
Epstein–Barr Virus and Human Herpesvirus-6 Reactivation in Acute COVID-19 Patients

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Cohort
2.2. Patient Selection and Samples
2.3. EBV and HHV-6 Assays
2.4. Data Analysis
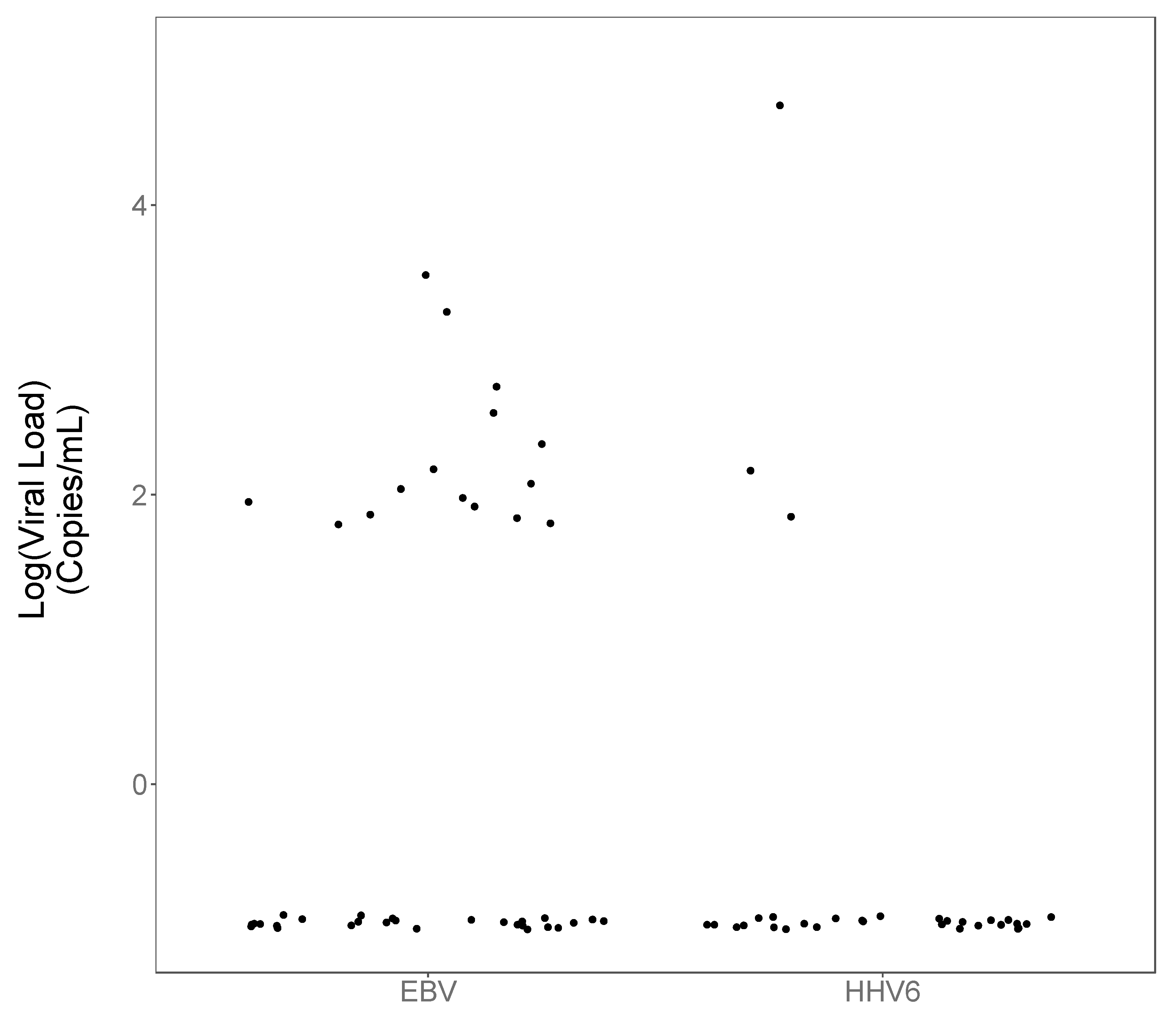
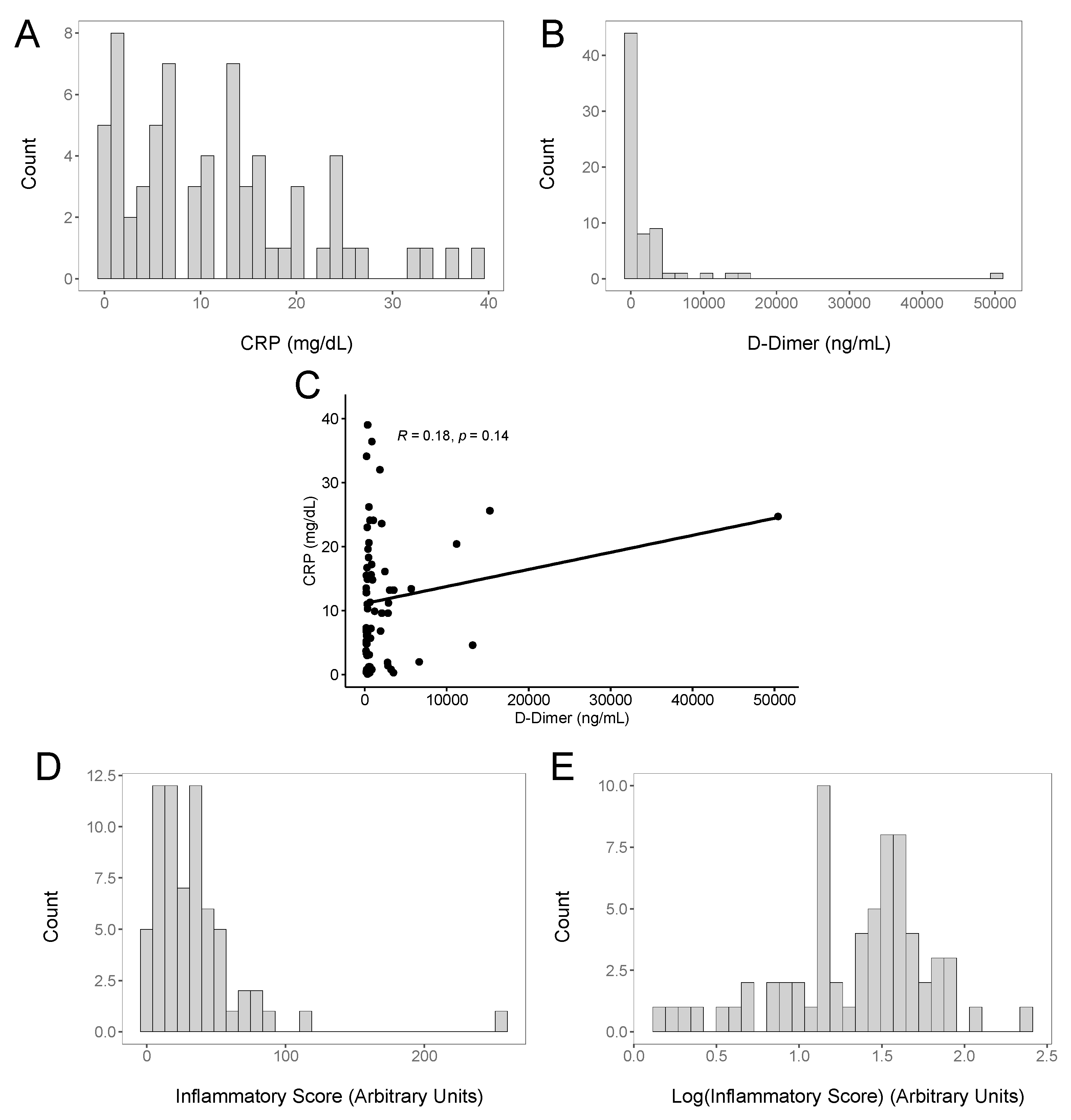
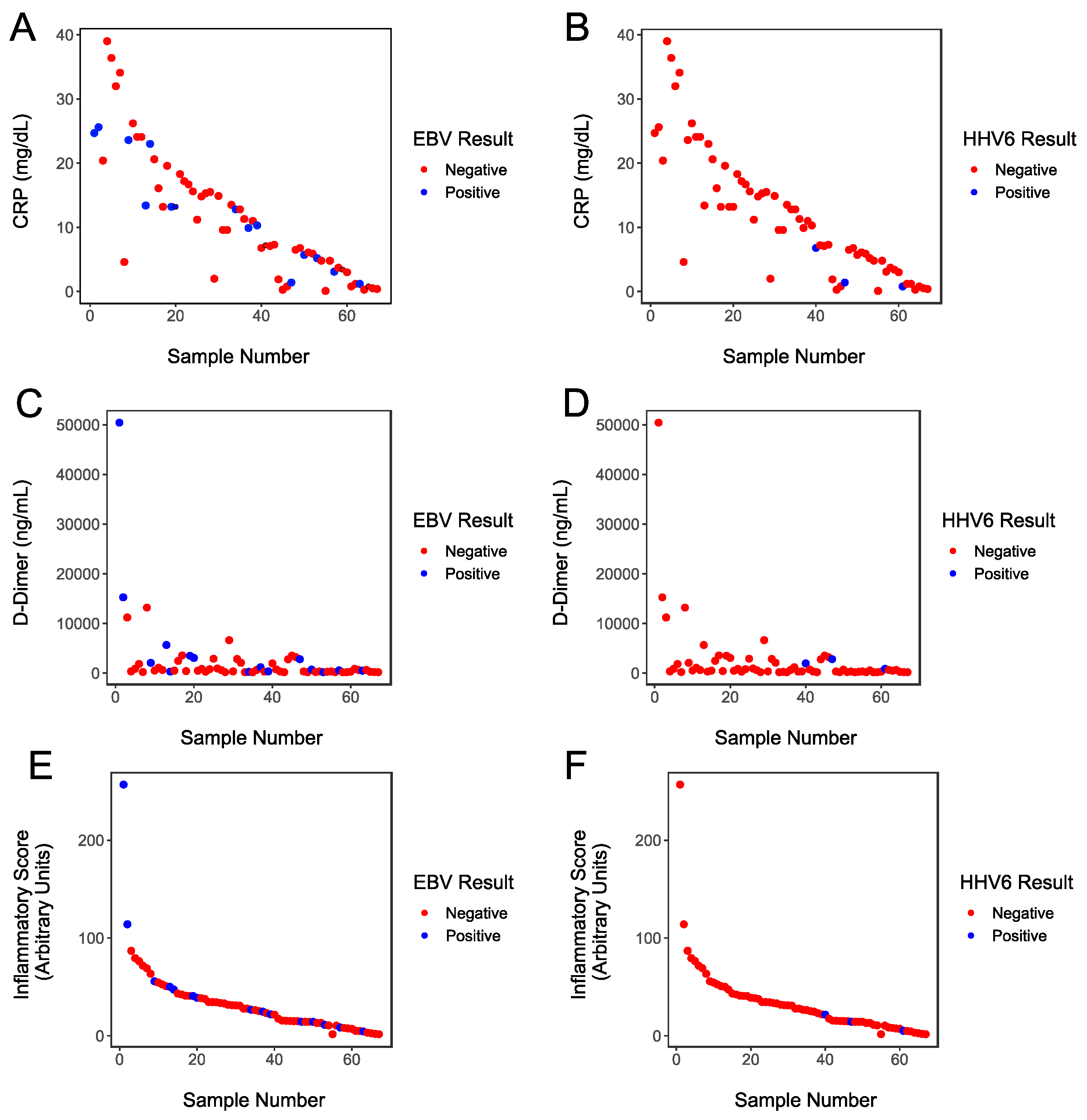
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyal, P.; Choi, J.J.; Pinheiro, L.C.; Schenck, E.J.; Chen, R.; Jabri, A.; Satlin, M.J.; Campion, T.R., Jr.; Nahid, M.; Ringel, J.B.; et al. Clinical Characteristics of Covid-19 in New York City. N. Engl. J. Med. 2020, 382, 2372–2374. [Google Scholar] [CrossRef]
- Fried, M.W.; Crawford, J.M.; Mospan, A.R.; Watkins, S.E.; Munoz, B.; Zink, R.C.; Elliott, S.; Burleson, K.; Landis, C.; Reddy, K.R.; et al. Patient Characteristics and Outcomes of 11,721 Patients With Coronavirus Disease 2019 (COVID-19) Hospitalized Across the United States. Clin. Infect. Dis. 2020, 72, e558–e565. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, S.; Kumar, D.; Rouse, B.T. Herpesviruses: Harmonious pathogens but relevant cofactors in other diseases? Front. Cell. Infect. Microbiol. 2018, 8, 177. [Google Scholar] [CrossRef]
- Strausbaugh, L.J.; Caserta, M.T.; Mock, D.J.; Dewhurst, S. Review, part 1: Human herpesvirus-6-basic biology, diagnostic testing, and antiviral efficacy. J. Med. Virol. 2010, 82, 1560–1568. [Google Scholar] [CrossRef]
- Caserta, M.T.; Mock, D.J.; Dewhurst, S. Human herpesvirus 6. Clin. Infect. Dis. 2001, 33, 829–833. [Google Scholar] [CrossRef]
- Faulkner, G.C.; Krajewski, A.S.; Crawford, D.H. The ins and outs of EBV infection. Trends Microbiol. 2000, 8, 185–189. [Google Scholar] [CrossRef]
- Amon, W.; Farrell, P.J. Reactivation of Epstein–Barr virus from latency. Rev. Med. Virol. 2005, 15, 149–156. [Google Scholar] [CrossRef]
- Cohen, J.I. Herpesvirus latency. J. Clin. Investig. 2020, 130, 3361–3369. [Google Scholar] [CrossRef]
- Tzellos, S.; Farrell, P.J. Epstein–Barr virus sequence variation—Biology and disease. Pathogens 2012, 1, 156–174. [Google Scholar] [CrossRef] [Green Version]
