonso-Sampedro, M.; Vazquez-Temprano, N.; Tuñez, C.; Rey, D.; García-Iglesias, C.; Sopeña, B.; Gude, F.; Gonzalez-Quintela, A. Factors Influencing Erythrocyte Sedimentation Rate in Adults: New Evidence for an Old Test. Medicine 2019, 98, e16816. [Google Scholar] [CrossRef] [PubMed]
- Fabry, T.L. Mechanism of Erythrocyte Aggregation and Sedimentation. Blood 1987, 70, 1572–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vink, H.; Wieringa, P.A.; Spaan, J.A. Evidence That Cell Surface Charge Reduction Modifes Capillary Red Cell Velocity-Flux Relationships in Hamster Cremaster Muscle. J. Physiol. 1995, 489 Pt 1, 193–201. [Google Scholar] [CrossRef]
- Oberleithner, H.; Wälte, M.; Kusche-Vihrog, K. Sodium Renders Endothelial Cells Sticky for Red Blood Cells. Front. Physiol. 2015, 6, 188. [Google Scholar] [CrossRef] [Green Version]
- Levi, M.; van der Poll, T. Inflammation and Coagulation. Crit. Care Med. 2010, 38, S26-34. [Google Scholar] [CrossRef]
- Nader, E.; Skinner, S.; Romana, M.; Fort, R.; Lemonne, N.; Guillot, N.; Gauthier, A.; Antoine-Jonville, S.; Renoux, C.; Hardy-Dessources, M.-D.; et al. Blood Rheology: Key Parameters, Impact on Blood Flow, Role in Sickle Cell Disease and Effects of Exercise. Front. Physiol. 2019, 10, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jubran, A. Pulse Oximetry. Intensive Care Med. 2004, 30, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Lowe, C.; Ghareeb, S.A.; Artinian, N.T. Pulse Oximetry in Adults. Am. J. Nurs. 2009, 109, 52–59; quiz 60. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.P.; Giuliano, K.; Chen, H.-M. Pulse Oximetry Revisited: “But His O(2) Sat Was Normal!”. Clin. Nurse Spec. 2006, 20, 268–272. [Google Scholar] [CrossRef]
- Faisal, H.; Ali, S.T.; Xu, J.; Nisar, T.; Sabawi, M.; Salazar, E.; Masud, F.N. Carboxyhemoglobinemia in Critically Ill Coronavirus Disease 2019 Patients. J. Clin. Med. 2021, 10, 2731. [Google Scholar] [CrossRef]
- Martini, A.L.; Carli, G.; Kiferle, L.; Piersanti, P.; Palumbo, P.; Morbelli, S.; Calcagni, M.L.; Perani, D.; Sestini, S. Time-Dependent Recovery of Brain Hypometabolism in Neuro-COVID-19 Patients. Eur. J. Nucl. Med. Mol. Imaging 2022, 50, 90–102. [Google Scholar] [CrossRef]
- Guedj, E.; Campion, J.Y.; Dudouet, P.; Kaphan, E.; Bregeon, F.; Tissot-Dupont, H.; Guis, S.; Barthelemy, F.; Habert, P.; Ceccaldi, M.; et al. 18F-FDG Brain PET Hypometabolism in Patients with Long COVID. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 2823–2833. [Google Scholar] [CrossRef]
- Morand, A.; Campion, J.-Y.; Lepine, A.; Bosdure, E.; Luciani, L.; Cammilleri, S.; Chabrol, B.; Guedj, E. Similar Patterns of [18F]-FDG Brain PET Hypometabolism in Paediatric and Adult Patients with Long COVID: A Paediatric Case Series. Eur. J. Nucl. Med. Mol. Imaging 2022, 49, 913–920. [Google Scholar] [CrossRef]
- Snyder, B.; Simone, S.M.; Giovannetti, T.; Floyd, T.F. Cerebral Hypoxia: Its Role in Age-Related Chronic and Acute Cognitive Dysfunction. Anesth. Analg. 2021, 132, 1502–1513. [Google Scholar] [CrossRef]
- Areza-Fegyveres, R.; Kairalla, R.A.; Carvalho, C.R.R.; Nitrini, R. Cognition and Chronic Hypoxia in Pulmonary Diseases. Dement. Neuropsychol. 2010, 4, 14–22. [Google Scholar] [CrossRef]
Figure 1.
Multifactorial pathogenesis of COVID-19-related neurological complications includes hyperinflammation and cytokine storm, resulting in structural and metabolic RBC abnormalities, hemodynamic disorders, aggregation, thrombosis, impairment of the oxygen-transport function, hypoxia, encephalopathy, AD-related symptoms, and cognitive deficits.
Figure 1.
Multifactorial pathogenesis of COVID-19-related neurological complications includes hyperinflammation and cytokine storm, resulting in structural and metabolic RBC abnormalities, hemodynamic disorders, aggregation, thrombosis, impairment of the oxygen-transport function, hypoxia, encephalopathy, AD-related symptoms, and cognitive deficits.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kosenko, E.; Tikhonova, L.; Alilova, G.; Montoliu, C. Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 5739. https://doi.org/10.3390/ijms24065739
AMA Style
Kosenko E, Tikhonova L, Alilova G, Montoliu C. Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer’s Disease. International Journal of Molecular Sciences. 2023; 24(6):5739. https://doi.org/10.3390/ijms24065739
Chicago/Turabian StyleKosenko, Elena, Lyudmila Tikhonova, Gubidat Alilova, and Carmina Montoliu. 2023. "Erythrocytes Functionality in SARS-CoV-2 Infection: Potential Link with Alzheimer’s Disease" International Journal of Molecular Sciences 24, no. 6: 5739. https://doi.org/10.3390/ijms24065739
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.