
Lineage 1 Porcine Reproductive and Respiratory Syndrome Virus Attenuated Live Vaccine Provides Broad Cross-Protection against Homologous and Heterologous NADC30-Like Virus Challenge in Piglets


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statements
2.2. Cells and Viruses
2.3. Phylogenetic and Genomic Recombination Analysis
2.4. Viral Culturing and Attenuation
2.5. Whole-Genome Sequencing of Lineage 1 PRRSV SD at Different Passages
2.6. Safety Evaluation of High-Dose and Repeated-Dose of SD-R
2.7. Evaluation of Immunoprotection of SD-R against Homologous and Heterologous Strains
2.8. Serological Examination
2.9. Viremia and Viral Loads in Tissue Assessment
2.10. Histological Examination
2.11. Statistical Analysis
3. Results
3.1. Genomic Characteristics of NADC30-like PRRSV SD and HLJWK108-1711
3.2. Nucleotide and Amino Acid Mutations of Different SD Passages
3.3. Both High-Dose and Repeated-Dose Tests of SD-R Are Safe for Piglets
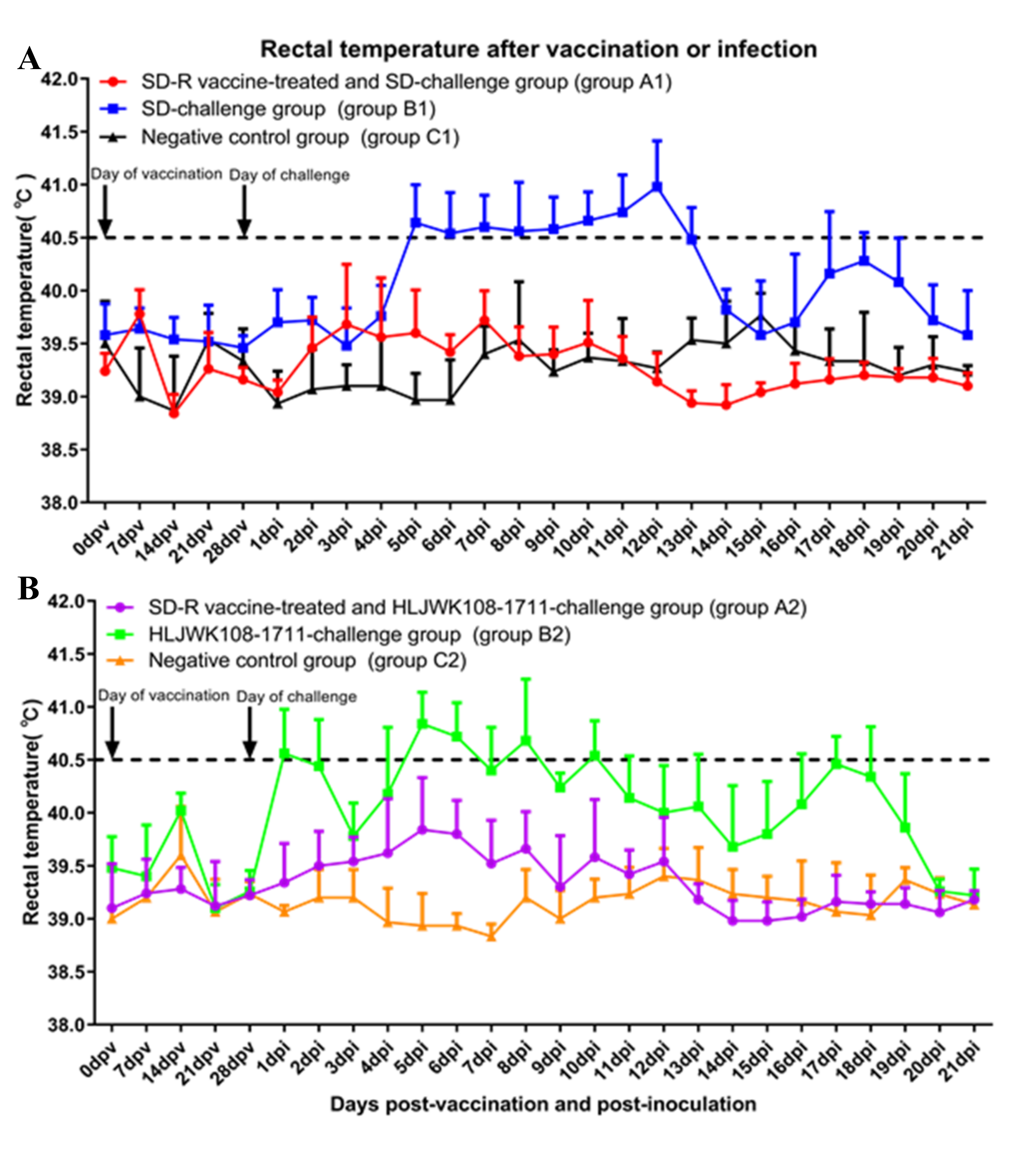
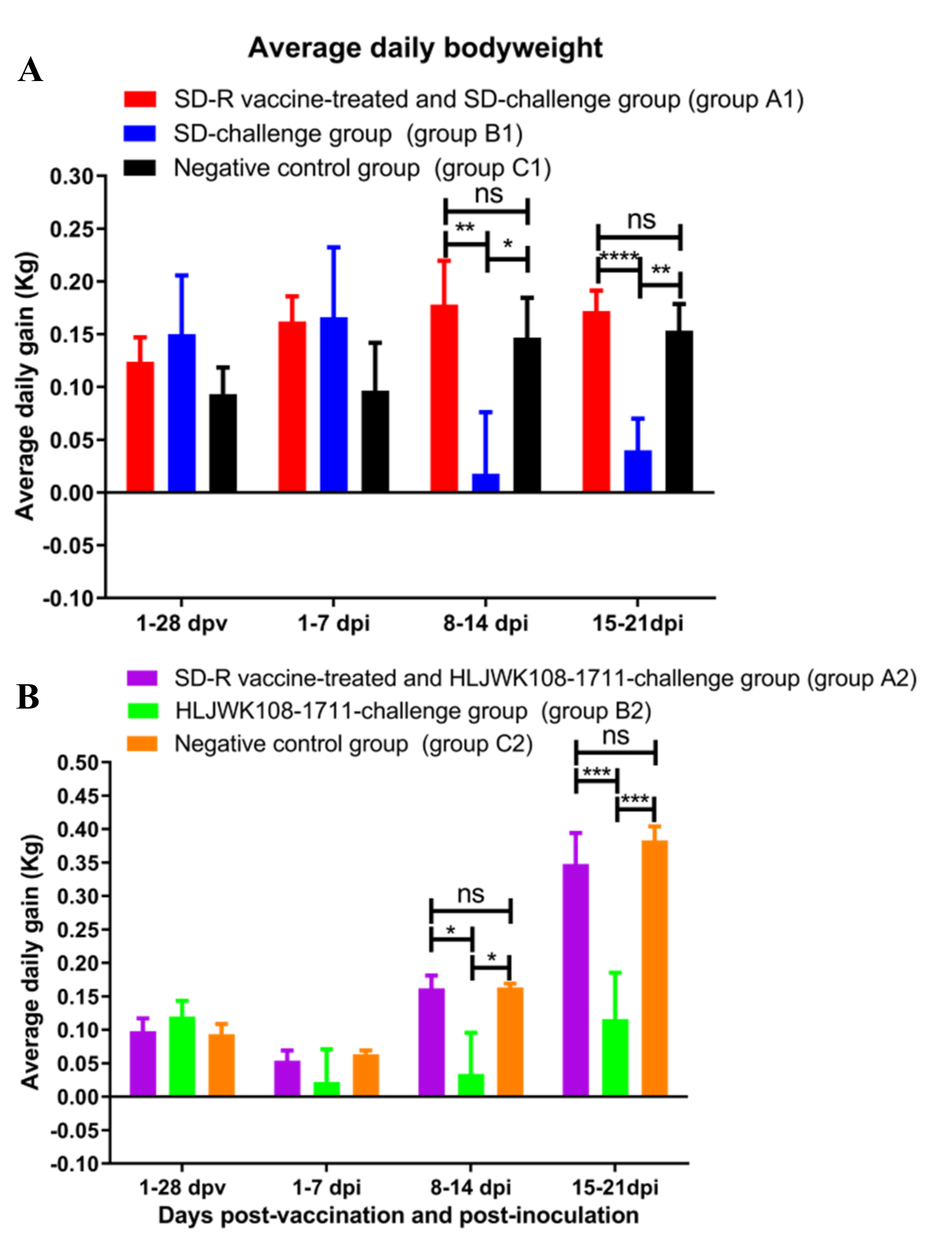
3.4. Clinical Reactions after Immunization and Challenge
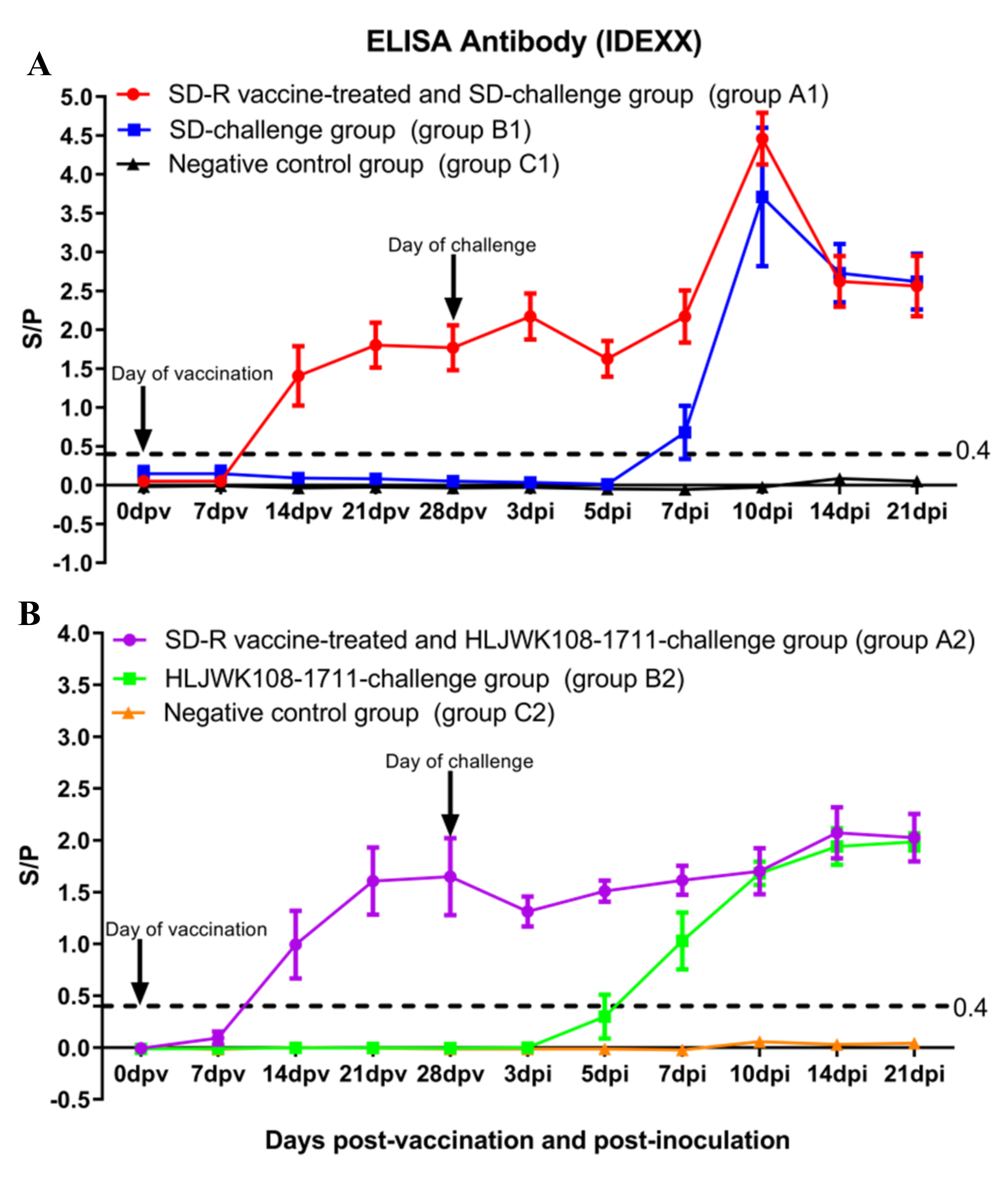
3.5. Antibody Responses in Immunized or Challenged Piglets
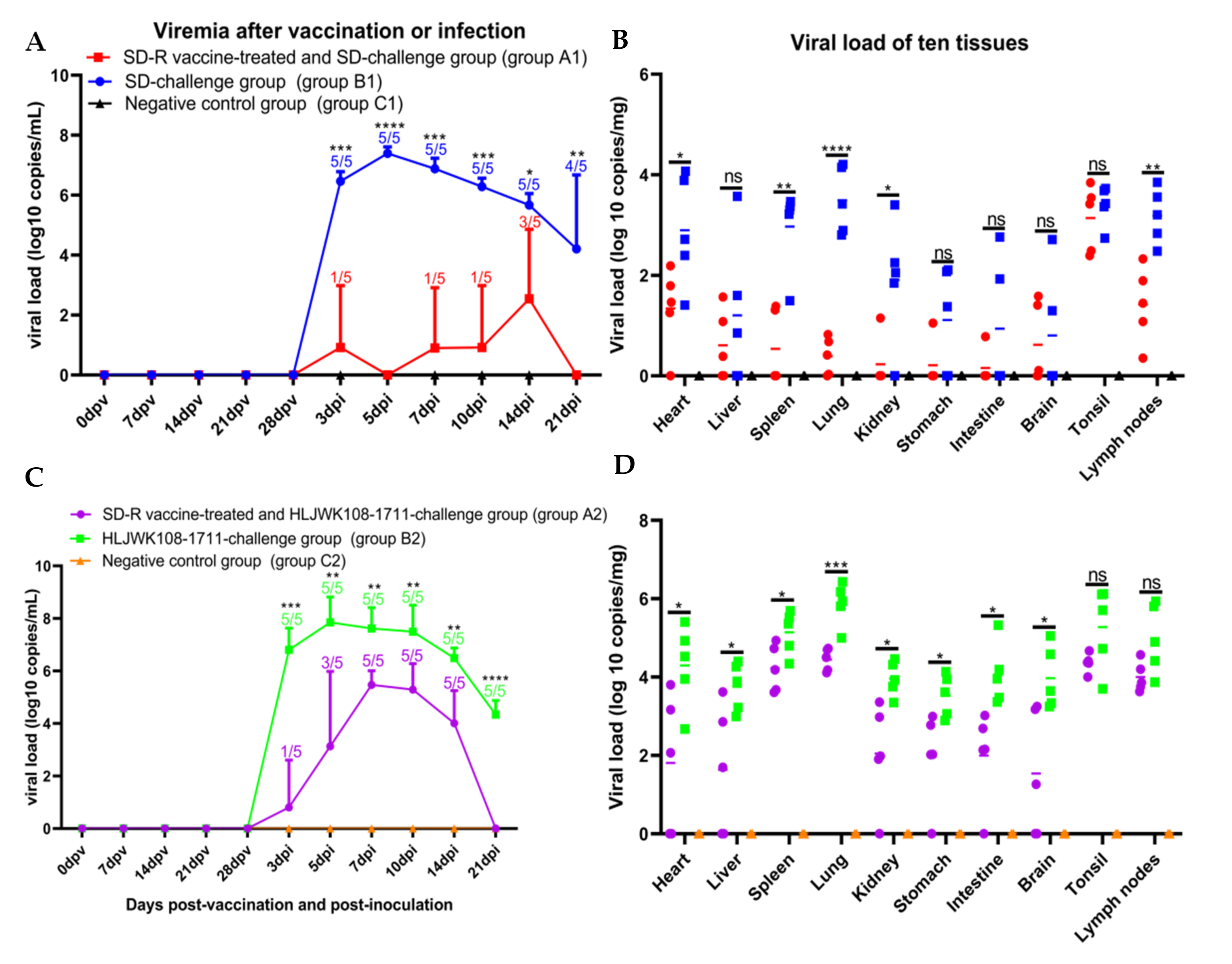
3.6. Viremia and Viral Tissue Distribution between the Immunized-Challenge Group and the Challenge Group
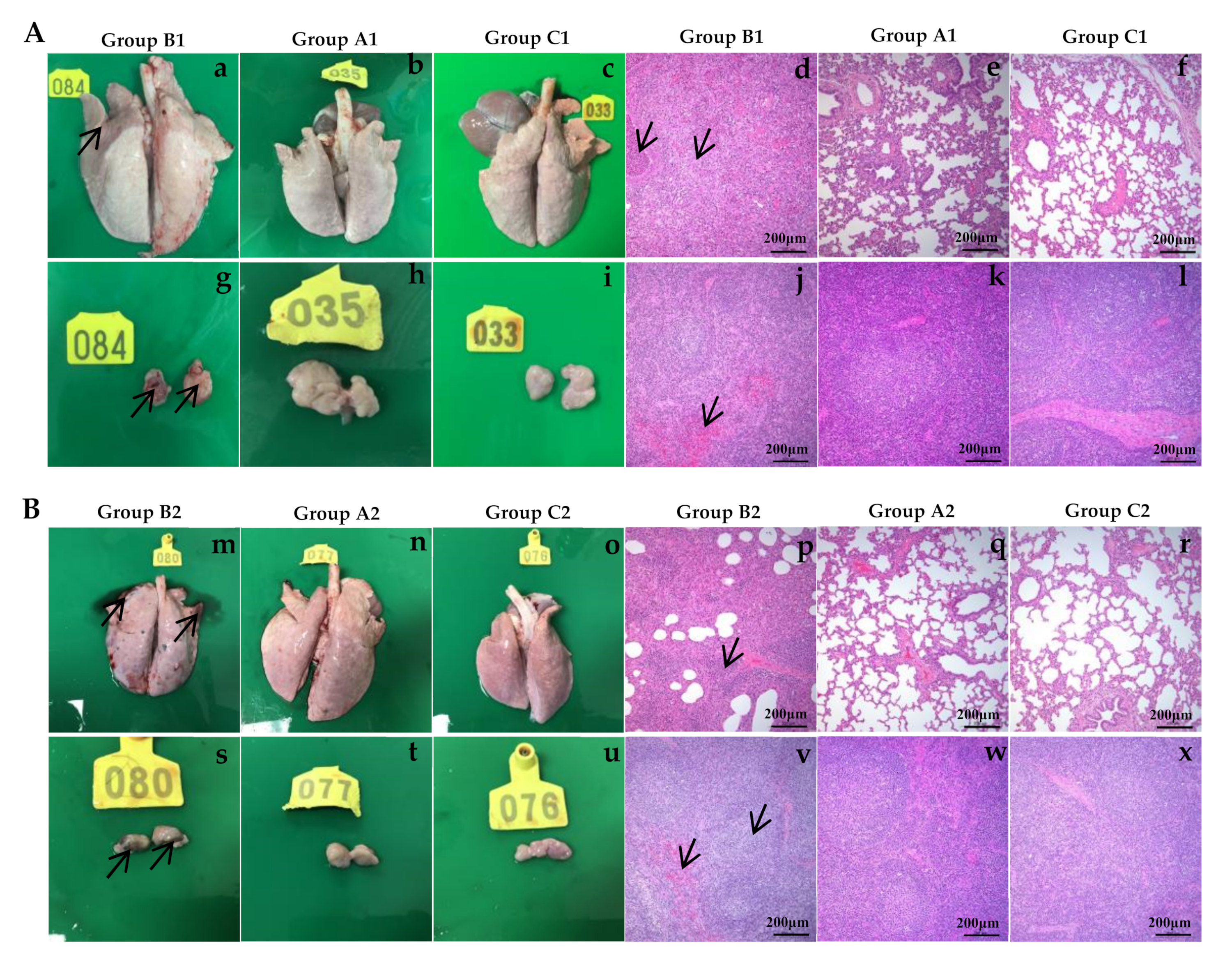
3.7. Gross Pathological and Histopathological Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V.; Nathues, C. Cost of porcine reproductive and respiratory syndrome virus at individual farm level—An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Stadejek, T.; Stankevicius, A.; Murtaugh, M.P.; Oleksiewicz, M.B. Molecular evolution of PRRSV in Europe: Current state of play. Vet. Microbiol. 2013, 165, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Hui, R.K.