
Phytochemical Discrimination, Biological Activity and Molecular Docking of Water-Soluble Inhibitors from Saussurea costus Herb against Main Protease of SARS-CoV-2

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Plasmids
2.2. Extraction and Preparation for GC-MS Analysis
2.3. GC-MS Analysis Conditions
2.4. Molecular Docking
2.5. Antibacterial Activity Evaluation
2.6. Cytotoxicity Assays
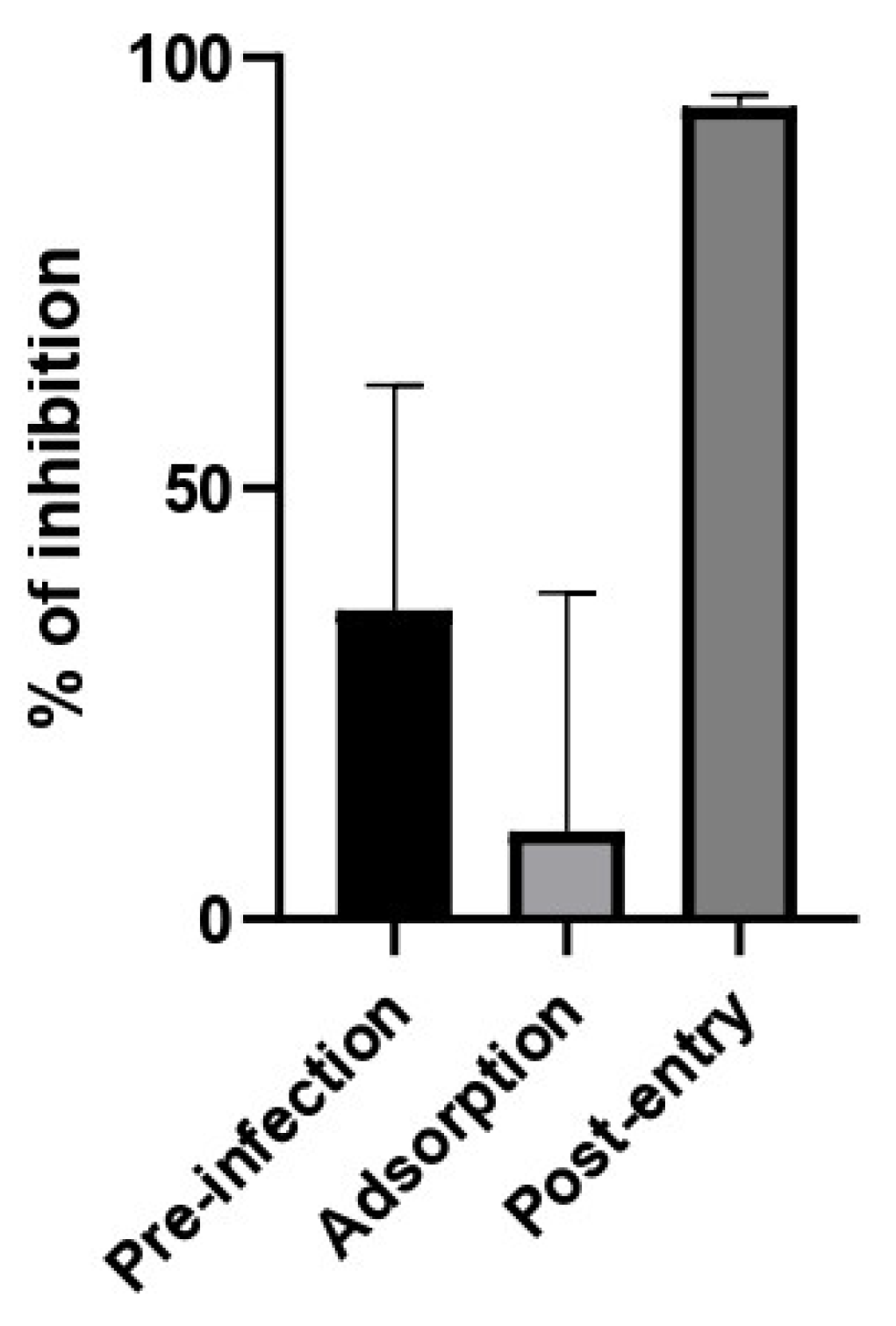
2.7. Antiviral Activity against SARS-CoV-2
2.8. Time of Addition Assay of HSV-1
2.9. Statistical Analysis
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Zimmermann, P.; Curtis, N. Coronavirus infections in children including COVID-19: An overview of the epidemiology, clinical features, diagnosis, treatment and prevention options in children. Pediatric Infect. Dis. J. 2020, 39, 355. [Google Scholar] [CrossRef] [PubMed]
- Nadda, R.K.; Ali, A.; Goyal, R.C.; Khosla, P.K.; Goyal, R. Aucklandia costus (syn. Saussurea costus): Ethnopharmacology of an endangered medicinal plant of the Himalayan region. J. Ethnopharmacol. 2020, 263, 113199. [Google Scholar] [CrossRef] [PubMed]
- Wald, E.R.; Schmit, K.M.; Gusland, D.Y. A pediatric infectious disease perspective on COVID-19. Clin. Infect. Dis. 2021, 72, 1660–1666. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Wang, Q.; Vlasova, A.N.; Jung, K.; Xiao, S. Coronaviruses. Dis. Swine 2019, 3, 488–523. [Google Scholar]
- McIntosh, K.; Perlman, S. Coronaviruses, including severe acute respiratory syndrome (SARS) and Middle East respiratory syndrome (MERS). Mandell Douglas Bennett’s Princ. Pract. Infect. Dis. 2015, 2, 1928–1936. [Google Scholar]
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent endemic coronavirus infection is associated with less-severe COVID-19. J. Clin. Investig. 2021, 131, 1–5. [Google Scholar] [CrossRef]
- Fani, M.; Teimoori, A.; Ghafari, S. Comparison of the COVID-19 (SARS-CoV-2) pathogenesis with SARS-CoV and MERS-CoV infections. Future Virol. 2020, 15, 317–323. [Google Scholar] [CrossRef]
- Holmes, E.C.; Goldstein, S.A.; Rasmussen, A.L.; Robertson, D.L.; Crits-Christoph, A.; Wertheim, J.O.; Anthony, S.J.; Barclay, W.S.; Boni, M.F.; Doherty, P.C. The origins of SARS-CoV-2: A critical review. Cell 2021, 184, 4848–4856. [Google Scholar] [CrossRef]
- Wang, H.; Li, X.; Li, T.; Zhang, S.; Wang, L.; Wu, X.; Liu, J. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1629–1635. [Google Scholar] [CrossRef]
- Hu, J.; Peng, P.; Cao, X.; Wu, K.; Chen, J.; Wang, K.; Tang, N.; Huang, A.-L. Increased immune escape of the new SARS-CoV-2 variant of concern Omicron. Cell. Mol. Immunol. 2022, 19, 293–295. [Google Scholar] [CrossRef]
