
Lack of Evidence on Association between Iron Deficiency and COVID-19 Vaccine-Induced Neutralizing Humoral Immunity
 , , , , ,
, , , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Demographic Data
2.2. Hematological and Biochemistry Testing
2.3. Serological Assays
2.4. Statistical Analysis
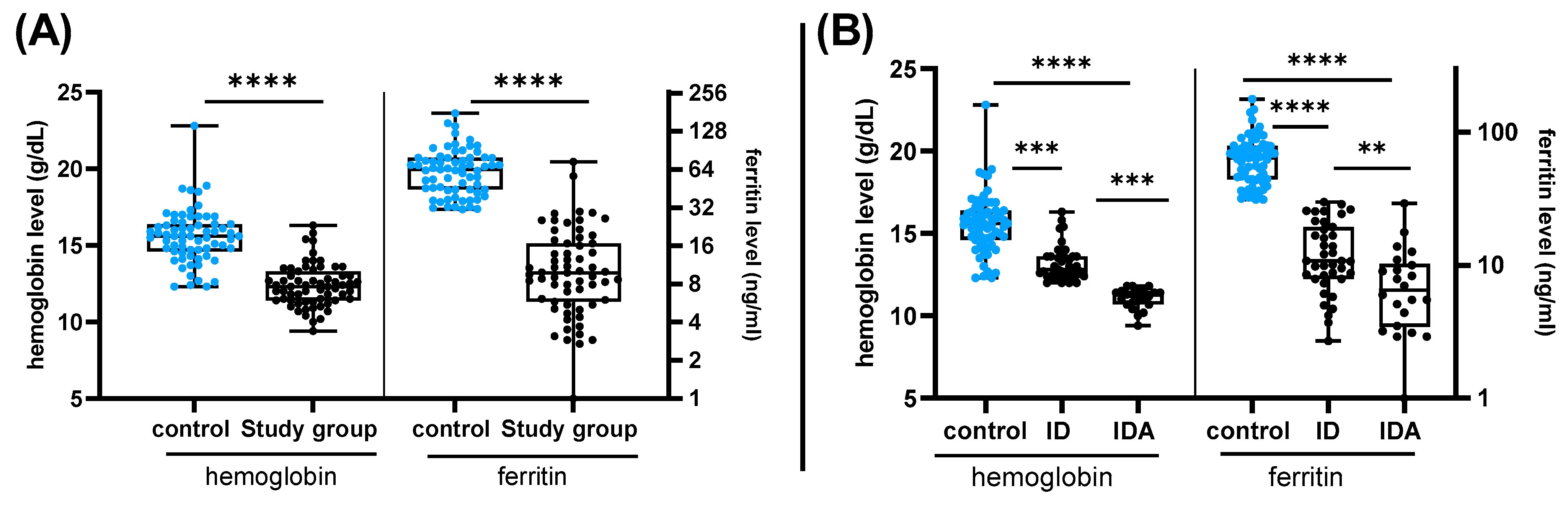
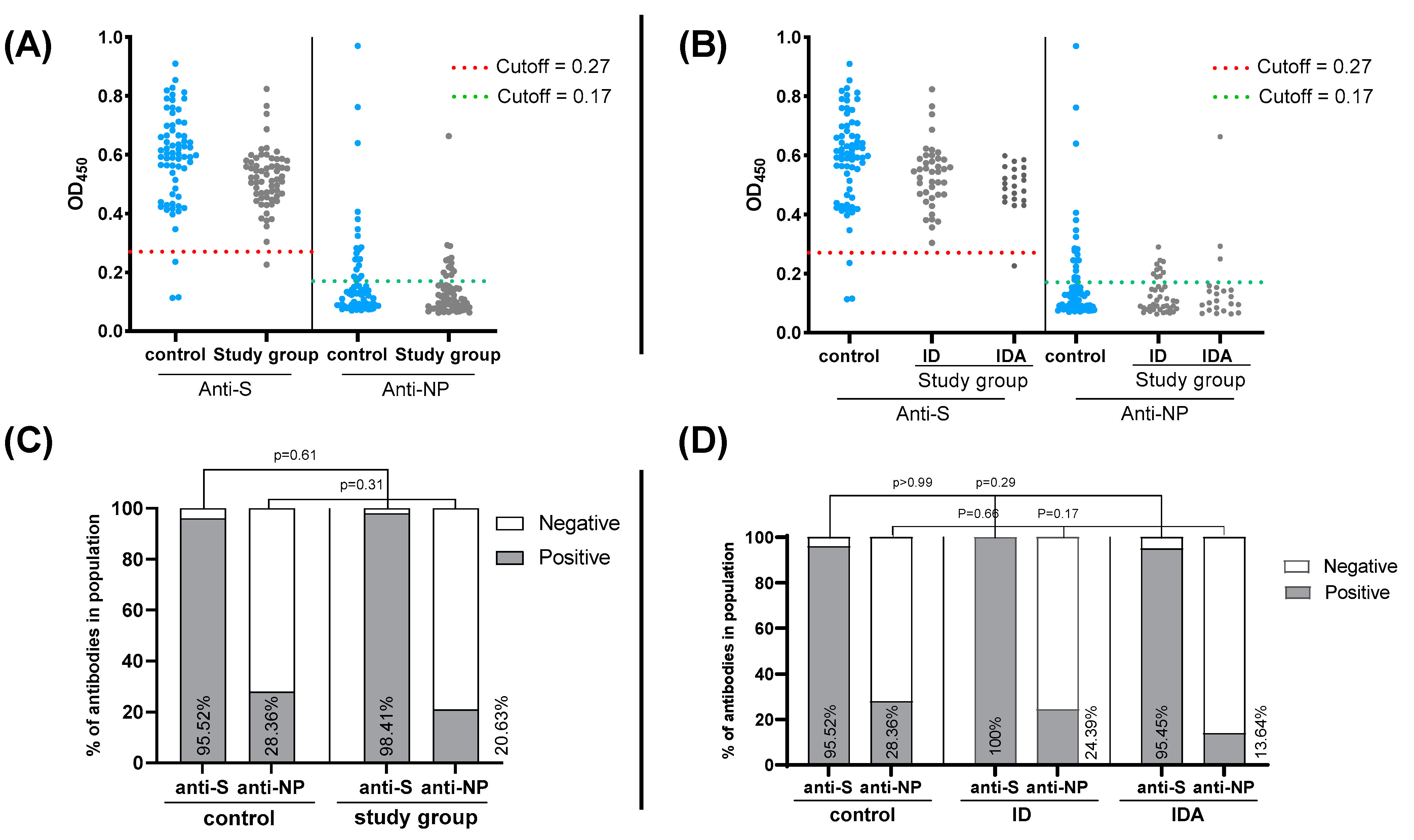
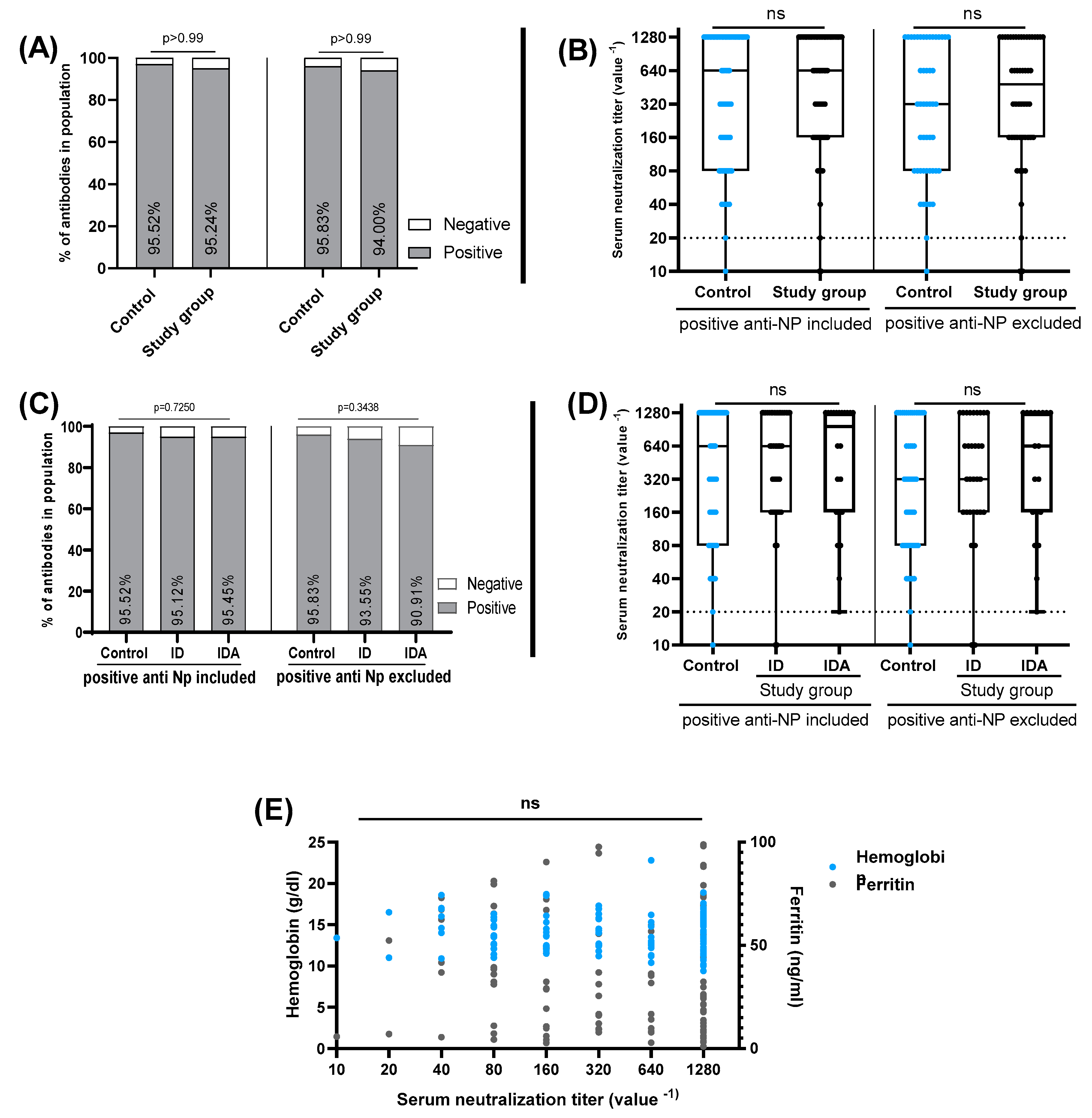
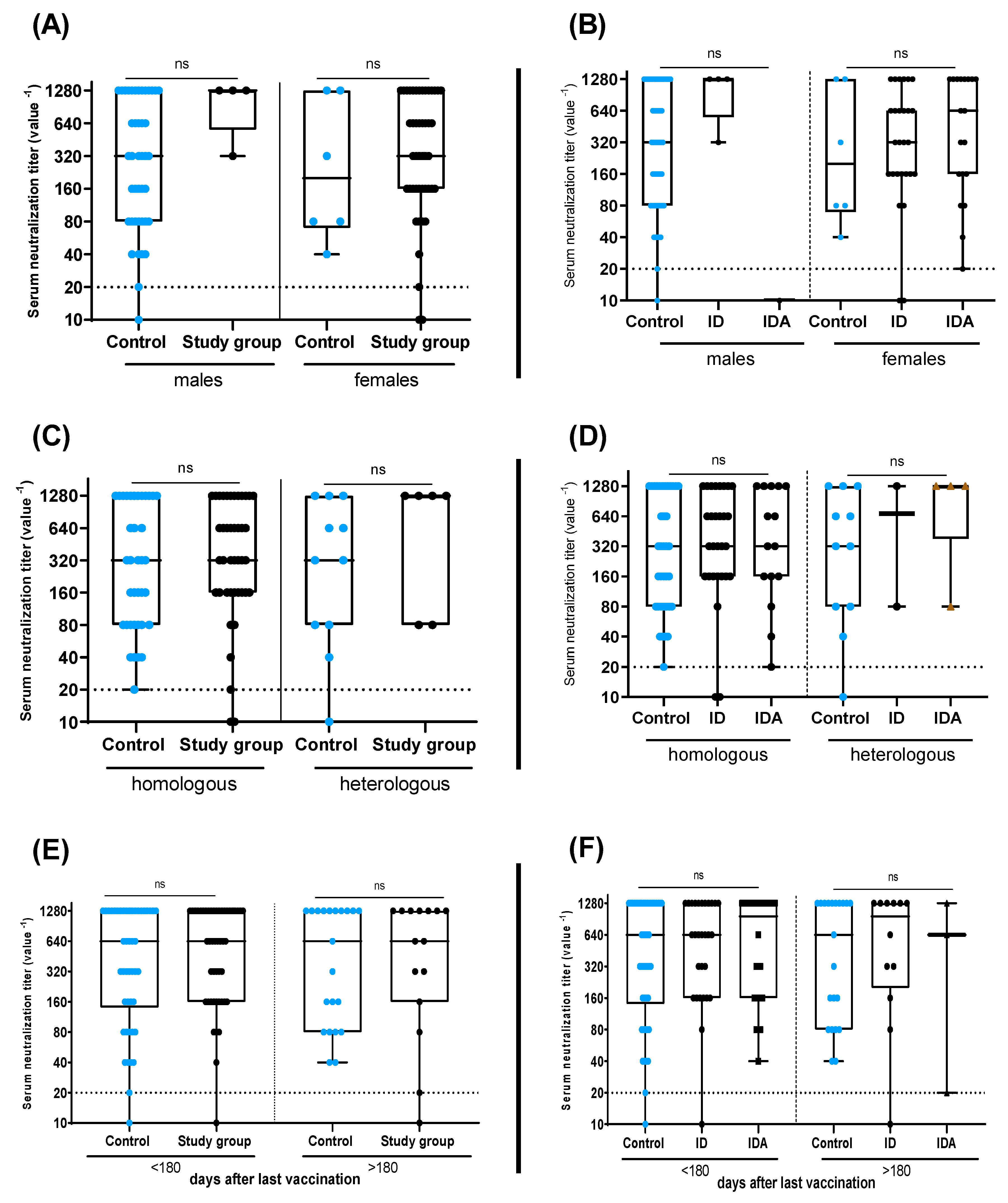
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmed, W.A.; Dada, A.; Alshukairi, A.N.; Sohrab, S.S.; Faizo, A.A.; Tolah, A.M.; El-Kafrawy, S.A.; Bajrai, L.H.; Moalim, H.M.; Aly, M.H.; et al. Seroprevalence of neutralizing antibodies to severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) among healthcare workers in Makkah, Saudi Arabia. J. King Saud. Univ. Sci. 2021, 33, 101366. [Google Scholar] [CrossRef] [PubMed]
- Alandijany, T.A.; El-Kafrawy, S.A.; Tolah, A.M.; Sohrab, S.S.; Faizo, A.A.; Hassan, A.M.; Alsubhi, T.L.; Othman, N.A.; Azhar, E.I. Development and Optimization of In-house ELISA for Detection of Human IgG Antibody to SARS-CoV-2 Full Length Spike Protein. Pathogens 2020, 9, 803. [Google Scholar] [CrossRef] [PubMed]
- Alandijany, T.A.; Faizo, A.A. Development of Serological Assays and Seroprevalence Studies of the New Coronavirus 2019 (COVID-19): Reports from Saudi Arabia. Healthcare 2021, 9, 1730. [Google Scholar] [CrossRef] [PubMed]
- Almarashda, A.M.; Rabbani, S.A.; Kurian, M.T.; Cherian, A. Clinical Characteristics, Risk Factors for Severity and Pharmacotherapy in Hospitalized COVID-19 Patients in the United Arab Emirates. J. Clin. Med. 2022, 11, 2439. [Google Scholar] [CrossRef]
- Anichini, G.; Terrosi, C.; Gori Savellini, G.; Gandolfo, C.; Franchi, F.; Cusi, M.G. Neutralizing Antibody Response of Vaccinees to SARS-CoV-2 Variants. Vaccines 2021, 9, 517. [Google Scholar] [CrossRef]