- Reusch, J.A.; Nawandar, D.M.; Wright, K.L.; Kenney, S.C.; Mertz, J.E. Cellular differentiation regulator BLIMP1 induces Epstein–Barr virus lytic reactivation in epithelial and B cells by activating transcription from both the R and Z promoters. J. Virol. 2015, 89, 1731–1743. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, D.; Takada, K. Phosphatidylinositol 3-kinase is a determinant of responsiveness to B cell antigen receptor-mediated Epstein–Barr virus activation. J. Immunol. 2004, 172, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Luka, J.; Kallin, B.; Klein, G. Induction of the Epstein–Barr virus (EBV) cycle in latently infected cells by n-butyrate. Virology 1979, 94, 228–231. [Google Scholar] [CrossRef]
- Choi, C.K.; Ho, D.N.; Hui, K.F.; Kao, R.Y.; Chiang, A.K. Identification of Novel Small Organic Compounds with Diverse Structures for the Induction of Epstein–Barr Virus (EBV) Lytic Cycle in EBV-Positive Epithelial Malignancies. PLoS ONE 2015, 10, e0145994. [Google Scholar] [CrossRef]
- Fahmi, H.; Cochet, C.; Hmama, Z.; Opolon, P.; Joab, I. Transforming growth factor beta 1 stimulates expression of the Epstein–Barr virus BZLF1 immediate-early gene product ZEBRA by an indirect mechanism which requires the MAPK kinase pathway. J. Virol. 2000, 74, 5810–5818. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Perrine, S.P.; Williams, R.M.; Faller, D.V. Histone deacetylase inhibitors are potent inducers of gene expression in latent EBV and sensitize lymphoma cells to nucleoside antiviral agents. Blood 2012, 119, 1008–1017. [Google Scholar] [CrossRef]
- Flemington, E.; Speck, S.H. Epstein–Barr virus BZLF1 trans activator induces the promoter of a cellular cognate gene, c-fos. J. Virol. 1990, 64, 4549–4552. [Google Scholar] [CrossRef]
- Hall, C.B.; Caserta, M.T.; Schnabel, K.C.; McDermott, M.P.; Lofthus, G.K.; Carnahan, J.A.; Gilbert, L.M.; Dewhurst, S. Characteristics and Acquisition of Human Herpesvirus (HHV)-7 Infections in Relation to Infection with HHV-6. J. Infect. Dis. 2006, 193, 1063–1069. [Google Scholar] [CrossRef]