- Glaß, M.; Misiak, D.; Misiak, C.; Müller, S.; Rausch, A.; Angermann, K.; Hoyer, M.; Zabel, R.; Kehlen, A.; Möbius, B. Fast forward evolution in real time: The rapid spread of SARS-CoV-2 variant of concern lineage B. 1.1. 7 in Saxony-Anhalt over a period of 5 months. Lab. Med. 2022, 46, 71–75. [Google Scholar] [CrossRef]
- Grabowski, F.; Kochańczyk, M.; Lipniacki, T. The spread of SARS-CoV-2 variant Omicron with a doubling time of 2.0–3.3 days can be explained by immune evasion. Viruses 2022, 14, 294. [Google Scholar] [CrossRef] [PubMed]
- Pollard, C.A.; Morran, M.P.; Nestor-Kalinoski, A.L. The COVID-19 pandemic: A global health crisis. Physiol. Genom. 2020, 52, 549–557. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Coronavirus Disease 2019 (Covid-19): Situation Report, 73; World Health Organization: New York, NY, USA, 2020. [Google Scholar]
- World Health Organization. Coronavirus Disease (Covid-19): Situation Report, 182; World Health Organization: New York, NY, USA, 2020. [Google Scholar]
- Kokudo, N.; Sugiyama, H. Call for international cooperation and collaboration to effectively tackle the COVID-19 pandemic. Glob. Health Med. 2020, 2, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, J.; Razavi Bazaz, S.; Aboulkheyr Es, H.; Yaghobian Azari, D.; Thierry, B.; Ebrahimi Warkiani, M.; Ghadiri, M. Lung-on-a-chip: The future of respiratory disease models and pharmacological studies. Crit. Rev. Biotechnol. 2020, 40, 213–230. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Sadeek, A.; Abdallah, E. Medicinal Plants with Antiviral Properties to Tackle Covid-19 Pandemic: A Short-Review. Antivirals 2021, 2, 122–127. [Google Scholar]
- Adhikari, B.; Marasini, B.P.; Rayamajhee, B.; Bhattarai, B.R.; Lamichhane, G.; Khadayat, K.; Adhikari, A.; Khanal, S.; Parajuli, N. Potential roles of medicinal plants for the treatment of viral diseases focusing on COVID-19: A review. Phytother. Res. 2021, 35, 1298–1312. [Google Scholar] [CrossRef]
- Abdallah, E.M. Controlling Covid-19 Needs More Than Just a Vaccine; It Requires An Integrated Control Strategy. Open Access J. Biol. Sci. 2021, 3, 1395–1396. [Google Scholar] [CrossRef]
- Hussain, M.; Khera, R.A.; Iqbal, J.; Khalid, M.; Hanif, M.A. Phytochemicals: Key to effective anticancer drugs. Mini-Rev. Org. Chem. 2019, 16, 141–158. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, J.; Ping, S.; Ma, Q.; Chen, X.; Xuan, H.; Shi, J.; Zhang, C.; Hu, F. Anti-inflammatory effects of ethanol extracts of Chinese propolis and buds from poplar (Populus × canadensis). J. Ethnopharmacol. 2014, 155, 300–311. [Google Scholar] [CrossRef]