- Barefah, A.S.; Radhwi, O.O.; Alamri, S.S.; Alahwal, H.M.; Denetiu, I.; Almohammadi, A.T.; Bahashwan, S.M.; Qari, M.H.; Algaissi, A.; Alamer, E.; et al. Low clinical utility of testing for anti-platelet factor 4 in asymptomatic individuals after ChAdOx1 nCoV-19 vaccine. Int. J. Lab. Hematol. 2022, 44, 424–429. [Google Scholar] [CrossRef]
- Baronti, A.; Gentile, F.; Manetti, A.C.; Scatena, A.; Pellegrini, S.; Pucci, A.; Franzini, M.; Castiglione, V.; Maiese, A.; Giannoni, A.; et al. Myocardial Infarction Following COVID-19 Vaccine Administration: Post Hoc, Ergo Propter Hoc? Viruses 2022, 14, 1644. [Google Scholar] [CrossRef]
- Beard, J.L. Iron biology in immune function, muscle metabolism and neuronal functioning. J. Nutr. 2001, 131, 568S–580S. [Google Scholar] [CrossRef]
- Belali, T.M. Iron deficiency anaemia: Prevalence and associated factors among residents of northern Asir Region, Saudi Arabia. Sci. Rep. 2022, 12, 19170. [Google Scholar] [CrossRef]
- Bignucolo, A.; Scarabel, L.; Mezzalira, S.; Polesel, J.; Cecchin, E.; Toffoli, G. Sex Disparities in Efficacy in COVID-19 Vaccines: A Systematic Review and Meta-Analysis. Vaccines 2021, 9, 825. [Google Scholar] [CrossRef]
- Brüssow, H.; Sidoti, J.; Dirren, H.; Freire, W.B. Effect of malnutrition in Ecuadorian children on titers of serum antibodies to various microbial antigens. Clin. Diagn. Lab. Immunol. 1995, 2, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Camaschella, C. Iron-deficiency anemia. New Engl. J. Med. 2015, 372, 1832–1843. [Google Scholar] [CrossRef] [PubMed]
- Cherayil, B.J. Iron and Immunity: Immunological Consequences of Iron Deficiency and Overload. Arch. Immunol. Ther. Exp. 2010, 58, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Chiwona-Karltun, L.; Amuakwa-Mensah, F.; Wamala-Larsson, C.; Amuakwa-Mensah, S.; Abu Hatab, A.; Made, N.; Taremwa, N.K.; Melyoki, L.; Rutashobya, L.K.; Madonsela, T.; et al. COVID-19: From health crises to food security anxiety and policy implications. Ambio 2021, 50, 794–811. [Google Scholar] [CrossRef]