- Yao, K.; Crawford, J.R.; Komaroff, A.L.; Ablashi, D.V.; Jacobson, S. Review part 2: Human herpesvirus-6 in central nervous system diseases. J. Med. Virol. 2010, 82, 1669–1678. [Google Scholar] [CrossRef]
- De Bolle, L.; Naesens, L.; De Clercq, E. Update on Human Herpesvirus 6 Biology, Clinical Features, and Therapy. Clin. Microbiol. Rev. 2005, 18, 217–245. [Google Scholar] [CrossRef] [Green Version]
- Ablashi, D.; Agut, H.; Álvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as distinct viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.L.; Pritchett, J.C.; Leifer, C.; Zerr, D.M.; Koelle, D.M.; Di Luca, D.; Lusso, P. HHV-6B infection, T-cell reconstitution, and graft-vs-host disease after hematopoietic stem cell transplantation. Bone Marrow Transplant. 2018, 53, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.B.; Long, C.E.; Schnabel, K.C.; Caserta, M.T.; McIntyre, K.M.; Costanzo, M.A.; Knott, A.; Dewhurst, S.; Insel, R.A.; Epstein, L.G. Human herpesvirus-6 infection in children—A prospective study of complications and reactivation. N. Engl. J. Med. 1994, 331, 432–438. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Naesens, L.; De Bolle, L.; Schols, D.; Zhang, Y.; Neyts, J. Antiviral agents active against human herpesviruses HHV-6, HHV-7 and HHV-8. Rev. Med. Virol. 2001, 11, 381–395. [Google Scholar] [CrossRef]
- Pellett, P.E.; Ablashi, D.V.; Ambros, P.F.; Agut, H.; Caserta, M.T.; Descamps, V.; Flamand, L.; Gautheret-Dejean, A.; Hall, C.B.; Kamble, R.T.; et al. Chromosomally integrated human herpesvirus 6: Questions and answers. Rev. Med. Virol. 2012, 22, 144–155. [Google Scholar] [CrossRef]
- Ogata, M.; Satou, T.; Kadota, J.-i.; Saito, N.; Yoshida, T.; Okumura, H.; Ueki, T.; Nagafuji, K.; Kako, S.; Uoshima, N.; et al. Human Herpesvirus 6 (HHV-6) Reactivation and HHV-6 Encephalitis After Allogeneic Hematopoietic Cell Transplantation: A Multicenter, Prospective Study. Clin. Infect. Dis. 2013, 57, 671–681. [Google Scholar] [CrossRef]