- Ali, S.I.; Venkatesalu, V. Botany, traditional uses, phytochemistry and pharmacological properties of Saussurea costus—An endangered plant from Himalaya—A review. Phytochem. Lett. 2022, 47, 140–155. [Google Scholar] [CrossRef]
- Schippmann, U.; Leaman, D.J.; Cunningham, A. Impact of cultivation and gathering of medicinal plants on biodiversity: Global trends and issues. In FAO. 2002. Biodiversity and the Ecosystem Approach in Agriculture, Forestry and Fisheries. Satellite Event on the occasion of the Ninth Regular Session of the Commission on Genetic Resources for Food and Agriculture, Rome, Italy, 12–13 October 2002; Inter-Departmental Working Group on Biological Diversity for Food and Agriculture: Rome, Italy, 2002. [Google Scholar]
- Pandey, M.M.; Rastogi, S.; Rawat, A.K.S. Saussurea costus: Botanical, chemical and pharmacological review of an ayurvedic medicinal plant. J. Ethnopharmacol. 2007, 110, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Baik, K.U.; Jung, J.H.; Park, M.H. In vitro anti-inflammatory effects of cynaropicrin, a sesquiterpene lactone, from Saussurea lappa. Eur. J. Pharmacol. 2000, 398, 399–407. [Google Scholar] [CrossRef]
- Julianti, T.; Hata, Y.; Zimmermann, S.; Kaiser, M.; Hamburger, M.; Adams, M. Antitrypanosomal sesquiterpene lactones from Saussurea costus. Fitoterapia 2011, 82, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-K.; Park, S.-J.; Nam, S.-Y.; Kang, S.; Hwang, J.; Lee, S.-J.; Im, D.-S. Anti-allergic effects of sesquiterpene lactones from Saussurea costus (Falc.) Lipsch. determined using in vivo and in vitro experiments. J. Ethnopharmacol. 2018, 213, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Avdeeva, E.; Reshetov, Y.; Domrachev, D.; Gulina, E.; Krivoshchekov, S.; Shurupova, M.; Brazovskii, K.; Belousov, M. Constituent composition of the essential oils from some species of the genus Saussurea DC. Nat. Prod. Res. 2022, 36, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.F.; Shetaia, Y.M.; Tayel, A.A.; Munshi, A.M.; Alatawi, F.A.; Alsieni, M.A.; Al-Saman, M.A. Exploring the Antifungal Activity and Action of Saussurea costus Root Extracts against Candida albicans and Non-albicans Species. Antibiotics 2022, 11, 327. [Google Scholar] [CrossRef]
- Abdel-Wahhab, K.G.; Mannaa, F.A.; El-Sahra, D.G.; Morsy, F.A.; Gomaa, H.F. Effect of oral administration of methanolic root extract of Saussurea costus to rats after propylthiouracil-induced hypothyroid obesity. Comp. Clin. Pathol. 2022, 31, 377–390. [Google Scholar] [CrossRef]