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus. Res. 2010, 154, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Schneider, P.; Zhang, W.P.; Faaberg, K.S.; Nelson, E.A.; Rowland, R.R. Diversity and evolution of a newly emerged North American Type 1 porcine arterivirus: Analysis of isolates collected between 1999 and 2004. Arch. Virol. 2007, 152, 1009–1017. [Google Scholar] [CrossRef]
- Ropp, S.L.; Wees, C.E.; Fang, Y.; Nelson, E.A.; Rossow, K.D.; Bien, M.; Arndt, B.; Preszler, S.; Steen, P.; Christopher-Hennings, J.; et al. Characterization of emerging European-like porcine reproductive and respiratory syndrome virus isolates in the United States. J. Virol. 2004, 78, 3684–3703. [Google Scholar] [CrossRef] [Green Version]
- Dewey, C.; Charbonneau, G.; Carman, S.; Hamel, A.; Nayar, G.; Friendship, R.; Eernisse, K.; Swenson, S. Lelystad-like strain of porcine reproductive and respiratory syndrome virus (PRRSV) identified in Canadian swine. Can. Vet. J. 2000, 41, 493–494. [Google Scholar]
- Lee, C.; Kim, H.; Kang, B.; Yeom, M.; Han, S.; Moon, H.; Park, S.; Kim, H.; Song, D.; Park, B. Prevalence and phylogenetic analysis of the isolated type I porcine reproductive and respiratory syndrome virus from 2007 to 2008 in Korea. Virus. Genes 2010, 40, 225–230. [Google Scholar] [CrossRef]
- Thanawongnuwech, R.; Amonsin, A.; Tatsanakit, A.; Damrongwatanapokin, S. Genetics and geographical variation of porcine reproductive and respiratory syndrome virus (PRRSV) in Thailand. Vet. Microbiol. 2004, 101, 9–21. [Google Scholar] [CrossRef]
- Chen, N.; Cao, Z.; Yu, X.; Deng, X.; Zhao, T.; Wang, L.; Liu, Q.; Li, X.; Tian, K. Emergence of novel European genotype porcine reproductive and respiratory syndrome virus in mainland China. J. Gen. Virol. 2011, 92, 880–892. [Google Scholar] [CrossRef]