- Ichiche, M.; Kiesch, N.; De Bels, D. DRESS syndrome associated with HHV-6 reactivation. Eur. J. Intern. Med. 2003, 14, 498–500. [Google Scholar] [CrossRef]
- Ahluwalia, J.; Abuabara, K.; Perman, M.J.; Yan, A.C. Human herpesvirus 6 involvement in paediatric drug hypersensitivity syndrome. Br. J. Dermatol. 2015, 172, 1090–1095. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Fujita, A.; Yagami, A.; Suzuki, K.; Matsunaga, K.; Ihira, M.; Asano, Y. Human herpesvirus 6 reactivation and inflammatory cytokine production in patients with drug-induced hypersensitivity syndrome. J. Clin. Virol. 2006, 37 (Suppl. 1), S92–S96. [Google Scholar] [CrossRef]
- Kondo, K.; Yamanishi, K. HHV-6A, 6B, and 7: Molecular basis of latency and reactivation. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple early factors anticipate post-acute COVID-19 sequelae. Cell 2022, 185, 881–895.e20. [Google Scholar] [CrossRef]
- Zubchenko, S.; Kril, I.; Nadizhko, O.; Matsyura, O.; Chopyak, V. Herpesvirus infections and post-COVID-19 manifestations: A pilot observational study. Rheumatol. Int. 2022, 42, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Gold, J.E.; Okyay, R.A.; Licht, W.E.; Hurley, D.J. Investigation of Long COVID Prevalence and Its Relationship to Epstein–Barr Virus Reactivation. Pathogens 2021, 10, 763. [Google Scholar] [CrossRef] [PubMed]
- Walton, A.H.; Muenzer, J.T.; Rasche, D.; Boomer, J.S.; Sato, B.; Brownstein, B.H.; Pachot, A.; Brooks, T.L.; Deych, E.; Shannon, W.D.; et al. Reactivation of Multiple Viruses in Patients with Sepsis. PLoS ONE 2014, 9, e98819. [Google Scholar] [CrossRef]
- Libert, N.; Bigaillon, C.; Chargari, C.; Bensalah, M.; De Rudnicki, S.; Muller, V.; Mérat, S.; Nicolas, L.; Violaine, M.; Stéphane, M.; et al. Epstein–Barr virus reactivation in critically ill immunocompetent patients. Biomed. J. 2015, 38, 70–76. [Google Scholar] [CrossRef]