- Ashry, M.; Galal ELSahra, D.; Abdel-Wahhab, K.G.; Abdelsalam, M.E.; Elmashad, W.; El-Bitar, A.M.; Gomaa, H.F. Saussurea Costus Extract Has Anti-İnflammatory, Antioxidant and Hormonal Effects Against Testicular Toxicity Induced by Oxaliplatin in Male Albino Rats. Iran. J. Toxicol. 2022, 16, 83–90. [Google Scholar] [CrossRef]
- Mohsen, E.; El-Far, A.H.; Godugu, K.; Elsayed, F.; Mousa, S.A.; Younis, I.Y. SPME and solvent-based GC–MS metabolite profiling of Egyptian marketed Saussurea costus (Falc.) Lipsch. concerning its anticancer activity. Phytomed. Plus 2022, 2, 100209. [Google Scholar] [CrossRef]
- Rumlová, M.; Ruml, T. In vitro methods for testing antiviral drugs. Biotechnol. Adv. 2018, 36, 557–576. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, E.M. Plants: An alternative source for antimicrobials. J. Appl. Pharm. Sci. 2011, 1, 16–20. [Google Scholar]
- Elzupir, A.O. Inhibition of SARS-CoV-2 main protease 3CLpro by means of α-ketoamide and pyridone-containing pharmaceuticals using in silico molecular docking. J. Mol. Struct. 2020, 1222, 128878. [Google Scholar] [CrossRef] [PubMed]
- Anju, A.; Chaturvedi, S.; Chaudhary, V.; Pant, P.; Hussain, F.; Mishra, A.K. Virtual screening of quinoline derived library for SARS-COV-2 targeting viral entry and replication. J. Biomol. Struct. Dyn. 2021, 1–30. [Google Scholar] [CrossRef]
- Hamed, M.I.; Darwish, K.M.; Soltane, R.; Chrouda, A.; Mostafa, A.; Shama, N.M.A.; Elhady, S.S.; Abulkhair, H.S.; Khodir, A.E.; Elmaaty, A.A. β-Blockers bearing hydroxyethylamine and hydroxyethylene as potential SARS-CoV-2 Mpro inhibitors: Rational based design, in silico, in vitro, and SAR studies for lead optimization. RSC Adv. 2021, 11, 35536–35558. [Google Scholar] [CrossRef]
- Rana, S.; Kumar, P.; Sharma, A.; Sharma, S.; Giri, R.; Ghosh, K.S. Identification of Naturally Occurring Antiviral Molecules for SARS-CoV-2 Mitigation. Open COVID J. 2021, 1, 38–46. [Google Scholar] [CrossRef]
- El-Shamy, N.T.; Alkaoud, A.M.; Hussein, R.K.; Ibrahim, M.A.; Alhamzani, A.G.; Abou-Krisha, M.M. DFT, ADMET and Molecular Docking Investigations for the Antimicrobial Activity of 6, 6′-Diamino-1, 1′, 3, 3′-Tetramethyl-5, 5′-(4-chlorobenzylidene) bis [pyrimidine-2, 4 (1H, 3H)-dione]. Molecules 2022, 27, 620. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, C.; Tieleman, D.P. Supramolecular Organization of SARS-CoV and SARS-CoV-2 Virions Revealed by Coarse-Grained Models of Intact Virus Envelopes. J. Chem. Inf. Modeling 2021, 62, 176–186. [Google Scholar] [CrossRef]
