
Antifungal Drug Susceptibility and Genetic Characterization of Fungi Recovered from COVID-19 Patients

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Culture Conditions
2.2. DNA Extraction
2.3. Antifungal Susceptibility Testing
2.4. Sequencing of FKS1 and ERG11
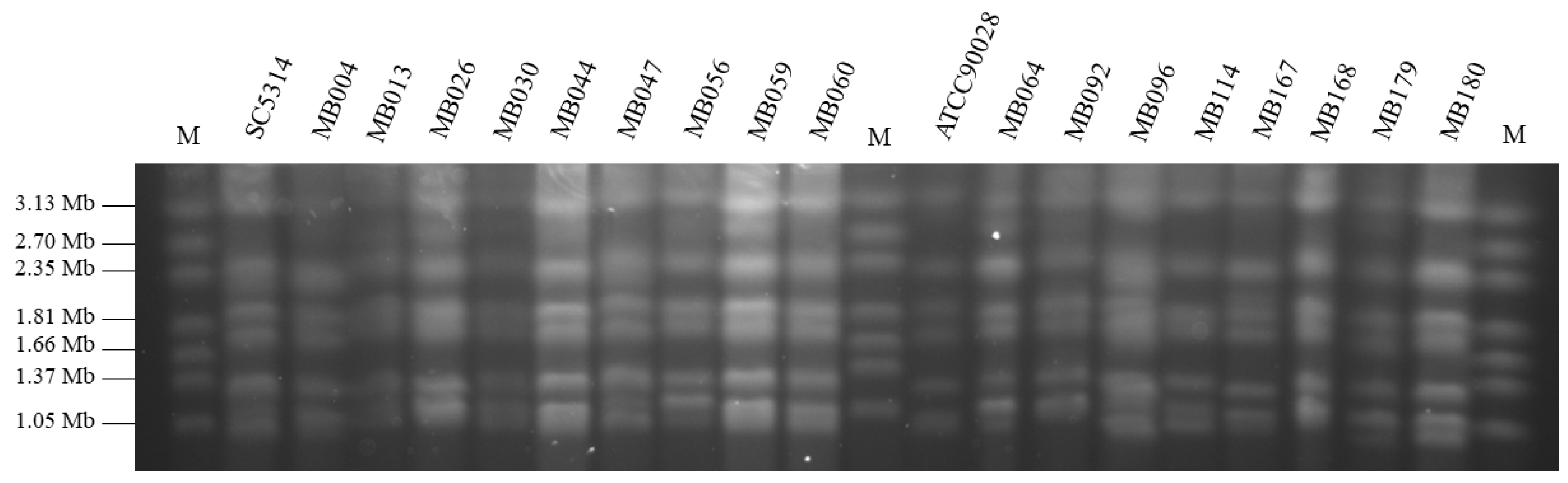
2.5. Karyotyping of C. albicans Isolates
2.6. Multilocus Sequence Typing (MLST) of C. albicans Isolates
2.7. Typing of A. fumigatus Isolates
2.8. Clinical Data Review
3. Results
3.1. Species Distribution of Fungal Isolates Recovered from COVID-19 Patients
3.2. Antifungal Susceptibility Testing and Molecular Resistance Determinants Analysis
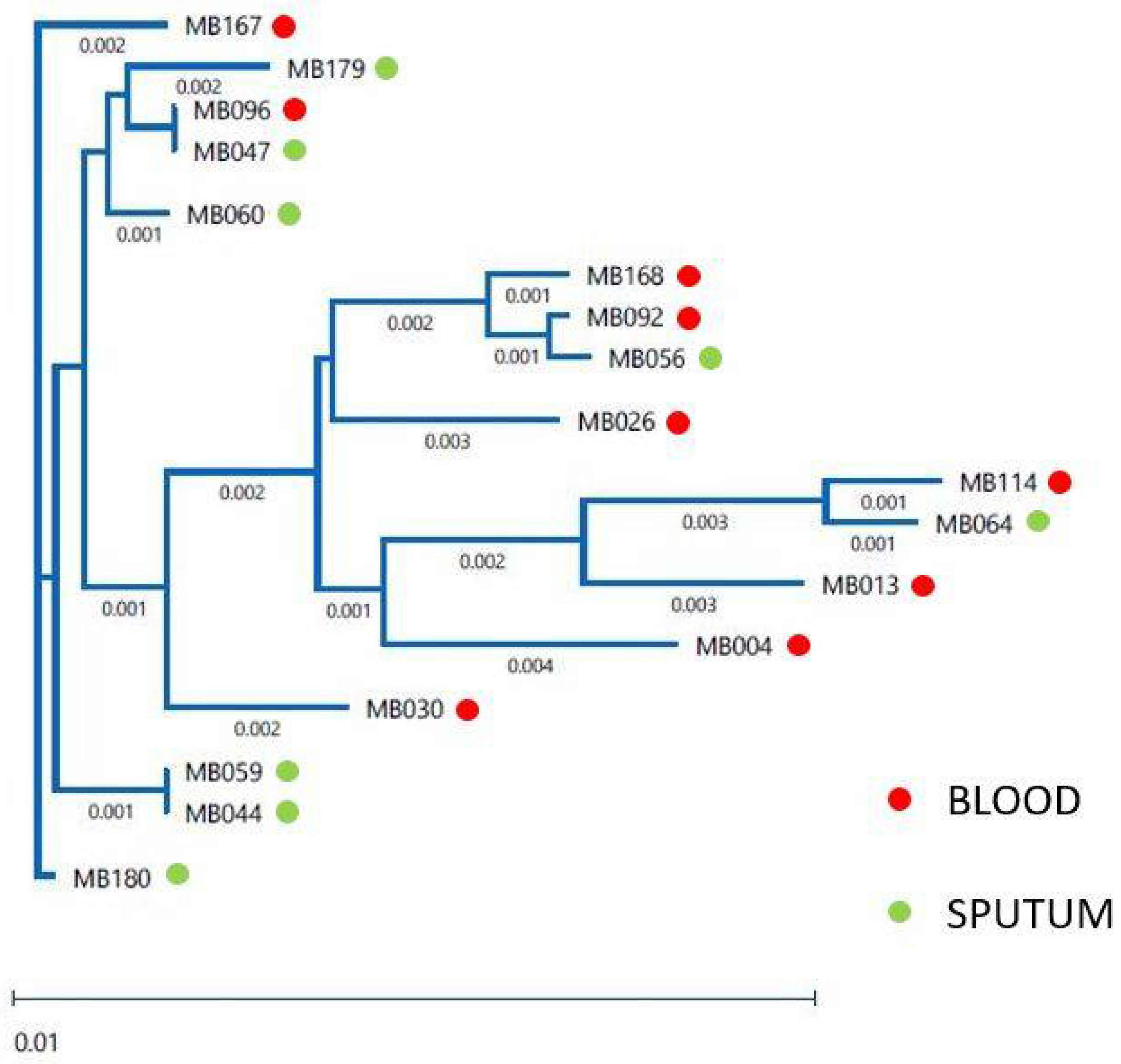
3.3. Relatedness of the Clinical Isolates
3.4. Clinical Data Review
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. WHO Coronavirus Disease (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 6 May 2021).
- Liderot, K.; Ahl, M.; Ozenci, V. Secondary bacterial infections in patients with seasonal influenza A and pandemic H1N1. BioMed Res. Int. 2013, 2013, 376219. [Google Scholar] [CrossRef] [PubMed]
- Rijnders, B.J.A.; Schauwvlieghe, A.; Wauters, J. Influenza-associated pulmonary aspergillosis: A local or global lethal combination? Clin. Infect. Dis. 2020, 71, 1764–1767. [Google Scholar] [CrossRef]
- Zheng, Z.G.; Chen, R.C.; Li, Y.M. The clinical characteristics of secondary infection of lower respiratory in severe acute respiratory syndrome. Chin. J. Respir. Crit. Care Med. 2003, 2, 270–274. [Google Scholar]
- Hwang, D.M.; Chamberlain, D.W.; Poutanen, S.M.; Low, D.E.; Asa, S.L.; Butany, J. Pulmonary pathology of severe acute respiratory syndrome in Toronto. Mod. Pathol. 2005, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.P.; Wei, R.; Verhoef, J. Real time assay of Aspergillus should be used in SARS patients receiving corticosteroids. BMJ 2003, 327, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Goyal, P.; Choi, J.J.; Pinheiro, L.C.; Schenck, E.J.; Chen, R.; Jabri, A.; Satlin, M.J.; Campion, T.R., Jr.; Nahid, M.; Ringel, J.B.; et al. Clinical Characteristics of Covid-19 in New York City. N. Engl. J. Med. 2020, 382, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vidal, C.; Sanjuan, G.; Moreno-Garcia, E.; Puerta-Alcalde, P.; Garcia-Pouton, N.; Chumbita, M.; Fernandez-Pittol, M.; Pitart, C.; Inciarte, A.; Bodro, M.; et al. Incidence of co-infections and superinfections in hospitalized patients with COVID-19: A retrospective cohort study. Clin. Microbiol. Infect. 2020, 27, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Nucci, M.; Barreiros, G.; Guimaraes, L.F.; Deriquehem, V.A.S.; Castineiras, A.C.; Nouer, S.A. Increased incidence of candidemia in a tertiary care hospital with the COVID-19 pandemic. Mycoses 2021, 64, 152–156. [Google Scholar] [CrossRef]
- Mastrangelo, A.; Germinario, B.N.; Ferrante, M.; Frangi, C.; Li Voti, R.; Muccini, C.; Ripa, M.; COVID-BioB Study Group. Candidemia in COVID-19 patients: Incidence and characteristics in a prospective cohort compared to historical non-COVID-19 controls. Clin. Infect. Dis. 2020, ciaa1594. [Google Scholar] [CrossRef] [PubMed]