- L’Huillier, A.G.; Mardegan, C.; Cordey, S.; Luterbacher, F.; Papis, S.; Hugon, F.; Kaiser, L.; Gervaix, A.; Posfay-Barbe, K.; Galetto-Lacour, A. Enterovirus, parechovirus, adenovirus and herpes virus type 6 viraemia in fever without source. Arch. Dis. Child. 2020, 105, 180–186. [Google Scholar] [CrossRef]
- Krumina, A.; Chapenko, S.; Kenina, V.; Mihailova, M.; Logina, I.; Rasa, S.; Gintere, S.; Viksna, L.; Svirskis, S.; Murovska, M. The role of HHV-6 and HHV-7 infections in the development of fibromyalgia. J. NeuroVirol. 2019, 25, 194–207. [Google Scholar] [CrossRef]
- Secchiero, P.; Carrigan, D.R.; Asano, Y.; Benedetti, L.; Crowley, R.W.; Komaroff, A.L.; Gallo, R.C.; Lusso, P. Detection of human herpesvirus 6 in plasma of children with primary infection and immunosuppressed patients by polymerase chain reaction. J. Infect. Dis. 1995, 171, 273–280. [Google Scholar] [CrossRef]
- Chapenko, S.; Millers, A.; Nora, Z.; Logina, I.; Kukaine, R.; Murovska, M. Correlation between HHV-6 reactivation and multiple sclerosis disease activity. J. Med. Virol. 2003, 69, 111–117. [Google Scholar] [CrossRef]
- Soldan, S.S.; Berti, R.; Salem, N.; Secchiero, P.; Flamand, L.; Calabresi, P.A.; Brennan, M.B.; Maloni, H.W.; McFarland, H.F.; Lin, H.C.; et al. Association of human herpes virus 6 (HHV-6) with multiple sclerosis: Increased IgM response to HHV-6 early antigen and detection of serum HHV-6 DNA. Nat. Med. 1997, 3, 1394–1397. [Google Scholar] [CrossRef]
- Akhyani, N.; Berti, R.; Brennan, M.B.; Soldan, S.S.; Eaton, J.M.; McFarland, H.F.; Jacobson, S. Tissue distribution and variant characterization of human herpesvirus (HHV)-6: Increased prevalence of HHV-6A in patients with multiple sclerosis. J. Infect. Dis. 2000, 182, 1321–1325. [Google Scholar] [CrossRef]
- Berti, R.; Brennan, M.B.; Soldan, S.S.; Ohayon, J.M.; Casareto, L.; McFarland, H.F.; Jacobson, S. Increased detection of serum HHV-6 DNA sequences during multiple sclerosis (MS) exacerbations and correlation with parameters of MS disease progression. J. NeuroVirol. 