- Beltrán-Pavez, C.; Riquelme-Barrios, S.; Oyarzún-Arrau, A.; Gaete-Argel, A.; González-Stegmaier, R.; Cereceda-Solis, K.; Aguirre, A.; Travisany, D.; Palma-Vejares, R.; Barriga, G.P. Insights into neutralizing antibody responses in individuals exposed to SARS-CoV-2 in Chile. Sci. Adv. 2021, 7, eabe6855. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Elzupir, A.O. Caffeine and caffeine-containing pharmaceuticals as promising inhibitors for 3-chymotrypsin-like protease of SARS-CoV-2. J. Biomol. Struct. Dyn. 2022, 40, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- Al-Janabi, A.S.; Elzupir, A.O.; Yousef, T.A. Synthesis, anti-bacterial evaluation, DFT study and molecular docking as a potential 3-chymotrypsin-like protease (3CLpro) of SARS-CoV-2 inhibitors of a novel Schiff bases. J. Mol. Struct. 2021, 1228, 129454. [Google Scholar] [CrossRef] [PubMed]
- Elzupir, A.O. Molecular Docking and Dynamics Investigations for Identifying Potential Inhibitors of the 3-Chymotrypsin-like Protease of SARS-CoV-2: Repurposing of Approved Pyrimidonic Pharmaceuticals for COVID-19 Treatment. Molecules 2021, 26, 7458. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminformatics 2011, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shapovalov, M.V.; Dunbrack, R.L., Jr. A smoothed backbone-dependent rotamer library for proteins derived from adaptive kernel density estimates and regressions. Structure 2011, 19, 844–858. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Al-Mijalli, S.H.; Assaggaf, H.; Qasem, A.; El-Shemi, A.G.; Abdallah, E.M.; Mrabti, H.N.; Bouyahya, A. Antioxidant, Antidiabetic, and Antibacterial Potentials and Chemical Composition of Salvia officinalis and Mentha suaveolens Grown Wild in Morocco. Adv. Pharmacol. Pharm. Sci. 2022, 2022, 2844880. [Google Scholar] [CrossRef]
- Gabaglio, S.; Alvarenga, N.; Cantero-González, G.; Degen, R.; Ferro, E.; Langjahr, P.; Chnaiderman, J.; Sotelo, P. A quantitative PCR assay for antiviral activity screening of medicinal plants against Herpes simplex 1. Nat. Prod. Res. 2021, 35, 2926–2930. [Google Scholar] [CrossRef]
- González-Maldonado, P.; Alvarenga, N.; Burgos-Edwards, A.; Flores-Giubi, M.E.; Barúa, J.E.; Romero-Rodríguez, M.C.; Soto-Rifo, R.; Valiente-Echeverría, F.; Langjahr, P.; Cantero-González, G. Screening of Natural Products Inhibitors of SARS-CoV-2 Entry. Molecules 2022, 27, 1743. [Google Scholar] [CrossRef] [PubMed]
- Zia, M.; Muhammad, S.; Bibi, S.; Abbasi, S.W.; Al-Sehemi, A.G.; Chaudhary, A.R.; Bai, F.Q. Exploring the potential of novel phenolic compounds as potential therapeutic candidates against SARS-CoV-2, using quantum chemistry, molecular docking and dynamic studies. Bioorganic Med. Chem. Lett. 2021, 43, 128079. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.J.; Liang, P.H. Characterization and inhibition of the main protease of severe acute respiratory syndrome coronavirus. ChemBioEng Rev. 2015, 2, 118–132. [Google Scholar] [CrossRef]