- Villanueva-Lozano, H.; Trevino-Rangel, R.J.; Gonzalez, G.M.; Ramirez-Elizondo, M.T.; Lara-Medrano, R.; Aleman-Bocanegra, M.C.; Guajardo-Lara, C.E.; Gaona-Chavez, N.; Castilleja-Leal, F.; Torre-Amione, G.; et al. Outbreak of Candida auris infection in a COVID-19 hospital in Mexico. Clin. Microbiol. Infect. 2021, 27, 813–816. [Google Scholar] [CrossRef]
- Posteraro, B.; Torelli, R.; Vella, A.; Leone, P.M.; De Angelis, G.; De Carolis, E.; Ventura, G.; Sanguinetti, M.; Fantoni, M. Pan-Echinocandin-Resistant Candida glabrata Bloodstream Infection Complicating COVID-19: A Fatal Case Report. J. Fungi 2020, 6, 163. [Google Scholar] [CrossRef] [PubMed]
- Alanio, A.; Delliere, S.; Fodil, S.; Bretagne, S.; Megarbane, B. Prevalence of putative invasive pulmonary aspergillosis in critically ill patients with COVID-19. Lancet Respir. Med. 2020, 8, e48–e49. [Google Scholar] [CrossRef]
- Clancy, C.J.; Nguyen, M.H. COVID-19, superinfections and antimicrobial development: What can we expect? Clin. Infect. Dis. 2020, 71, 2736–2743. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yu, Y.; Xu, J.; Shu, H.; Xia, J.; Liu, H.; Wu, Y.; Zhang, L.; Yu, Z.; Fang, M.; et al. Clinical course and outcomes of critically ill patients with SARS-CoV-2 pneumonia in Wuhan, China: A single-centered, retrospective, observational study. Lancet Respir. Med. 2020, 8, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Tu, L.; Zhu, P.; Mu, M.; Wang, R.; Yang, P.; Wang, X.; Hu, C.; Ping, R.; Hu, P.; et al. Clinical Features of 85 Fatal Cases of COVID-19 from Wuhan. A Retrospective Observational Study. Am. J. Respir. Crit. Care Med. 2020, 201, 1372–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arentz, M.; Yim, E.; Klaff, L.; Lokhandwala, S.; Riedo, F.X.; Chong, M.; Lee, M. Characteristics and Outcomes of 21 Critically Ill Patients With COVID-19 in Washington State. JAMA 2020, 323, 1612–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Grasselli, G.; Zangrillo, A.; Zanella, A.; Antonelli, M.; Cabrini, L.; Castelli, A.; Cereda, D.; Coluccello, A.; Foti, G.; Fumagalli, R.; et al. Baseline Characteristics and Outcomes of 1591 Patients Infected With SARS-CoV-2 Admitted to ICUs of the Lombardy Region, Italy. JAMA 2020, 323, 1574–1581. [Google Scholar] [CrossRef] [Green Version]
- Brillowska-Dabrowska, A.; Nielsen, S.S.; Nielsen, H.V.; Arendrup, M.C. Optimized 5-hour multiplex PCR test for the detection of tinea unguium: Performance in a routine PCR laboratory. Med. Mycol. 2010, 48, 828–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeast. In CLSI Standard M27, 4th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Performance Standards for Antifungal Susceptibility Testing of Yeasts. In CLSI Supplement M60, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. In CLSI Standard M38, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Performance Standards for Antifungal Susceptibility Testing of Filamentous Fungi. In CLSI Supplement M61, 2nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Mellado, E.; Garcia-Effron, G.; Alcazar-Fuoli, L.; Melchers, W.J.; Verweij, P.E.