
Vitamin D Status and Immune Response in Hospitalized Patients with Moderate and Severe COVID-19

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Results
2.1. Comparison According to Disease Severity
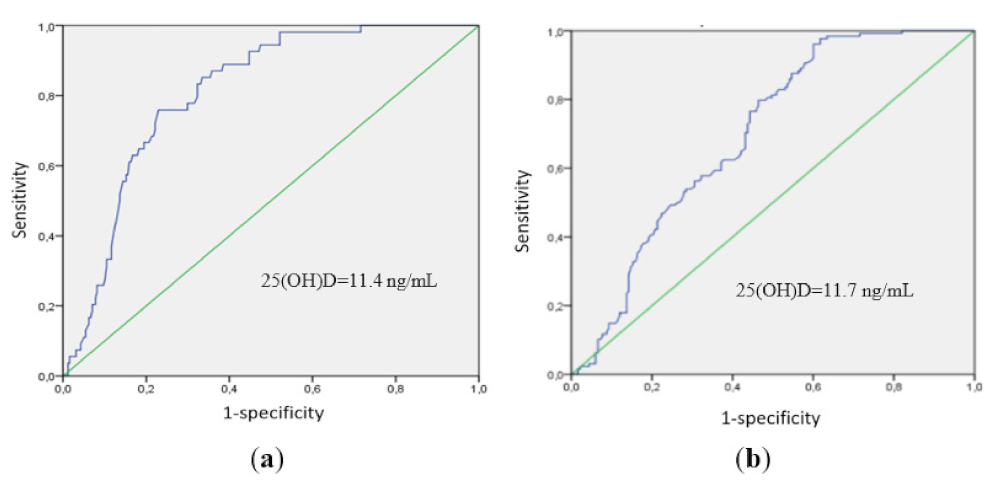
2.2. Comparison According to Vitamin D Status
2.3. Immune Status in Patients with COVID-19 and Vitamin D Deficiency and Insufficiency
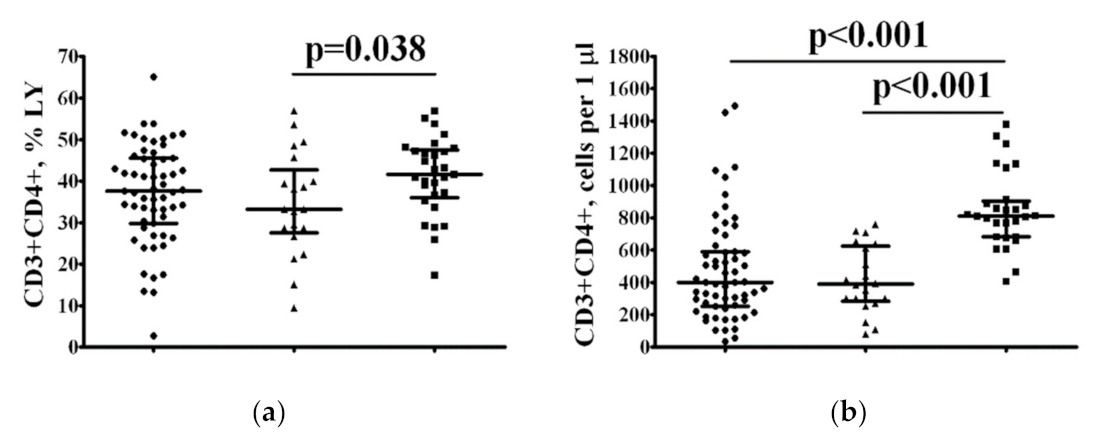
2.3.1. Decreased Frequency of Th Cells According to Vitamin D Status
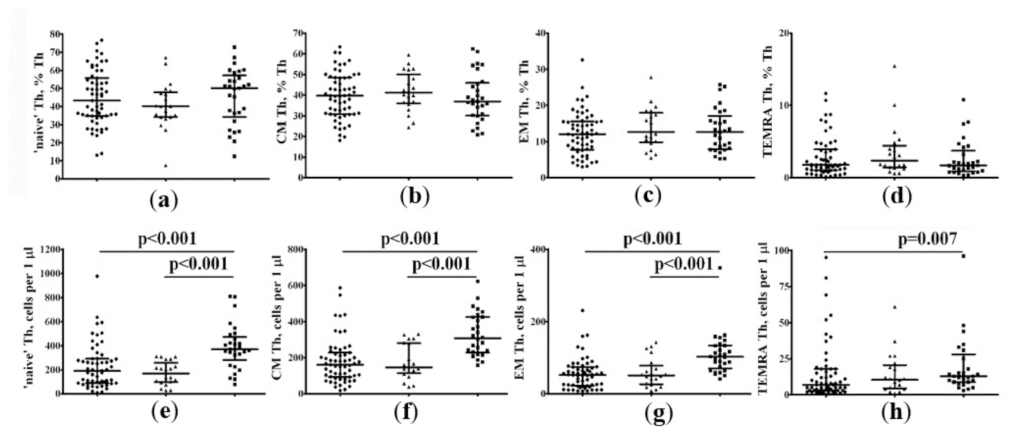
2.3.2. Alterations in the Main T-Cell Subsets in Patients with COVID-19 According to Vitamin D Status
2.3.3. Imbalance of the Central Memory “Polarized” Th Cell Subsets According to Vitamin D Status
2.3.4. Alterations in the Tfh Subsets in Patients with COVID-19 According to Vitamin D Status
2.3.5. Imbalance of Effector Memory “Polarized” Th Cell Subsets According to Vitamin D Status
3. Discussion
4. Materials and Methods
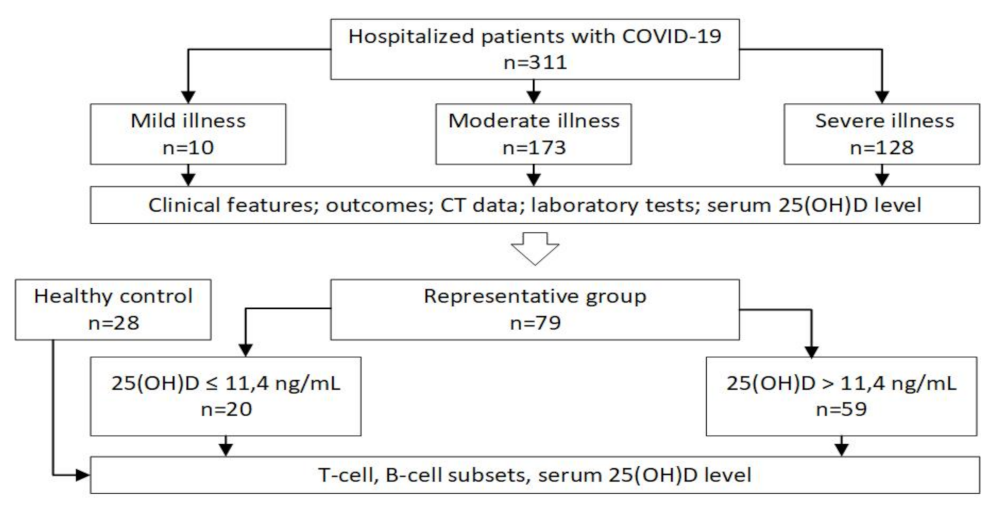
4.1. Patients
4.2. Physical Data
4.3. Laboratory Tests
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khalifa, S.A.M.; Swilam, M.M.; El-Wahed, A.A.A.; Du, M.; El-Seedi, H.H.R.; Kai, G.; Masry, S.H.D.; Abdel-Daim, M.M.; Zou, X.; Halabi, M.F.; et al. Beyond the Pandemic: COVID-19 Pandemic Changed the Face of Life. Int. J. Environ. Res. Public Health 2021, 18, 5645. [Google Scholar] [CrossRef] [PubMed]
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19). In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Selvaraj, P.; Harishankar, M.; Afsal, K. Vitamin D: Immuno-modulation and tuberculosis treatment. Can. J. Physiol. Pharmacol. 2015, 93, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Meinken, C.; Kamen, D.L.; Wagner, M.; Bals, R.; Steinmeyer, A.; Zügel, U.; Gallo, R.L.; Eisenberg, D.; Hewison, M.; Hollis, B.W.; et al. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef]
- Wang, T.T.; Nestel, F.P.; Bourdeau, V.; Nagai, Y.; Wang, Q.; Liao, J.; Tavera-Mendoza, L.; Lin, R.; Hanrahan, J.W.; Mader, S.; et al. Cutting edge: 1,25-dihydroxyvitamin D3 is a direct inducer of antimicrobial peptide gene expression. J. Immunol. 2004, 173, 2909–2912. [Google Scholar] [CrossRef] [Green Version]
- Urashima, M.; Segawa, T.; Okazaki, M.; Kurihara, M.; Wada, Y.; Ida, H. Randomized trial of vitamin D supplementation to prevent seasonal influenza A in schoolchildren. Am. J. Clin. Nutr. 2010, 91, 1255–1260. [Google Scholar] [CrossRef] [Green Version]