- Muhseen, Z.T.; Hameed, A.R.; Al-Hasani, H.M.; ul Qamar, M.T.; Li, G. Promising terpenes as SARS-CoV-2 spike receptor-binding domain (RBD) attachment inhibitors to the human ACE2 receptor: Integrated computational approach. J. Mol. Liq. 2020, 320, 114493. [Google Scholar] [CrossRef] [PubMed]
- Giofrè, S.V.; Napoli, E.; Iraci, N.; Speciale, A.; Cimino, F.; Muscarà, C.; Molonia, M.S.; Ruberto, G.; Saija, A. Interaction of selected terpenoids with two SARS-CoV-2 key therapeutic targets: An in silico study through molecular docking and dynamics simulations. Comput. Biol. Med. 2021, 134, 104538. [Google Scholar] [CrossRef]
- Mei, J.; Zhou, Y.; Yang, X.; Zhang, F.; Liu, X.; Yu, B. Active components in Ephedra sinica stapf disrupt the interaction between ACE2 and SARS-CoV-2 RBD: Potent COVID-19 therapeutic agents. J. Ethnopharmacol. 2021, 278, 114303. [Google Scholar] [CrossRef]
- Hanh, T.T.H.; Hang, D.T.T.; Van Minh, C.; Dat, N.T. Anti-inflammatory effects of fatty acids isolated from Chromolaena odorata. Asian Pac. J. Trop. Med. 2011, 4, 760–763. [Google Scholar] [CrossRef] [Green Version]
- Auwal, M.S.; Saka, S.; Mairiga, I.A.; Sanda, K.A.; Shuaibu, A.; Ibrahim, A. Preliminary Phytochemical and Elemental Analysis of Aqueous and Fractionated Pod Extracts of Acacia nilotica (Thorn Mimosa). Vet. Res. Forum 2014, 5, 95–100. [Google Scholar]
- Villaseñor, I.M.; Cabrera, M.; Meneses, K.; Rivera, V.; Villasenor, R. Comparative antidiabetic activities of some medicinal plants. Philipp. J. Sci. 1998, 127, 261–266. [Google Scholar]
- Loza-Mejía, M.A.; Salazar, J.R. Sterols and triterpenoids as potential anti-inflammatories: Molecular docking studies for binding to some enzymes involved in inflammatory pathways. J. Mol. Graph. Model. 2015, 62, 18–25. [Google Scholar] [CrossRef]
- Kim, D.Y.; Choi, B.Y. Costunolide—A bioactive sesquiterpene lactone with diverse therapeutic potential. Int. J. Mol. Sci. 2019, 20, 2926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gwari, G.; Bhandari, U.; Andola, H.C.; Lohani, H.; Chauhan, N. Volatile constituents of Saussurea costus roots cultivated in Uttarakhand Himalayas, India. Pharmacogn. Res. 2013, 5, 179. [Google Scholar]