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. A new Aspergillus fumigatus resistance mechanism conferring in vitro cross-resistance to azole antifungals involves a combination of cyp51A alterations. Antimicrob. Agents Chemother. 2007, 51, 1897–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, F.Y.; Liang, H.Y.; Jia, J.H. Taxonomic relationships among the taxa in the Candida guilliermondii complex, as revealed by comparative electrophoretic karyotyping. Int. J. Syst. Evol. Microbiol. 2000, 50, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Bougnoux, M.E.; Tavanti, A.; Bouchier, C.; Gow, N.A.; Magnier, A.; Davidson, A.D.; Maiden, M.C.; D’Enfert, C.; Odds, F.C. Collaborative consensus for optimized multilocus sequence typing of Candida albicans. J. Clin. Microbiol. 2003, 41, 5265–5266. [Google Scholar] [CrossRef] [Green Version]
- Odds, F.C.; Bougnoux, M.E.; Shaw, D.J.; Bain, J.M.; Davidson, A.D.; Diogo, D.; Jacobsen, M.D.; Lecomte, M.; Li, S.Y.; Tavanti, A.; et al. Molecular phylogenetics of Candida albicans. Eukaryot. Cell 2007, 6, 1041–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Rubio, R.; Escribano, P.; Gomez, A.; Guinea, J.; Mellado, E. Comparison of Two Highly Discriminatory Typing Methods to Analyze Aspergillus fumigatus Azole Resistance. Front. Microbiol. 2018, 9, 1626. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Mellado, E.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Aspergillus section Fumigati: Antifungal susceptibility patterns and sequence-based identification. Antimicrob. Agents Chemother. 2008, 52, 1244–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Garcia-Effron, G.; Lass-Florl, C.; Lopez, A.G.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Perlin, D.S. Echinocandin susceptibility testing of Candida species: Comparison of EUCAST EDef 7.1, CLSI M27-A3, Etest, disk diffusion, and agar dilution methods with RPMI and isosensitest media. Antimicrob. Agents Chemother. 2010, 54, 426–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Effron, G.; Katiyar, S.K.; Park, S.; Edlind, T.D.; Perlin, D.S. A naturally occurring proline-to-alanine amino acid change in Fks1p in Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis accounts for reduced echinocandin susceptibility. Antimicrob. Agents. Chemother. 2008, 52, 2305–2312. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, Y.; Wu, J.; Li, Y.; Zhou, X.; Li, X.; Chen, H.; Guo, M.; Chen, S.; Sun, F.; et al. Risks and features of secondary infections in severe and critical ill COVID-19 patients. Emerg. Microbes Infect. 2020, 9, 1958–1964. [Google Scholar] [CrossRef]
- Lamoth, F.; Glampedakis, E.; Boillat-Blanco, N.; Oddo, M.; Pagani, J.L. Incidence of invasive pulmonary aspergillosis among critically ill COVID-19 patients. Clin. Microbiol. Infect. 2020, 26, 1706–1708. [Google Scholar] [CrossRef] [PubMed]
- Rutsaert, L.; Steinfort, N.; Van Hunsel, T.; Bomans, P.; Naesens, R.; Mertes, H.; Dits, H.; Van Regenmortel, N. COVID-19-associated invasive pulmonary aspergillosis. Ann. Intensive Care 2020, 10, 71. [Google Scholar] [CrossRef] [PubMed]