- Mercola, J.; Grant, W.B.; Wagner, C.L. Evidence Regarding Vitamin D and Risk of COVID-19 and Its Severity. Nutrients 2020, 12, 3361. [Google Scholar] [CrossRef]
- Panagiotou, G.; Tee, S.A.; Ihsan, Y.; Athar, W.; Marchitelli, G.; Kelly, D.; Boot, C.S.; Stock, N.; Macfarlane, J.; Martineau, A.R.; et al. Low serum 25-hydroxyvitamin D (25[OH]D) levels in patients hospitalized with COVID-19 are associated with greater disease severity. Clin. Endocrinol. 2020, 93, 508–511. [Google Scholar] [CrossRef]
- Carpagnano, G.E.; Di Lecce, V.; Quaranta, V.N.; Zito, A.; Buonamico, E.; Capozza, E.; Palumbo, A.; Di Gioia, G.; Valerio, V.N.; Resta, O. Vitamin D deficiency as a predictor of poor prognosis in patients with acute respiratory failure due to COVID-19. J. Endocrinol. Investig. 2021, 44, 765–771. [Google Scholar] [CrossRef]
- Pizzini, A.; Aichner, M.; Sahanic, S.; Bohm, A.; Egger, A.; Hoermann, G.; Kurz, K.; Widmann, G.; Bellmann-Weiler, R.; Weiss, G. Impact of Vitamin D Deficiency on COVID-19—A Prospective Analysis from the CovILD Registry. Nutrients 2020, 12, 2775. [Google Scholar] [CrossRef]
- Macaya, F.; Espejo Paeres, C.; Valls, A.; Fernandez-Ortiz, A.; Gonzalez Del Castillo, J.; Martin-Sanchez, J.; Runkle, I.; Rubio Herrera, M.A. Interaction between age and vitamin D deficiency in severe COVID-19 infection. Nutr. Hosp. 2020, 37, 1039–1042. [Google Scholar] [CrossRef]
- Karonova, T.L.; Andreeva, A.T.; Golovatuk, K.A.; Bykova, E.S.; Simanenkova, A.V.; Vashukova, M.A.; Grant, W.B.; Shlyakhto, E.V. Low 25(OH)D Level is Associated with Severe Course and Poor Prognosis in COVID-19. Nutrients 2021, 13, 3021. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, H.A.; de Silva, N.L.; Sumanatilleke, M.; de Silva, S.D.N.; Gamage, K.K.K.; Dematapitiya, C.; Kuruppu, D.C.; Ranasinghe, P.; Pathmanathan, S.; Katulanda, P. Prognostic and therapeutic role of vitamin D in COVID-19: Systematic review and meta-analysis. J. Clin. Endocrinol. Metab. 2021, 2021, dgab892. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Heaney, R.P. Functional indices of vitamin D status and ramifications of vitamin D deficiency. Am. J. Clin. Nutr. 2004, 80 (Suppl. S6), 1706S–1709S. [Google Scholar] [CrossRef]