2002, 8, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.C.; Wang, W.Y.; Chen, K.Y.; Wei, Y.H.; Liang, W.M.; Jan, J.S.; Jiang, R.S. Quantification of plasma Epstein–Barr virus DNA in patients with advanced nasopharyngeal carcinoma. N. Engl. J. Med. 2004, 350, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.C.A.; Woo, J.K.S.; King, A.; Zee, B.C.Y.; Lam, W.K.J.; Chan, S.L.; Chu, S.W.I.; Mak, C.; Tse, I.O.L.; Leung, S.Y.M.; et al. Analysis of Plasma Epstein–Barr Virus DNA to Screen for Nasopharyngeal Cancer. N. Engl. J. Med. 2017, 377, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.T.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. JAMA 2021, 326, 1614–1621. [Google Scholar] [CrossRef]
- Hill, J.A.; Mayer, B.T.; Xie, H.; Leisenring, W.M.; Huang, M.L.; Stevens-Ayers, T.; Milano, F.; Delaney, C.; Jerome, K.R.; Zerr, D.M.; et al. Kinetics of Double-Stranded DNA Viremia After Allogeneic Hematopoietic Cell Transplantation. Clin. Infect. Dis. 2018, 66, 368–375. [Google Scholar] [CrossRef]
- Black, S.; Kushner, I.; Samols, D. C-reactive Protein. J. Biol. Chem. 2004, 279, 48487–48490. [Google Scholar] [CrossRef]
- Sproston, N.R.; Ashworth, J.J. Role of C-Reactive Protein at Sites of Inflammation and Infection. Front. Immunol. 2018, 9, 754. [Google Scholar] [CrossRef]
- Weitz, J.I.; Fredenburgh, J.C.; Eikelboom, J.W. A Test in Context: D-Dimer. J. Am. Coll. Cardiol. 2017, 70, 2411–2420. [Google Scholar] [CrossRef]
- Johnson, E.D.; Schell, J.C.; Rodgers, G.M. The D-dimer assay. Am. J. Hematol. 2019, 94, 833–839. [Google Scholar] [CrossRef]
- Rostami, M.; Mansouritorghabeh, H. D-dimer level in COVID-19 infection: A systematic review. Expert Rev. Hematol. 2020, 13, 1265–1275. [Google Scholar] [CrossRef]
- Paliogiannis, P.; Mangoni, A.A.; Dettori, P.; Nasrallah, G.K.; Pintus, G.; Zinellu, A. D-Dimer Concentrations and COVID-19 Severity: A Systematic Review and Meta-Analysis. Front. Public Health 2020, 8, 432. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Chiang, H.H.; Cho, Y.T.; Chang, C.Y.; Chen, K.L.; Yang, C.W.; Lee, Y.H.; Chu, C.Y. Human herpes virus reactivations and dynamic cytokine profiles in patients with cutaneous adverse drug reactions—A prospective comparative study. Allergy 2015, 70, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Sousa, H.; Silva, J.; Azevedo, L.; Pinto-Correia, A.L.; Catarino, R.; Pinto, D.; Lopes, C.; Medeiros, R. Epstein–Barr virus in healthy individuals from Portugal. Acta Médica Port. 2011, 24, 707–712. [Google Scholar]