- White, P.L.; Dhillon, R.; Cordey, A.; Hughes, H.; Faggian, F.; Soni, S.; Pandey, M.; Whitaker, H.; May, A.; Morgan, M.; et al. A national strategy to diagnose COVID-19 associated invasive fungal disease in the ICU. Clin. Infect. Dis. 2020, ciaa1298. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Tarai, B.; Singh, A.; Sharma, A. Multidrug-Resistant Candida auris Infections in Critically Ill Coronavirus Disease Patients, India, April-July 2020. Emerg. Infect. Dis. 2020, 26, 2694–2696. [Google Scholar] [CrossRef] [PubMed]
- Peman, J.; Ruiz-Gaitan, A.; Garcia-Vidal, C.; Salavert, M.; Ramirez, P.; Puchades, F.; Garcia-Hita, M.; Alastruey-Izquierdo, A.; Quindos, G. Fungal co-infection in COVID-19 patients: Should we be concerned? Rev. Iberoam. Micol. 2020, 37, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R.; Jackson, B.R.; Vallabhaneni, S.; Ostrosky-Zeichner, L.; Pappas, P.G.; Chiller, T. Thinking beyond the Common Candida Species: Need for Species-Level Identification of Candida Due to the Emergence of Multidrug-Resistant Candida auris. J. Clin. Microbiol. 2017, 55, 3324–3327. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yu, Y.T.; Xu, C.H.; Chen, D.C. Candida Colonization in the Respiratory Tract: What Is the Significance? Front. Med. 2020, 7, 598037. [Google Scholar] [CrossRef] [PubMed]
- Mulet Bayona, J.V.; Tormo Palop, N.; Salvador Garcia, C.; Fuster Escriva, B.; Chanza Avino, M.; Ortega Garcia, P.; Gimeno Cardona, C. Impact of the SARS-CoV-2 Pandemic in Candidaemia, Invasive Aspergillosis and Antifungal Consumption in a Tertiary Hospital. J. Fungi 2021, 7, 440. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.S.; Mohsin, J.; Al-Huraizi, A.; Khamis, F. COVID-19 associated invasive candidiasis. J. Infect. 2021, 82, e45–e46. [Google Scholar] [CrossRef] [PubMed]
- Gorkem, A.; Sav, H.; Kaan, O.; Eren, E. Coronavirus disease and candidemia infection: A case report. J. Mycol. Med. 2021, 31, 101155. [Google Scholar] [CrossRef] [PubMed]
- Ventoulis, I.; Sarmourli, T.; Amoiridou, P.; Mantzana, P.; Exindari, M.; Gioula, G.; Vyzantiadis, T.A. Bloodstream Infection by Saccharomyces cerevisiae in Two COVID-19 Patients after Receiving Supplementation of Saccharomyces in the ICU. J. Fungi 2020, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Meijer, E.F.J.; Dofferhoff, A.S.M.; Hoiting, O.; Buil, J.B.; Meis, J.F. Azole-Resistant COVID-19-Associated Pulmonary Aspergillosis in an Immunocompetent Host: A Case Report. J. Fungi 2020, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Philbin, M.; Hughes, G.; Bergin, C.; Talento, A.F. Antimicrobial stewardship challenges and innovative initiatives in the acute hospital setting during the COVID-19 pandemic. J. Antimicrob. Chemother. 2021, 76, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Adler, H.; Ball, R.; Fisher, M.; Mortimer, K.; Vardhan, M.S. Low rate of bacterial co-infection in patients with COVID-19. Lancet Microbe 2020, 1, e62. [Google Scholar] [CrossRef]
- Chen, T.; Wu, D.; Chen, H.; Yan, W.; Yang, D.; Chen, G.; Ma, K.; Xu, D.; Yu, H.; Wang, H.; et al. Clinical characteristics of 113 deceased patients with coronavirus disease 2019: Retrospective study. BMJ 2020, 368, m1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Cox, M.J.; Loman, N.; Bogaert, D.; O’Grady, J. Co-infections: Potentially lethal and unexplored in COVID-19. Lancet Microbe 2020, 1, e11. [Google Scholar] [CrossRef]