- Baeke, F.; Korf, H.; Overbergh, L.; van Etten, E.; Verstuyf, A.; Gysemans, C.; Mathieu, C. Human T lymphocytes are direct targets of 1,25-dihydroxyvitamin D3 in the immune system. J. Steroid Biochem. Mol. Biol. 2010, 121, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Hewison, M.; Freeman, L.; Hughes, S.V.; Evans, K.N.; Bland, R.; Eliopoulos, A.G.; Kilby, M.D.; Moss, P.A.; Chakraverty, R. Differential regulation of vitamin D receptor and its ligand in human monocyte-derived dendritic cells. J. Immunol. 2003, 170, 5382–5390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, S.M.; Findlay, E.G.; McHugh, B.J.; Mackellar, A.; Man, T.; Macmillan, D.; Wang, H.; Fitch, P.M.; Schwarze, J.; Davidson, D.J. The human cathelicidin LL-37 has antiviral activity against respiratory syncytial virus. PLoS ONE 2013, 8, e73659. [Google Scholar] [CrossRef]
- Kota, S.; Sabbah, A.; Chang, T.H.; Harnack, R.; Xiang, Y.; Meng, X.; Bose, S. Role of human beta-defensin-2 during tumor necrosis factor-alpha/NF-kappaB-mediated innate antiviral response against human respiratory syncytial virus. J. Biol. Chem. 2008, 283, 22417–22429. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Sun, J. Vitamin D, vitamin D receptor, and macroautophagy in inflammation and infection. Discov. Med. 2011, 11, 325–335. [Google Scholar] [CrossRef]
- Cutolo, M.; Paolino, S.; Smith, V. Evidences for a protective role of vitamin D in COVID-19. RMD Open 2020, 6, e001454. [Google Scholar] [CrossRef]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.; O’Garra, A. 1alpha,25-Dihydroxyvitamin d3 has a direct effect on I CD4(+) T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranow, C. Vitamin D and the immune system. J. Investig. Med. 2011, 59, 881–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, W.B.; Al Anouti, F.; Boucher, B.J.; Dursun, E.; Gezen-Ak, D.; Jude, E.B.; Karonova, T.L.; Pludowski, P. A Narrative Review of the Evidence for Variations in Serum 25-Hydroxyvitamin D Concentration Thresholds for Optimal Health. Nutrients 2022, 14, 639. [Google Scholar] [CrossRef]
- Morita, R.; Schmitt, N.; Bentebibel, S.E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human blood CXCR5(+) CD4(+) T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Chiodini, I.; Gatti, D.; Soranna, D.; Merlotti, D.; Mingiano, C.; Fassio, A.; Adami, G.; Falchetti, A.; Eller-Vainicher, C.; Rossini, M.; et al. Vitamin D Status and SARS-CoV-2 Infection and COVID-19 Clinical Outcomes. Front. Public Health 2021, 9, 736665. [Google Scholar] [CrossRef]