- Collins, H.L. The role of iron in infections with intracellular bacteria. Immunol. Lett. 2003, 85, 193–195. [Google Scholar] [CrossRef]
- Drakesmith, H.; Pasricha, S.R.; Cabantchik, I.; Hershko, C.; Weiss, G.; Girelli, D.; Stoffel, N.; Muckenthaler, M.U.; Nemeth, E.; Camaschella, C.; et al. Vaccine efficacy and iron deficiency: An intertwined pair? Lancet Haematol. 2021, 8, e666–e669. [Google Scholar] [CrossRef]
- Dufour, C.; Papadaki, H.; Warren, A. Expert Opinions for COVID-19 Vaccination in Patients with Non-Malignant Hematologic Diseases; European Hematology Association: Frankfurt, Germany, 2021. [Google Scholar]
- Faizo, A.A.; Alandijany, T.A.; Abbas, A.T.; Sohrab, S.S.; El-Kafrawy, S.A.; Tolah, A.M.; Hassan, A.M.; Azhar, E.I. A Reliable Indirect ELISA Protocol for Detection of Human Antibodies Directed to SARS-CoV-2 NP Protein. Diagnostics 2021, 11, 825. [Google Scholar] [CrossRef]
- Faizo, A.A.; Qashqari, F.S.; El-Kafrawy, S.A.; Barasheed, O.; Almashjary, M.N.; Alfelali, M.; Bawazir, A.A.; Albarakati, B.M.; Khayyat, S.A.; Hassan, A.M.; et al. A potential association between obesity and reduced effectiveness of COVID-19 vaccine-induced neutralizing humoral immunity. J. Med. Virol. 2022, 95, e28130. [Google Scholar] [CrossRef]
- Falahi, S.; Kenarkoohi, A. Host factors and vaccine efficacy: Implications for COVID-19 vaccines. J. Med. Virol. 2022, 94, 1330–1335. [Google Scholar] [CrossRef]
- Farsi, S.H.; Alandijany, T.A.; Radwi, M.; Farsi, A.; Bahaaziq, W.; Abushoshah, I.; El-Kafrawy, S.A.; Alsayed, S.; Azhar, E.I.; Boker, A. Prevalence of COVID-19 antibodies among operating room and critical care staff at a tertiary teaching hospital: A cross-sectional study. Saudi Med. J. 2021, 42, 742–749. [Google Scholar] [CrossRef]