- Smatti, M.K.; Yassine, H.M.; AbuOdeh, R.; AlMarawani, A.; Taleb, S.A.; Althani, A.A.; Nasrallah, G.K. Prevalence and molecular profiling of Epstein Barr virus (EBV) among healthy blood donors from different nationalities in Qatar. PLoS ONE 2017, 12, e0189033. [Google Scholar] [CrossRef]
- Hudnall, S.D.; Chen, T.; Allison, P.; Tyring, S.K.; Heath, A. Herpesvirus prevalence and viral load in healthy blood donors by quantitative real-time polymerase chain reaction. Transfusion 2008, 48, 1180–1187. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Song, J.; Liu, H.; Zheng, H.; Chen, C. Positive Epstein–Barr virus detection in coronavirus disease 2019 (COVID-19) patients. Sci. Rep. 2021, 11, 10902. [Google Scholar] [CrossRef]
- Paolucci, S.; Cassaniti, I.; Novazzi, F.; Fiorina, L.; Piralla, A.; Comolli, G.; Bruno, R.; Maserati, R.; Gulminetti, R.; Novati, S.; et al. EBV DNA increase in COVID-19 patients with impaired lymphocyte subpopulation count. Int. J. Infect. Dis. 2021, 104, 315–319. [Google Scholar] [CrossRef]
- Meng, M.; Zhang, S.; Dong, X.; Sun, W.; Deng, Y.; Li, W.; Li, R.; Annane, D.; Wu, Z.; Chen, D. COVID-19 associated EBV reactivation and effects of ganciclovir treatment. Immun. Inflamm. Dis. 2022, 10, e597. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | n = 67, Median, (IQR) |
|---|---|
| Age at Admission | 60 (48, 66) |
| First Weight (kg) | 91 (76, 112) |
| Unknown | 1 |
| Height (m) | 1.68 (1.60, 1.83) |
| Unknown | 6 |
| Gender | |
| Female | 28 (42%) |
| Male | 39 (58%) |
| Race | |
| African American | 23 (34%) |
| Native American | 1 (1.5%) |
| Caucasian | 31 (46%) |
| Other | 12 (18%) |
| Smoking Status | |
| Current | 1 (2.1%) |
| Former | 14 (29%) |
| Never | 24 (50%) |
| Unknown | 9 (19%) |