- AlSafar, H.; Grant, W.B.; Hijazi, R.; Uddin, M.; Alkaabi, N.; Tay, G.; Mahboub, B.; Al Anouti, F. COVID-19 Disease Severity and Death in Relation to Vitamin D Status among SARS-CoV-2-Positive UAE Residents. Nutrients 2021, 13, 1714. [Google Scholar] [CrossRef]
- Golovkin, A.; Kalinina, O.; Bezrukikh, V.; Aquino, A.; Zaikova, E.; Karonova, T. Imbalanced Immune Response of T cell and B cell Subsets in Patients with Moderate and Severe COVID-19. Viruses 2021, 13, 1966. Available online: https://www.mdpi.com/1999-4915/13/10/1966 (accessed on 29 January 2022). [CrossRef]
- Kudryavtsev, I.; Kalinina, O.; Bezrukikh, V.; Melnik, O.; Golovkin, A. The significance of phenotyping and quantification of plasma extracellular vesicles levels using high-sensitivity flow cytometry during COVID-19 treatment. Viruses 2021, 13, 767. Available online: https://www.mdpi.com/1999-4915/13/5/767 (accessed on 29 January 2022). [CrossRef]
- Malkova, A.; Kudlay, D.; Kudryavtsev, I.; Starshinova, A.; Yablonskiy, P.; Shoenfeld, Y. Immunogenetic Predictors of Severe COVID-19. Vaccines 2021, 9, 211. [Google Scholar] [CrossRef]
- Gallelli, L.; Mannino, G.C.; Luciani, F.; de Sire, A.; Mancuso, E.; Gangemi, P.; Cosco, L.; Monea, G.; Averta, C.; Minchella, P.; et al. Vitamin D Serum Levels in Subjects Tested for SARS-CoV-2: What Are the Differences among Acute, Healed, and Negative COVID-19 Patients? A Multicenter Real-Practice Study. Nutrients 2021, 13, 3932. [Google Scholar] [CrossRef]
- Dupuis, M.L.; Pagano, M.T.; Pierdominici, M.; Ortona, E. The role of vitamin D in autoimmune diseases: Could sex make the difference? Biol. Sex Differ. 2021, 12, 12. [Google Scholar] [CrossRef]
- Baeke, F.; Takiishi, T.; Korf, H.; Gysemans, C.; Mathieu, C. Vitamin D: Modulator of the immune system. Curr. Opin. Pharmacol. 2010, 10, 482–496. [Google Scholar] [CrossRef]