- Zhou, P.; Liu, Z.; Chen, Y.; Xiao, Y.; Huang, X.; Fan, X.G. Bacterial and fungal infections in COVID-19 patients: A matter of concern. Infect. Control Hosp. Epidemiol. 2020, 41, 1124–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perea, S.; Lopez-Ribot, J.L.; Kirkpatrick, W.R.; McAtee, R.K.; Santillan, R.A.; Martinez, M.; Calabrese, D.; Sanglard, D.; Patterson, T.F. Prevalence of molecular mechanisms of resistance to azole antifungal agents in Candida albicans strains displaying high-level fluconazole resistance isolated from human immunodeficiency virus-infected patients. Antimicrob. Agents Chemother. 2001, 45, 2676–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, S.A.; Colon, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of clinically derived mutations in ERG11 to azole resistance in Candida albicans. Antimicrob. Agents Chemother. 2015, 59, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, Y.; Zhao, Y.; Hu, X.; Cai, Z.; Liu, X.; Jin, G.; Zhang, J.; Zhang, J.; Liu, J.; Huang, X. In vitro fluconazole susceptibility of 1,903 clinical isolates of Candida albicans and the identification of ERG11 mutations. Microb. Drug Resist. 2013, 19, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, L.; Li, C. Susceptibility of clinical isolates of Candida species to fluconazole and detection of Candida albicans ERG11 mutations. J. Antimicrob. Chemother. 2008, 61, 798–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Boyken, L.; Hollis, R.J.; Kroeger, J.; Messer, S.A.; Tendolkar, S.; Diekema, D.J. In vitro susceptibility of clinical isolates of Aspergillus spp. to anidulafungin, caspofungin, and micafungin: A head-to-head comparison using the CLSI M38-A2 broth microdilution method. J. Clin. Microbiol. 2009, 47, 3323–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Boyken, L.; Hollis, R.J.; Kroeger, J.; Messer, S.A.; Tendolkar, S.; Diekema, D.J. Wild-type minimum effective concentration distributions and epidemiologic cutoff values for caspofungin and Aspergillus spp. as determined by Clinical and Laboratory Standards Institute broth microdilution methods. Diagn. Microbiol. Infect. Dis. 2010, 67, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Leonardelli, F.; Macedo, D.; Dudiuk, C.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Aspergillus fumigatus Intrinsic Fluconazole Resistance Is Due to the Naturally Occurring T301I Substitution in Cyp51Ap. Antimicrob. Agents Chemother. 2016, 60, 5420–5426. [Google Scholar] [CrossRef] [Green Version]
- McManus, B.A.; Coleman, D.C. Molecular epidemiology, phylogeny and evolution of Candida albicans. Infect. Genet. Evol. 2014, 21, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Selmecki, A.; Forche, A.; Berman, J. Aneuploidy and isochromosome formation in drug-resistant Candida albicans. Science 2006, 313, 367–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchonville, K.; Forche, A.; Tang, K.E.; Selmecki, A.; Berman, J. Aneuploid chromosomes are highly unstable during DNA transformation of Candida albicans. Eukaryot. Cell 2009, 8, 1554–1566. [Google Scholar] [CrossRef] [Green Version]