- Kim, E.J.; Lim, S.S.; Park, S.Y.; Shin, H.-K.; Kim, J.-S.; Park, J.H.Y. Apoptosis of DU145 human prostate cancer cells induced by dehydrocostus lactone isolated from the root of Saussurea lappa. Food Chem. Toxicol. 2008, 46, 3651–3658. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, E.M.; Qureshi, K.A.; Ali, A.M.; Elhassan, G.O. Evaluation of some biological properties of Saussurea costus crude root extract. Biosci. Biotechnol. Res. Commun. 2017, 10, 601–611. [Google Scholar] [CrossRef]
- Sagar, A.; Chauhan, V.; Prakash, V. Studies on endophytes and antibacterial activity of Saussurea costus (falc.). J. Drug Deliv. Ther. 2017, 7, 5–10. [Google Scholar] [CrossRef]
- Alaagib, R.M.O.; Ayoub, S.M.H. On the chemical composition and antibacterial activity of Saussurea lappa (Asteraceae). Pharma Innov. 2015, 4, 73. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
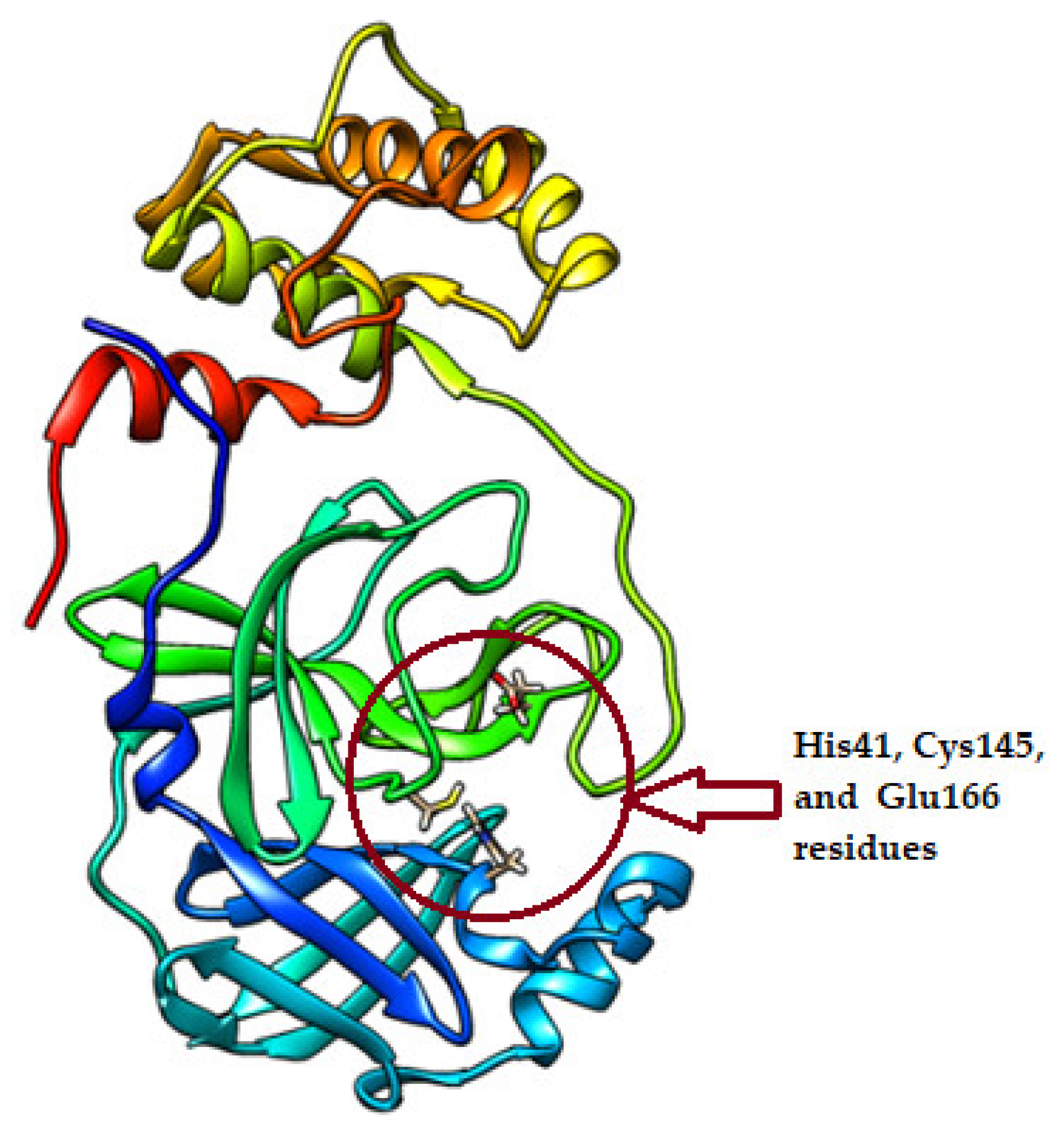
| Compound ID/Class | Structure | Binding Affinity (Kcal/Mol)/(RMSD) | Hydrogen Bond | van der Waals |
|---|---|---|---|---|
| Cmd1/carbohydrate |  | −4.7/ (0.00–3.2) | GLU 166, HIS 164, HIS 163 | HIS 164, MET 165, HIS 163, LEU 141, ASN 142, GLU 166, PHE 140, SER 144, CYS 145 |
| Cmd2/carbohydrate |  | −3.9/ (40.82–41.72) | HIS 163 | HIS 163, MET 165, GLU 166, SER 144, ASN 142, HIS 164, PHE 140 |
| Cmd3/carbohydrate |  | −4.3/ (40.10–41.20) | HIS 163 | GLU 166, HIS 163, MET 165, HIS 164, ASN 142, LEU 141, PHE 140, CYS 145 |
| Cmd5/carbohydrate |  | −4.4/ (27.24–29.57) | HIS 163, GLU 166 | HIS 163, MET 49, GLU 166, MET 165, HIS 41, GLN 189 |
| Cmd6/carboxylic acid |  | −3.8/ (25.23–26.15) | HIS 163, GLU 166, PHE 140 | GLU 166, MET 165, CYS 145, HIS 163, ASN 142, SER 144, PHE 140, LEU 141 |
| Cmd7/carbohydrate |  | −5.0/ (0.00–0.00) | HIS 163 | HIS 163, GLU 166, PHE 140, MET 165, HIS 164, CYS 145, ASN 142, SER 144, LEU 141 |
| Cmd8/fatty acid |  | −4.3/ (40.97–42.42) | HIS 163, GLU 166 | HIS 163, GLU 166, MET 165, HIS 164, PHE 140, MET 49, CYS 145, ASN 142, SER 144, LEU 141 |
| Cmd11/carbohydrate |  | −4.9/ (44.57–45.67) | - | HIS 163, GLU 166, MET 165, HIS 164, PHE 140, LEU 141, CYS 145, ASN 142, SER 144 |
| Cmd12/tannins |  | −4.6/ (44.33–45.19) | HIS 164 | HIS 163, HIS 164, MET 165, GLU 166, PHE 140, LEU 141, ASN 142, SER 144, CYS 145 |
| Cmd13/carbohydrate |  | −4.3/ (24.50–25.23) | PHE 140, HIS 163 | HIS 163, HIS 164, MET 165, GLU 166, PHE 140, LEU 141, ASN 142, SER 144, CYS 145, HIS 41 |
| Cmd15/fatty acid |  | −4.2/ (25.99–27.47) | HIS 163 | HIS 163, HIS 164, MET 165, GLU 166, PHE 140, LEU 141, GLY 143, ASN 142, SER 144, CYS 145 |
| Cmd18/Phenolic Compound |  | −4.9/ (42.17–45.62) | HIS 164, GLU 166 | GLN 189, PRO 168, THR 190, HIS 164, MET 165, GLU 166, MET 49, HIS 41, CYS 145 |
| Cmd20/carbohydrate |  | −5.1/ (29.29–33.07) | GLY 143 | MET 49, GLU 166, ASN 142, GLY 143, GLN 189, MET 165, LEU 141, HIS 41, CYS 145 |