- Chun, R.F.; Liu, P.T.; Modlin, R.L.; Adams, J.S.; Hewison, M. Impact of vitamin D on immune function: Lessons learned from genome-wide analysis. Front. Physiol. 2014, 5, 151. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Pantalena, L.C.; Liu, X.K.; Gaffen, S.L.; Liu, H.; Rohowsky-Kochan, C.; Ichiyama, K.; Yoshimura, A.; Steinman, L.; Christakos, S.; et al. 1,25-dihydroxyvitamin D(3) ameliorates Th17 autoimmunity via transcriptional modulation of interleukin-17A. Mol. Cell. Biol. 2011, 31, 3653–3669. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Zhou, R.; Luger, D.; Zhu, W.; Silver, P.B.; Grajewski, R.S.; Su, S.B.; Chan, C.C.; Adorini, L.; Caspi, R.R. Calcitriol suppresses antiretinal autoimmunity through inhibitory effects on the Th17 effector response. J. Immunol. 2009, 182, 4624–4632. [Google Scholar] [CrossRef]
- Todosenko, N.; Vulf, M.; Yurova, K.; Khaziakhmatova, O.; Mikhailova, L.; Litvinova, L. Causal Links between Hypovitaminosis D and Dysregulation of the T Cell Connection of Immunity Associated with Obesity and Concomitant Pathologies. Biomedicines 2021, 9, 1750. [Google Scholar] [CrossRef]
- Urry, Z.; Chambers, E.S.; Xystrakis, E.; Dimeloe, S.; Richards, D.F.; Gabryšová, L.; Christensen, J.; Gupta, A.; Saglani, S.; Bush’, A.; et al. The role of 1α,25-dihydroxyvitamin D3 and cytokines in the promotion of distinct Foxp3+ and IL-10+ CD4+ T cells. Eur. J. Immunol. 2012, 42, 2697–2708. [Google Scholar] [CrossRef] [Green Version]
- Sigmundsdottir, H.; Pan, J.; Debes, G.F.; Alt, C.; Habtezion, A.; Soler, D.; Butcher, E.C. DCs metabolize sunlight-induced vitamin D to ‘program’ T cell attraction to the epidermal chemokine CCL27. Nat. Immunol. 2007, 8, 285–293. [Google Scholar] [CrossRef]
- Dankers, W.; Davelaar, N.; van Hamburg, J.P.; van de Peppel, J.; Colin, E.M.; Lubberts, E. Human Memory Th17 Cell Populations Change into Anti-inflammatory Cells with Regulatory Capacity Upon Exposure to Active Vitamin D. Front. Immunol. 2019, 10, 1504. [Google Scholar] [CrossRef] [Green Version]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.T.; Linster, M.; Tan, C.W.; Le Bert, N.; Chia, W.N.; Kunasegaran, K.; Zhuang, Y.; Tham, C.Y.L.; Chia, A.; Smith, G.J.D.; et al. Early induction of functional SARS-CoV-2-specific T cells associates with rapid viral clearance and mild disease in COVID-19 patients. Cell Rep. 2021, 34, 108728. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Prevention, Diagnosis and Treatment of New Coronavirus Infection (COVID-19). Temporary Guidelines. Version 9 from 10/26/2020. Available online: http://www.consultant.ru/law/hotdocs/65400.html/ (accessed on 29 January 2022). (In Russia).