- Forche, A.; Magee, P.T.; Selmecki, A.; Berman, J.; May, G. Evolution in Candida albicans populations during a single passage through a mouse host. Genetics 2009, 182, 799–811. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Chen, Y.; Duan, L.; Zhao, J.; Qin, C.; Li, H.; Sun, J.; Han, L. Comparison of Two Typing Methods for Characterization of Azole Resistance in Aspergillus fumigatus from Potting Soil Samples in a Chinese Hospital. Antimicrob. Agents Chemother. 2020, 64, e01578-19. [Google Scholar] [CrossRef] [PubMed]
- Donato, M.L.; Park, S.; Baker, M.; Korngold, R.; Morawski, A.; Geng, X.; Tan, M.; Ip, A.; Goldberg, S.; Rowley, S.; et al. Clinical and laboratory evaluation of patients with SARS-CoV-2 pneumonia treated with high-titer convalescent plasma. JCI Insight 2021, 6, e143196. [Google Scholar] [CrossRef] [PubMed]
- Arastehfar, A.; Shaban, T.; Zarrinfar, H.; Roudbary, M.; Ghazanfari, M.; Hedayati, M.T.; Sedaghat, A.; Ilkit, M.; Najafzadeh, M.J.; Perlin, D.S. Candidemia among Iranian Patients with Severe COVID-19 Admitted to ICUs. J. Fungi 2021, 7, 280. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.L.; Lima, C.M.; Magalhaes, V.C.R.; Baltazar, L.M.; Peres, N.T.A.; Caligiorne, R.B.; Moura, A.S.; Fereguetti, T.; Martins, J.C.; Rabelo, L.F.; et al. Fungal and bacterial coinfections increase mortality of severely ill COVID-19 patients. J. Hosp. Infect. 2021, 113, 145–154. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate | Specimen | Identification | MIC [mg/L] * | FKS1 | Erg11/Cyp51A | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Original (at HUMC) | Species Confirmation (rDNA seq) | MCF | FLC | VRC | ITR | POS | ISA | ||||
| MB 004 | Blood | C. albicans | C. albicans | <0.03 | <0.25 | 0.03 | <0.03 | <0.03 | <0.03 | WT | E266D |
| MB 013 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | 0.03 | 0.03 | WT | V437I |
| MB 026 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | <0.03 | <0.03 | <0.03 | WT | E266D, H283H/L, K342K/R |
| MB 030 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | V437V/I |
| MB 044 | Sputum | yeast | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | K128K/T |
| MB 047 | Sputum | yeast | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | K128K/T |
| MB 056 | Sputum | yeast | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | V437V/I |
| MB 059 | Sputum | yeast (not Cryptococcus) | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | K128K/T |
| MB 060 | Sputum | yeast | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | K128K/T |
| MB 064 | Sputum | yeast (not Cryptococcus) | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | D116D/E, E266E/D |
| MB 092 | Blood | C. albicans | C. albicans | <0.03 | <0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | V437V/I |
| MB 096 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | D116E, K128K/T |
| MB 114 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | E266E/D, V437V/I |
| MB 167 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | D116E, K128T |
| MB 168 | Blood | C. albicans | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | V437V/I |