- Frost, J.N.; Tan, T.K.; Abbas, M.; Wideman, S.K.; Bonadonna, M.; Stoffel, N.U.; Wray, K.; Kronsteiner, B.; Smits, G.; Campagna, D.R.; et al. Hepcidin-Mediated Hypoferremia Disrupts Immune Responses to Vaccination and Infection. Med 2021, 2, 164–179.e112. [Google Scholar] [CrossRef] [PubMed]
- Girelli, D.; Marchi, G.; Busti, F.; Vianello, A. Iron metabolism in infections: Focus on COVID-19. Semin. Hematol. 2021, 58, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Gualano, R.C.; Hansen, M.J.; Vlahos, R.; Jones, J.E.; Park-Jones, R.A.; Deliyannis, G.; Turner, S.J.; Duca, K.A.; Anderson, G.P. Cigarette smoke worsens lung inflammation and impairs resolution of influenza infection in mice. Respir. Res. 2008, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Hariyanto, T.I.; Kurniawan, A. Anemia is associated with severe coronavirus disease 2019 (COVID-19) infection. Transfus. Apher. Sci. 2020, 59, 102926. [Google Scholar] [CrossRef]
- Haschka, D.; Hoffmann, A.; Weiss, G. Iron in immune cell function and host defense. Semin. Cell Dev. Biol. 2021, 115, 27–36. [Google Scholar] [CrossRef]
- Hippchen, T.; Altamura, S.; Muckenthaler, M.U.; Merle, U. Hypoferremia is Associated with Increased Hospitalization and Oxygen Demand in COVID-19 Patients. HemaSphere 2020, 4, e492. [Google Scholar] [CrossRef]
- Hirai, K.; Shimotashiro, M.; Sonoda, T.; Okumura, T.; Ookawara, S.; Morishita, Y. Factors associated with anti-SARS-CoV-2 spike antibody titers after a second BNT162b2 mRNA COVID-19 vaccination in Japanese hemodialysis patients. Clin. Exp. Nephrol. 2022, 9, 925–932. [Google Scholar] [CrossRef]
- Jabara, H.H.; Boyden, S.E.; Chou, J.; Ramesh, N.; Massaad, M.J.; Benson, H.; Bainter, W.; Fraulino, D.; Rahimov, F.; Sieff, C.; et al. A missense mutation in TFRC, encoding transferrin receptor 1, causes combined immunodeficiency. Nat. Genet. 2016, 48, 74–78. [Google Scholar] [CrossRef]