- Holick, M.F.; Binkley, N.C.; Bischoff-Ferrari, H.A.; Gordon, C.M.; Hanley, D.A.; Heaney, R.P.; Murad, M.H.; Weaver, C.M.; Endocrine Society. Evaluation, treatment, and prevention of vitamin D deficiency: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2011, 96, 1911–1930. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Lenig, D.; Förster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Barbarash, L.; Kudryavtsev, I.; Rutkovskaya, N.; Golovkin, A.T. Cell Response in Patients with Implanted Biological and Mechanical Prosthetic Heart Valves. Mediat. Inflamm. 2016, 2016, 1937564. [Google Scholar] [CrossRef]
- Kudryavtsev, I.; Serebriakova, M.; Zhiduleva, E.; Murtazalieva, P.; Titov, V.; Malashicheva, A.; Shishkova, A.; Semenova, D.; Irtyuga, O.; Isakov, D.; et al. CD73 Rather Than CD39 Is Mainly Involved in Controlling Purinergic Signaling in Calcified Aortic Valve Disease. Front. Genet. 2019, 10, 604. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Moderate (n = 173) | Severe (n = 128) | p |
|---|---|---|---|
| Age, years, Me + IQR [25; 75] | 59 [51; 68] | 66 [58; 75] | <0.001 |
| Gender, female, n (%) | 89 (51.4) | 61 (47.7) | 0.52 |
| Days from symptom onset to hospitalization, days, Me + IQR [25; 75] | 8 [6; 11] | 8 [6; 11] | 0.92 |
| Bed days, days, Me + IQR [25; 75] | 15 [13; 20] | 19 [13; 27] | 0.003 |
| CT lung involvement, %, Me + IQR [25; 75] | 30 [20; 45] | 68 [55; 76] | <0.001 |
| BMI, kg/m2, Me + IQR [25; 75] | 29 [26; 32] | 29 [25; 34] | 0.66 |
| Obesity, n (%) | 69 (39.9) | 55 (43.0) | 0.048 |
| DM type 2, n (%) | 47 (27.2) | 55 (43.0) | 0.004 |
| AH, n (%) | 123 (71.1) | 111 (86.7) | 0.001 |
| IHD, n (%) | 37 (21.4) | 66 (51.6) | <0.001 |
| CKD, n (%) | 13 (7.5) | 21 (16.4) | 0.017 |
| Neutrophils, ×109/L, Me + IQR [25; 75] | 4.8 [3.4; 7.3] | 5.7 [3.4; 8.1] | 0.16 |
| Lymphocytes, ×109/L, Me + IQR [25; 75] | 1.2 [0.8; 1.5] | 0.9 [0.7; 1.3] | 0.002 |
| NLR, Me + IQR [25; 75] | 4.2 [2.7; 7.1] | 5.7 [2.9; 10.1] | 0.007 |
| 25(OH)D, ng/mL, Me + IQR [25; 75] | 18.9 [10.2; 33.3] | 14.8 [8.3; 21.3] | 0.001 |
| Plasma glucose, mmol/L, Me + IQR [25; 75] | 6.5 [5.9; 8.0] | 7.7 [6.5; 9.3] | <0.001 |
| CRP, mg/L, Me + IQR [25; 75] | 39.6 [20.7; 91.4] | 93.1 [53.4; 190.7] | <0.001 |
| Ferritin, ng/mL, Me + IQR [25; 75] | 488 [234; 825] | 776 [381; 1296] | <0.001 |
| LDH, u/L, Me + IQR [25; 75] | 341 [267; 482] | 541 [399; 726] | <0.001 |
| D-dimer, µg/mL FEU, Me + IQR [25; 75] | 0.3 [0.2; 0.5] | 0.6 [0.4; 0.9] | <0.001 |
| Creatinine, mcmol/L, Me + IQR [25; 75] | 68 [58; 77] | 68 [51; 83] | 0.74 |
| Parameters | Vitamin D Status | p | |
|---|---|---|---|
| Normal (n = 69) | Deficiency/Insufficiency (n = 242) | ||