| MB 179 | Sputum | yeast (not Cryptococcus) | C. albicans | <0.03 | 0.25 | 0.03 | 0.03 | <0.03 | <0.03 | WT | D116D/E, K128K/T, V159V/I |
| MB 180 | Sputum | yeast (not Cryptococcus) | C. albicans | <0.03 | <0.25 | 0.06 | <0.03 | <0.03 | <0.03 | WT | D116D/E, K128K/T |
| MB 025 | Blood | C. parapsilosis | C. parapsilosis | 2 | 0.5 | 0.125 | <0.03 | <0.03 | <0.03 | WT ** | F215S |
| MB 099 | Blood | C. parapsilosis | C. parapsilosis | 2 | 0.5 | 0.125 | <0.03 | <0.03 | <0.03 | WT ** | F215S |
| MB 097 | Sputum | A. fumigatus | A. fumigatus | 0.06 | >64 | 0.25 | 0.5 | 0.03 | 0.125 | N/A | WT |
| MB 098 | Sputum | A. fumigatus | A. fumigatus | 0.06 | >64 | 0.25 | 0.5 | 0.03 | 0.125 | N/A | WT |
| Gene | Organism | Primer Name | Sequence | Use | Reference |
|---|---|---|---|---|---|
| rDNA | Candida spp. | Fun-rDNAF | GGTCATTTAGAGGAAGTAAAAGTCG | PCR + seq | S. Katiyar, personal communication |
| Fun-rDNAR | YGATATGCTTAAGTTCAGCGGGTA | PCR + seq | |||
| β-tubulin | Aspergillus spp. | btub2F | TTCACCTTCAGACCGGT | PCR + seq | [33] |
| btub4R | AGTTGTCGGGACGGAATAG | PCR + seq | |||
| FKS1 | C. albicans | F2426 | CATTGCTGTGGCCACTTTAG | PCR + seq | [34] |
| R2919 | GATTTCCATTTCCGTGGTAGC | PCR | |||
| F4590 | TACTATGGTCATCCAGGTTTCC | PCR + seq | |||
| R4954 | GGTCAAATCAGTGAAAACCG | PCR | |||
| C. parapsilosis | CparF | CTCCAAGTCCTCATATGCAC | PCR + seq | ||
| CparR | AGATGTTTCTCCATGGTGTC | PCR | |||
| F4500 | AAGATTGGTGCTGGTATGGG | PCR + seq | [35] | ||
| R5112 | TAATGGTGCTTGCCAATGAG | PCR | |||
| ERG11 | C. albicans | Ca_ERG11_F | ATG GCT ATT GTT GAA ACT GTC ATT G | PCR | This study * |
| Ca_ERG11_R | TTA AAA CAT ACA AGT TTC TCT TTT TTC CC | PCR | |||
| Ca_ERG11_731-750_F | GGA GAC GTG ATG CTG CTC AA | seq | |||
| Ca_ERG11_918-938_R | GCA GAA GTA TGT TGA CCA CCC | seq | |||
| C. parapsilosis | Cpara_erg11_F1 | TCCCTACCTTCGTTCATC | PCR + seq | ||
| Cpara_erg11_R1 | CGAGGTGAGTCAACAAAG | PCR + seq | |||
| Cpara_erg11_F2 | AGAGACGGGTGACATTG | seq | |||
| Cpara_erg11_R2 | TGGCACTAGTATGCTGTC | seq | |||
| CYP51A promoter | A. fumigatus | A5 | CTT TTT CGA CTG CCG CGC | PCR + seq | [28] |
| A7 | TCA TAT GTT GCT CAG CGG | PCR + seq | |||
| CYP51A | A. fumigatus | P450.1 | ATG GTG CCG ATG CTA TGG | PCR | |
| P450.2 | CTG TCT CAC TTG GAT GTG | PCR | |||
| CypA1 | CTT ACG GCC TAC ATG GCC | seq | |||
| CypA2 | TTC GAC CGC TTC TCC CAG | seq | |||
| A3 | TAG TCC ATT GAC GAC CCC | seq | |||
| TRESPERG genotyping | A. fumigatus | CSP1F | TTGGGTGGCATTGTGCCAA | PCR + seq | [32] |
| CSP2R | GAGCATGACAACCCAGATACCA | PCR + seq | |||
| MP2A | ATGCGGTTCTCTGCGTTA | PCR | |||
| MP2B | CAGCAACAGTGCAAATGC | PCR | |||
| MP2_P1 | CTCGAACTTGGCTACGAC | seq | |||
| MP2_P2 | AGGTAGTGGAGGTCACTG | seq | |||
| CFEMA | ATGAAGGCCTCTGTGTC | PCR + seq | |||
| CFEMB | AGGATAATCAAGGCAGCG | PCR + seq | |||
| ERG4B_P1 | ATGACTGTCACACGCTCC | PCR + seq | |||
| ERG4B_P2 | TAGACGGCACCAATCCAC | PCR + seq |
| Isolate | Specimen | AAT1a | ACC1 | ADP1 | MPI | SYA1 | VPS13 | ZWF1b | ST |
|---|---|---|---|---|---|---|---|---|---|
| MB004 | Blood | 43 | 14 | 8 | 4 | 7 | 10 | 8 | 927 |
| MB013 | Blood | 117 | 7 | 21 | 34 | * 238 | 24 | 6 | New ST, A |