- Jason, J.; Archibald, L.; Nwanyanwu, O.; Bell, M.; Jensen, R.; Gunter, E.; Buchanan, I.; Larned, J.; Kazembe, P.; Dobbie, H. The effects of iron deficiency on lymphocyte cytokine production and activation: Preservation of hepatic iron but not at all cost. Clin. Exp. Immunol. 2001, 126, 466–473. [Google Scholar] [CrossRef]
- Kassebaum, N.J.; Jasrasaria, R.; Naghavi, M.; Wulf, S.K.; Johns, N.; Lozano, R.; Regan, M.; Weatherall, D.; Chou, D.P.; Eisele, T.P.; et al. A systematic analysis of global anemia burden from 1990 to 2010. Blood 2014, 123, 615–624. [Google Scholar] [CrossRef]
- Lanser, L.; Burkert, F.R.; Bellmann-Weiler, R.; Schroll, A.; Wildner, S.; Fritsche, G.; Weiss, G. Dynamics in Anemia Development and Dysregulation of Iron Homeostasis in Hospitalized Patients with COVID-19. Metabolites 2021, 11, 653. [Google Scholar] [CrossRef]
- Mast, A.E.; Blinder, M.A.; Gronowski, A.M.; Chumley, C.; Scott, M.G. Clinical utility of the soluble transferrin receptor and comparison with serum ferritin in several populations. Clin. Chem. 1998, 44, 45–51. [Google Scholar] [CrossRef]
- Miller, J.L. Iron deficiency anemia: A common and curable disease. Cold Spring Harb. Perspect Med. 2013, 3, a011866. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol. Asp. Med. 2020, 75, 100864. [Google Scholar] [CrossRef] [PubMed]
- Oppenheimer, S.J. Iron and its relation to immunity and infectious disease. J. Nutr. 2001, 131, 616S–633S; discussion 633S–635S. [Google Scholar] [CrossRef] [PubMed]
- Radhwi, O.O.; Jan, H.; Waheeb, A.; Alamri, S.S.; Alahwal, H.M.; Denetiu, I.; Almanzlawey, A.; Al-Marzouki, A.F.; Almohammadi, A.T.; Bahashwan, S.M.; et al. Immunogenicity of The BNT162b2 COVID-19 mRNA and ChAdOx1 nCoV-19 Vaccines in Patients with Hemoglobinopathies. Vaccines 2022, 10, 151. [Google Scholar] [CrossRef]