| Age, years, Me + IQR [25; 75] | 58 [48; 69] | 64 [55; 72] | 0.002 |
| Gender, female, n (%) | 37 (53.6) | 120 (49.6) | 0.550 |
| Disease severity, n (%) | |||
| Mild Moderate Severe | 7 (10.2) 56 (81.1) 6 (8.7) | 3 (1.3) 117 (48.3) 122 (50.4) | < 0.001 |
| Vitamin D status, n (%) | |||
| Normal Insufficiency Deficiency Severe deficiency | 69 (100) - - - | - 58 (23.9) 184 (76) 102 (42.1) | |
| Days from symptom onset to hospitalization, days, Me + IQR [25; 75] | 7 [4; 10] | 8 [6; 11] | 0.050 |
| Bed days, days, Me + IQR [25; 75] | 14 [9; 19] | 17 [13; 23] | 0.002 |
| ICU, n (%) | 9 (13) | 97 (40,1) | 0.001 |
| Discharge, n (%) | 66 (95.6) | 191 (78.9) | 0.001 |
| CT lung involvement, %, Me + IQR [25; 75] | 15 [5; 25] | 48 [35; 70] | <0.001 |
| CT grading, n (%) | |||
| 1 2 3 | 39 (56.5) 16 (23.2) 1 (1.4) | 24 (9.9) 107 (44.2) 64 (26.4) | <0.001 |
| BMI, kg/m2, Me + IQR [25; 75] | 28 [25; 32] | 30 [26; 33] | 0.13 |
| Obesity, n (%) | 22 (31.8) | 103 (42.6) | 0.27 |
| DM type 2, n (%) | 22 (31.8) | 82 (33.8) | 0.70 |
| AH, n (%) | 48 (69.6) | 192 (79.3) | 0.08 |
| IHD, n (%) | 19 (27.5) | 87 (35.9) | 0.19 |
| CKD, n (%) | 5 (7.2) | 29 (11.9) | 0.26 |
| 25(OH)D, ng/mL, Me + IQR [25; 75] | 36.1 [32.7; 38.0] | 13.7 [8.2; 19.3] | <0.001 |
| Neutrophils, ×109/L, Me + IQR [25; 75] | 4.8 [3.1; 6.5] | 5.3 [3.4; 7.8] | 0.12 |
| Lymphocytes, ×109/L, Me + IQR [25; 75] | 1.2 [0.8; 1.5] | 1.0 [0.8; 1.4] | 0.08 |
| NLR, Me + IQR [25; 75] | 4.1 [2.4; 6.8] | 5.2 [2.8; 9.1] | 0.03 |
| Plasma glucose, mmol/L, Me + IQR [25; 75] | 6.6 [5.7; 8.2] | 7.0 [6.1; 8.6] | 0.04 |
| CRP, mg/L, Me + IQR [25; 75] | 28.3 [7.6; 62.5] | 79.7 [32.5; 146.7] | <0.001 |
| Ferritin, ng/mL, Me + IQR [25; 75] | 411 [183; 786] | 627 [289; 1155] | 0.002 |
| LDH, u/L, Me + IQR [25; 75] | 331 [239; 455] | 449 [316; 609] | <0.001 |
| D-dimer, µg/mL FEU, Me + IQR [25; 75] | 0.3 [0.2; 0.4] | 0.4 [0.3; 0.7] | 0.002 |
| Creatinine, µmol/L, Me + IQR [25; 75] | 71 [60.5; 82.5] | 65 [55; 79] | 0.043 |
| Parameters | Serum 25(OH)D Level, ng/mL | p | ||
|---|---|---|---|---|
| ≥30.0 (n = 69) | 20.0–29.9 (n = 57) | <20 (n = 185) | ||
| Age, years, Me + IQR (25; 75) | 58 [48; 69] | 65 [55; 71] | 64 [56; 72] | 0.003 |
| Gender, female, n (%) | 37 (53.6) | 29 (50.8) | 91 (49.1) | 0.38 |
| Disease severity, n (%) | ||||
| Mild Moderate Severe | 7 (10.2) 56 (81.1) 6 (8.7) | 3 (1.3) 28 (49.1) 29 (50.8) | - 89 (48.1) 93 (50.2) | <0.001 |
| Days from symptom onset to hospitalization, days, Me + IQR [25; 75] | 7 [4; 10] | 9 [6; 23] | 8 [7; 11] | 0.006 |
| Bed days, days, Me + IQR [25; 75] | 14 [9; 19] | 16 [12; 22] | 17 [13; 23] | 0.002 |