| Cmd21/Carboxylic acid |  | −4.3/ (28.11–30.02) | GLU 166, GLY 143 | GLY 143, ASN 142, MET 49, THR 26, LEU 27, HIS 41, HIS 164, GLU 166, GLN 189, MET 165, CYS 145 |
| Cmd26/Fatty acid |  | −4.2/ (27.28–29.77) | HIS 163, SER 144 | SER 144, HIS 163, THR 25, LEU 27, GLU 166, GLN 189, ASN 142, MET 49, PHE 140, LEU 141 |
| Cmd30/Terpenoid |  | −5.2/ (29.65–31.24) | - | MET 49, THR 25, THR 26, LEU 27, HIS 41, ASN 142, CYS 145, HIS 164 |
| Cmd34/Hydrocarbon |  | −6.3/ (42.35–46.32) | GLU 166 | GLU 166, ASN 142, PRO 168, PHE 140, LEU 141, HIS 163, MET 165, ASN 142, CYS 145, LEU 167, HIS 164, GLN 189 |
| Cmd35/Terpenoid |  | −6.3/ (30.17–32.16) | GLU 166 | THR 25, MET 165, GLU 166, GLN 189, ASN 142, THR 45, SER 46, MET 49, CYS 44, HIS 41, CYS 145, THR 24 |
| Cmd38/Fatty acid |  | −3.7/ (26.63–30.65) | HIS 163 | HIS 163, GLU 166, MET 49, SER 144, GLN 189, PHE 140, LEU 141, ASN 142, HIS 41, HIS 164, MET 165 |
| Cmd40/Terpenoid |  | −5.4/ (43.11–45.74) | HIS 164 | GLU 166, MET 49, GLN 189, ASN 142, MET 165, CYS 145, HIS 164 |
| Cmd41/Fatty acid |  | −4.3/ (28.45–31.03) | HIS 163, PHE 140 | HIS 163, ASN 142, SER 144, LEU 141, PHE 140, LEU 27, GLN 189, MET 49, THR 25, THR 26, GLU 166, CYS 145, GLY 143, HIS 41, MET 165 |
| Cmd42/Sterol |  | −6.5/ (30.43–33.11) | - | MET 49, HIS 163, ASN 142, SER 144, LEU 141, PHE 140, LEU 27, GLN 189, THR 25, THR 26, GLU 166, CYS 145, GLY 143, HIS 41, MET 165 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Idriss, H.; Siddig, B.; Maldonado, P.G.; Elkhair, H.M.; Alakhras, A.I.; Abdallah, E.M.; Torres, P.H.S.; Elzupir, A.O. Phytochemical Discrimination, Biological Activity and Molecular Docking of Water-Soluble Inhibitors from Saussurea costus Herb against Main Protease of SARS-CoV-2. Molecules 2022, 27, 4908. https://doi.org/10.3390/molecules27154908
Idriss H, Siddig B, Maldonado PG, Elkhair HM, Alakhras AI, Abdallah EM, Torres PHS, Elzupir AO. Phytochemical Discrimination, Biological Activity and Molecular Docking of Water-Soluble Inhibitors from Saussurea costus Herb against Main Protease of SARS-CoV-2. Molecules. 2022; 27(15):4908. https://doi.org/10.3390/molecules27154908
Chicago/Turabian StyleIdriss, Hajo, Babeker Siddig, Pamela González Maldonado, H. M. Elkhair, A. I. Alakhras, Emad M. Abdallah, Pablo Hernán Sotelo Torres, and Amin O. Elzupir. 2022. "Phytochemical Discrimination, Biological Activity and Molecular Docking of Water-Soluble Inhibitors from Saussurea costus Herb against Main Protease of SARS-CoV-2" Molecules 27, no. 15: 4908. https://doi.org/10.3390/molecules27154908