- Santos-Araújo, C.; Mota Veiga, P.; Santos, M.J.; Santos, L.; Romãozinho, C.; Silva, M.; Lucas, C.; Duarte, M.L.; Haarhaus, M.; Haase, M.; et al. Time-dependent evolution of IgG antibody levels after first and second dose of mRNA-based SARS-CoV-2 vaccination in haemodialysis patients: A multicentre study. Nephrol. Dial. Transplant. 2022, 37, 375–381. [Google Scholar] [CrossRef]
- Self, W.H.; Tenforde, M.W.; Stubblefield, W.B.; Feldstein, L.R.; Steingrub, J.S.; Shapiro, N.I.; Ginde, A.A.; Prekker, M.E.; Brown, S.M.; Peltan, I.D.; et al. Decline in SARS-CoV-2 Antibodies after Mild Infection among Frontline Health Care Personnel in a Multistate Hospital Network—12 States, April–August 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1762–1766. [Google Scholar] [CrossRef]
- Stefan, N.; Birkenfeld, A.L.; Schulze, M.B. Global pandemics interconnected - obesity, impaired metabolic health and COVID-19. Nat. Reviews Endocrinol. 2021, 17, 135–149. [Google Scholar] [CrossRef]
- Stevens, G.A.; Finucane, M.M.; De-Regil, L.M.; Paciorek, C.J.; Flaxman, S.R.; Branca, F.; Peña-Rosas, J.P.; Bhutta, Z.A.; Ezzati, M. Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: A systematic analysis of population-representative data. Lancet Glob. Health 2013, 1, e16–e25. [Google Scholar] [CrossRef] [Green Version]
- Stoffel, N.U.; Uyoga, M.A.; Mutuku, F.M.; Frost, J.N.; Mwasi, E.; Paganini, D.; Van der Klis, F.R.; Malhotra, I.J.; LaBeaud, A.D.; Ricci, C. Iron deficiency anemia at time of vaccination predicts decreased vaccine response and iron supplementation at time of vaccination increases humoral vaccine response: A birth cohort study and a randomized trial follow-up study in Kenyan infants. Front. Immunol. 2020, 11, 1313. [Google Scholar] [CrossRef] [PubMed]
- Szczepanek, J.; Skorupa, M.; Goroncy, A.; Jarkiewicz-Tretyn, J.; Wypych, A.; Sandomierz, D.; Jarkiewicz-Tretyn, A.; Dejewska, J.; Ciechanowska, K.; Pałgan, K.; et al. Anti-SARS-CoV-2 IgG against the S Protein: A Comparison of BNT162b2, mRNA-1273, ChAdOx1 nCoV-2019 and Ad26.COV2.S Vaccines. Vaccines 2022, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Taneri, P.E.; Gómez-Ochoa, S.A.; Llanaj, E.; Raguindin, P.F.; Rojas, L.Z.; Roa-Díaz, Z.M.; Salvador, D., Jr.; Groothof, D.; Minder, B.; Kopp-Heim, D.; et al. Anemia and iron metabolism in COVID-19: A systematic review and meta-analysis. Eur. J. Epidemiol. 