| MB026 | Blood | 25 | 7 | 6 | 3 | 6 | 27 | 37 | 90 |
| MB030 | Blood | 20 | 3 | 6 | 2 | 51 | 132 | 5 | New ST, B |
| MB044 | Sputum | 2 | 5 | 5 | 2 | 2 | 6 | 20 | 485 |
| MB047 | Sputum | 8 | 5 | * 6 | 2 | 2 | 6 | 5 | New ST, C |
| MB056 | Sputum | 33 | 14 | 38 | 2 | * 136 | 122 | 15 | New ST, D |
| MB059 | Sputum | 2 | 5 | 5 | 2 | 2 | 6 | 20 | 485 |
| MB060 | Sputum | 8 | 3 | 6 | 2 | 2 | 6 | 49 | New ST, E |
| MB064 | Sputum | 13 | 7 | 15 | 6 | 7 | 55 | 15 | 1830 |
| MB092 | Blood | 33 | 7 | 6 | 2 | 78 | 122 | 15 | New ST, F |
| MB096 | Blood | 8 | 5 | * 6 | 2 | 2 | 6 | 5 | New ST, C |
| MB114 | Blood | 13 | 10 | 15 | 6 | 7 | 15 | 15 | New ST, G |
| MB167 | Blood | 2 | 2 | 5 | 2 | 2 | 68 | 5 | New ST, H |
| MB168 | Blood | 33 | 3 | 38 | 2 | 78 | 122 | 22 | New ST, I |
| MB179 | Sputum | 8 | 2 | 5 | 9 | 2 | 6 | 5 | 285 |
| MB180 | Sputum | 2 | 3 | 5 | 2 | 2 | 6 | 5 | 66 |
| Characteristic | Values |
|---|---|
| Age | 60 (55–69) (49–84) |
| BMI | 29.3 (24.96–37.34) (22.91–58.5) |
| ≥30 | 8 (42.1%) |
| Sex | |
| Male | 13 (68.4%) |
| Female | 6 (31.6%) |
| Comorbidities | |
| Diabetes | 7 (36.8%) |
| Asthma | 3 (15.8%) |
| COPD | 1 (5.3%) |
| Hypertension | 12 (63.2%) |
| Cancer | 0 |
| Hypercholesterolemia | 5 (26.3%) |
| Hyperlipidemia | 2 (10.5%) |
| Arthritis | 4 (21.1%) |
| Treatment | |
| Antibiotics | 19 (100%) |
| Antifungals | 12 (63.2%) |
| Corticosteroids | 18 (94.7%) |
| Hydroxychloroquine | 14 (73.7%) |
| Remdesivir | 3 (15.8%) |
| Lopinavir-ritonavir | 1 (5.3%) |
| Sarilumab | 1 (5.3%) |
| Fungal culture | |
| Day of the first blood culture | 14 (11–19) (8–28) |
| Day of the first sputum culture | 12 (6–18) (1–40) |
| Fungal culture only | 6 (31.6%) |
| Fungal and bacterial culture | 13 (68.4%) |
| Hospitalization | |
| Days of hospital stay | 29 (15–33) (9–81) |
| ICU admission | 6 (31.6%) |
| Days in ICU | 24 (8.25–41.25) (0–54) |
| Ventilation | 18 (94.7%) * |
| Days on ventilator | 24 (11–31.25) (7–81) |
| Death | 17 (89.5%) |
| Respiratory failure | 14/17 (82.3%) |
| Cardiac failure | 2/17 (11.8%) |
| No data | 1/17 (5.9%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kordalewska, M.; Guerrero, K.D.; Garcia-Rubio, R.; Jiménez-Ortigosa, C.; Mediavilla, J.R.; Cunningham, M.H.; Hollis, F.; Hong, T.; Chow, K.F.; Kreiswirth, B.N.; et al. Antifungal Drug Susceptibility and Genetic Characterization of Fungi Recovered from COVID-19 Patients. J. Fungi 2021, 7, 552. https://doi.org/10.3390/jof7070552
Kordalewska M, Guerrero KD, Garcia-Rubio R, Jiménez-Ortigosa C, Mediavilla JR, Cunningham MH, Hollis F, Hong T, Chow KF, Kreiswirth BN, et al. Antifungal Drug Susceptibility and Genetic Characterization of Fungi Recovered from COVID-19 Patients. Journal of Fungi. 2021; 7(7):552. https://doi.org/10.3390/jof7070552
Chicago/Turabian StyleKordalewska, Milena, Kevin D. Guerrero, Rocio Garcia-Rubio, Cristina Jiménez-Ortigosa, José R. Mediavilla, Marcus H. Cunningham, Frank Hollis, Tao Hong, Kar Fai Chow, Barry N. Kreiswirth, and et al. 2021. "Antifungal Drug Susceptibility and Genetic Characterization of Fungi Recovered from COVID-19 Patients" Journal of Fungi 7, no. 7: 552. https://doi.org/10.3390/jof7070552