| NA | 19 |
| Need for ICU | |
| No | 28 (42%) |
| Yes | 39 (58%) |
| Need for Intubation | |
| No | 41 (61%) |
| Yes | 26 (39%) |
| Blood Type | |
| A− | 2 (4.8%) |
| A+ | 14 (33%) |
| AB+ | 1 (2.4%) |
| B+ | 3 (7.1%) |
| O− | 1 (2.4%) |
| O+ | 21 (50%) |
| Not determined | 25 |
| Highest Temperature | |
| <38 | 42 (49%) |
| 101 | 12 (18%) |
| >38.5 | 12 (18%) |
| >40 | 1 (1.5%) |
| Length of Stay (Days) | 10 (5, 22) |
| Time to Death (Days) | |
| 6 | 1 |
| 10–20 | 2 |
| 20–30 | 3 |
| >30 | 2 |
| Highest D-Dimer | 605 (280, 2074) |
| Highest CRP | 10 (4, 16) |
| BMI | 33 (27, 38) |
| Need for Vasopressors | |
| Yes | 44 (66%) |
| Dexamethasone | |
| Yes | 41 (61%) |
| Prednisone | |
| Yes | 3 (13%) |
| Methylprednisolone | |
| Yes | 5 (22%) |
| Tocilizumab | |
| Yes | 4 (6%) |
| Remdesivir | |
| Yes | 27 (40%) |
| Creatinine | 1.00 (0.80, 1.45) |
| Hgb | 12.4 (10.8, 13.6) |
| IFNa3 | 45 (36, 72) |
| Not determined | 45 |
| Highest Ferritin | 1277 (7, 7161) |
| Not determined | 5 |
| Highest Troponin | |
| <0.04 | 47 (73%) |
| 0.04–0.4 | 12 (18%) |
| >0.6 | 3 (5%) |
| Not determined | 5 |
| Highest Creatine | 1.10 (0.90, 1.80) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brooks, B.; Tancredi, C.; Song, Y.; Mogus, A.T.; Huang, M.-L.W.; Zhu, H.; Phan, T.L.; Zhu, H.; Kadl, A.; Woodfolk, J.; et al. Epstein–Barr Virus and Human Herpesvirus-6 Reactivation in Acute COVID-19 Patients. Viruses 2022, 14, 1872. https://doi.org/10.3390/v14091872
Brooks B, Tancredi C, Song Y, Mogus AT, Huang M-LW, Zhu H, Phan TL, Zhu H, Kadl A, Woodfolk J, et al. Epstein–Barr Virus and Human Herpesvirus-6 Reactivation in Acute COVID-19 Patients. Viruses. 2022; 14(9):1872. https://doi.org/10.3390/v14091872
Chicago/Turabian StyleBrooks, Bailey, Christina Tancredi, Yufeng Song, Alemu Tekewe Mogus, Meei-Li W. Huang, Haiying Zhu, Tuan L. Phan, Harrison Zhu, Alexandra Kadl, Judith Woodfolk, and et al. 2022. "Epstein–Barr Virus and Human Herpesvirus-6 Reactivation in Acute COVID-19 Patients" Viruses 14, no. 9: 1872. https://doi.org/10.3390/v14091872