2020, 35, 763–773. [Google Scholar] [CrossRef]
- Wang, W.; Huang, B.; Zhu, Y.; Tan, W.; Zhu, M. Ferritin nanoparticle-based SARS-CoV-2 RBD vaccine induces a persistent antibody response and long-term memory in mice. Cell. Mol. Immunol. 2021, 18, 749–751. [Google Scholar] [CrossRef] [PubMed]
- Wendel, S.; Fontão-Wendel, R.; Fachini, R.; Candelaria, G.; Scuracchio, P.; Achkar, R.; Brito, M.; Reis, L.F.; Camargo, A.; Amano, M.; et al. A longitudinal study of convalescent plasma (CCP) donors and correlation of ABO group, initial neutralizing antibodies (nAb), and body mass index (BMI) with nAb and anti-nucleocapsid (NP) SARS-CoV-2 antibody kinetics: Proposals for better quality of CCP collections. Transfusion 2021, 61, 1447–1460. [Google Scholar] [CrossRef]
- Yathapu, S.R.; Kondapalli, N.B.; Srivalliputturu, S.B.; Hemalatha, R.; Bharatraj, D.K. Effect of lead exposure and nutritional iron-deficiency on immune response: A vaccine challenge study in rats. J. Immunotoxicol. 2020, 17, 144–152. [Google Scholar] [CrossRef]
- Yu, D.; Huo, J.; Xie, L.; Wang, L. Meta-analysis of studies on cut-off value of serum ferritin for identifying iron deficiency. J. Hyg. Res. 2013, 42, 228–235. [Google Scholar]
- Zhu, Z.; Xu, L.; Chen, G. Is there a difference in the efficacy of COVID-19 vaccine in males and females?—A systematic review and meta-analysis. Hum. Vaccines Immunother. 2021, 17, 4741–4746. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
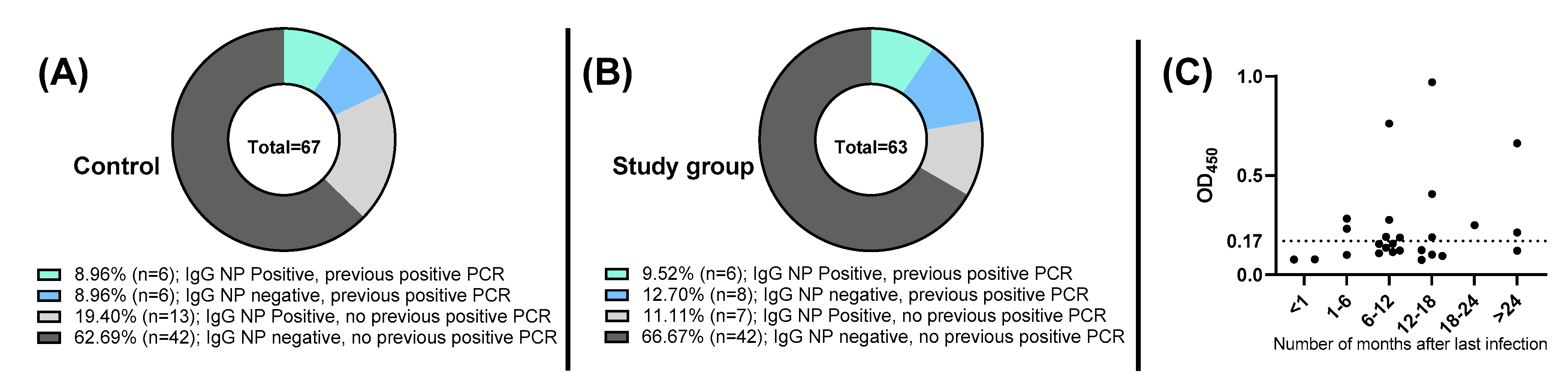{kind=link}
{kind=link}
{kind=link}
| Variables | Control (n = 67) | Study Group (n = 63) | p Value | |||
|---|---|---|---|---|---|---|