| ICU, n (%) | 9 (13) | 26 (45.6) | 71 (38.5) | 0.01 |
| Discharge, n (%) | 66 (95.6) | 45 (78.9) | 146 (78.9) | 0.02 |
| CT lung involvement, %, Me + IQR [25; 75] | 15 [5; 25] | 46.5 [35; 65] | 50 [35; 70] | <0.001 |
| BMI, kg/m2, Me + IQR [25; 75] | 28 [25; 32] | 28 [26; 32] | 30 [26; 33] | 0.26 |
| Obesity, n (%) | 22 (31.8) | 21 (36.8) | 82 (44.3) | 0.18 |
| DM type 2, n (%) | 22 (31.8) | 16 (28) | 66 (35.6) | 0.32 |
| AH, n (%) | 48 (69.6) | 44 (77.2) | 148 (80) | 0.15 |
| IHD, n (%) | 19 (27.5) | 16 (28) | 71 (38.4) | 0.09 |
| CKD, n (%) | 5 (7.2) | 6 (10.5) | 23 (12.4) | 0.22 |
| 25(OH)D, ng/mL, Me + IQR [25; 75] | 36.1 [32.7; 38.0] | 25.4 [22.1; 26.6] | 11.2 [7.2; 14.9] | <0.001 |
| Neutrophils, ×109/L, Me + IQR [25; 75] | 4.8 [3.1; 6.5] | 5.7 [3.1; 7.5] | 5.2 [3.4; 7.8] | 0.46 |
| Lymphocytes, ×109/L, Me + IQR [25; 75] | 1.2 [0.8; 1.5] | 1.1 [0.8; 1.5] | 1.0 [0.7; 1.39] | 0.02 |
| NLR, Me + IQR [25; 75] | 4.1 [2.4; 6.8] | 4.8 [2.3; 9.1] | 5.2 [2.7; 8.7] | 0.006 |
| Plasma glucose, mmol/L, Me + IQR [25; 75] | 6.6 [5.7; 8.2] | 6.7 [6.0; 8.2] | 7.1 [6.2; 8.8] | 0.038 |
| CRP, mg/L, Me + IQR [25; 75] | 28.3 [7.6; 62.5] | 86.8 [31.8; 183.6] | 77.9 [33.3; 132.6] | <0.001 |
| Ferritin, ng/mL, Me + IQR [25; 75] | 411 [183; 786] | 594 [290; 836] | 662 [289; 1193] | 0.003 |
| LDH, u/L, Me + IQR [25; 75] | 331 [239; 455] | 417 [318; 575] | 430 [314; 628] | <0.001 |
| D-dimer, µg/mL FEU, Me + IQR [25; 75] | 0.3 [0.2; 0.4] | 0.5 [0.3; 0.8] | 0.4 [0.5; 0.7] | 0.001 |
| Creatinine, µmol/L, Me + IQR [25; 75] | 71 [60.5; 82.5] | 65 [55; 84] | 67 [55; 79] | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karonova, T.L.; Kudryavtsev, I.V.; Golovatyuk, K.A.; Aquino, A.D.; Kalinina, O.V.; Chernikova, A.T.; Zaikova, E.K.; Lebedev, D.A.; Bykova, E.S.; Golovkin, A.S.; et al. Vitamin D Status and Immune Response in Hospitalized Patients with Moderate and Severe COVID-19. Pharmaceuticals 2022, 15, 305. https://doi.org/10.3390/ph15030305
Karonova TL, Kudryavtsev IV, Golovatyuk KA, Aquino AD, Kalinina OV, Chernikova AT, Zaikova EK, Lebedev DA, Bykova ES, Golovkin AS, et al. Vitamin D Status and Immune Response in Hospitalized Patients with Moderate and Severe COVID-19. Pharmaceuticals. 2022; 15(3):305. https://doi.org/10.3390/ph15030305
Chicago/Turabian StyleKaronova, Tatiana L., Igor V. Kudryavtsev, Ksenia A. Golovatyuk, Arthur D. Aquino, Olga V. Kalinina, Alena T. Chernikova, Ekaterina K. Zaikova, Denis A. Lebedev, Ekaterina S. Bykova, Alexey S. Golovkin, and et al. 2022. "Vitamin D Status and Immune Response in Hospitalized Patients with Moderate and Severe COVID-19" Pharmaceuticals 15, no. 3: 305. https://doi.org/10.3390/ph15030305