| Hemoglobin (g/dl) | 15.6 ± 1.74 | 12.39 ± 1.38 | <0.0001 | |||
| Ferritin (ng/mL) | 66.3 ± 29.13 | 13.3 ± 12.16 | <0.0001 | |||
| Age | 21.42 ± 2.48 | 22.47 ± 6.2 | 0.854 | |||
| Body mass index (BMI) | 25.54 ± 6.65 | 23.13 ± 5.29 | 0.0998 | |||
| Gender | Male | 60 | 89.55% | 5 | 7.94% | <0.0001 |
| Female | 7 | 10.45% | 58 | 92.06% | ||
| Previously diagnosed with COVID-19 by RT-PCR | Yes | 12 | 17.91% | 14 | 22.22% | 0.6617 |
| No | 55 | 82.09% | 49 | 77.78% | ||
| Vaccination | Homologous | 53 | 79.10% | 55 | 87.30% | 0.2477 |
| Pfizer | 48 | 71.64% | 53 | 84.13% | ||
| AstraZeneca | 5 | 7.46% | 2 | 3.17% | ||
| Heterologous | 14 | 20.90% | 8 | 12.70% | ||
| Pfizer/AstraZeneca | 14 | 20.90% | 7 | 11.11% | ||
| AstraZeneca/Moderna | 0 | 0.00% | 1 | 1.59% | ||
| Days since vaccination | <180 | 46 | 68.66% | 48 | 76.19% | 0.4332 |
| >180 | 21 | 31.34% | 15 | 23.81% | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faizo, A.A.; Bawazir, A.A.; Almashjary, M.N.; Hassan, A.M.; Qashqari, F.S.; Barefah, A.S.; El-Kafrawy, S.A.; Alandijany, T.A.; Azhar, E.I. Lack of Evidence on Association between Iron Deficiency and COVID-19 Vaccine-Induced Neutralizing Humoral Immunity. Vaccines 2023, 11, 327. https://doi.org/10.3390/vaccines11020327
Faizo AA, Bawazir AA, Almashjary MN, Hassan AM, Qashqari FS, Barefah AS, El-Kafrawy SA, Alandijany TA, Azhar EI. Lack of Evidence on Association between Iron Deficiency and COVID-19 Vaccine-Induced Neutralizing Humoral Immunity. Vaccines. 2023; 11(2):327. https://doi.org/10.3390/vaccines11020327
Chicago/Turabian StyleFaizo, Arwa A., Asma A. Bawazir, Majed N. Almashjary, Ahmed M. Hassan, Fadi S. Qashqari, Ahmed S. Barefah, Sherif A. El-Kafrawy, Thamir A. Alandijany, and Esam I. Azhar. 2023. "Lack of Evidence on Association between Iron Deficiency and COVID-19 Vaccine-Induced Neutralizing Humoral Immunity" Vaccines 11, no. 2: 327. https://doi.org/10.3390/vaccines11020327