- Deng, M.C.; Chang, C.Y.; Huang, T.S.; Tsai, H.J.; Chang, C.; Wang, F.I.; Huang, Y.L. Molecular epidemiology of porcine reproductive and respiratory syndrome viruses isolated from 1991 to 2013 in Taiwan. Arch. Virol. 2015, 160, 2709–2718. [Google Scholar] [CrossRef] [PubMed]
- Chueh, L.L.; Lee, K.H.; Wang, F.I.; Pang, V.F.; Weng, C.N. Sequence analysis of the nucleocapsid protein gene of the porcine reproductive and respiratory syndrome virus Taiwan MD-001 strain. Adv. Exp. Med. Biol. 1998, 440, 795–799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Zhang, H.L.; Xu, H.; Tang, Y.D.; Leng, C.L.; Peng, J.M.; Wang, Q.; An, T.Q.; Cai, X.H.; Fan, J.H.; et al. Two novel recombinant porcine reproductive and respiratory syndrome viruses belong to sublineage 3.5 originating from sublineage 3.2. Transbound. Emerg. Dis. 2019, 66, 2592–2600. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.K.; Chen, Y.J.; Cai, Y.; Li, Q.; Xie, J.X.; Liang, G.; Gao, Q.; Yu, Z.Q.; Lu, G.; Huang, L.Z.; et al. Insights into the evolutionary history and epidemiological characteristics of the emerging lineage 1 porcine reproductive and respiratory syndrome viruses in China. Transbound. Emerg. Dis. 2020, 67, 2630–2641. [Google Scholar] [CrossRef] [PubMed]
- Paploski, I.A.D.; Pamornchainavakul, N.; Makau, D.N.; Rovira, A.; Corzo, C.A.; Schroeder, D.C.; Cheeran, M.C.; Doeschl-Wilson, A.; Kao, R.R.; Lycett, S.; et al. Phylogenetic Structure and Sequential Dominance of Sub-Lineages of PRRSV Type-2 Lineage 1 in the United States. Vaccines 2021, 9, 608. [Google Scholar] [CrossRef]
- Yu, F.; Yan, Y.; Shi, M.; Liu, H.Z.; Zhang, H.L.; Yang, Y.B.; Huang, X.Y.; Gauger, P.C.; Zhang, J.; Zhang, Y.H.; et al. Phylogenetics, Genomic Recombination, and NSP2 Polymorphic Patterns of Porcine Reproductive and Respiratory Syndrome Virus in China and the United States in 2014–2018. J. Virol. 2020, 94, e01813–e01819. [Google Scholar] [CrossRef]
- Makau, D.N.; Alkhamis, M.A.; Paploski, I.A.D.; Corzo, C.A.; Lycett, S.; VanderWaal, K. Integrating animal movements with phylogeography to model the spread of PRRSV in the USA. Virus. Evol. 2021, 7, veab060. [Google Scholar] [CrossRef]
- Makau, D.N.; Paploski, I.A.D.; Corzo, C.A.; VanderWaal, K. Dynamic network connectivity influences the spread of a sub-lineage of porcine reproductive and respiratory syndrome virus. Transbound. Emerg. Dis. 2021, 69, 524–537. [Google Scholar] [CrossRef]
- Ramirez, M.; Bauermann, F.V.; Navarro, D.; Rojas, M.; Manchego, A.; Nelson, E.A.; Diel, D.G.; Rivera, H. Detection of porcine reproductive and respiratory syndrome virus (PRRSV) 1-7-4-type strains in Peru. Transbound. Emerg. Dis. 2019, 66, 1107–1113. [Google Scholar] [CrossRef]
- Kim, S.C.; Jeong, C.G.; Park, G.S.; Park, J.Y.; Jeoung, H.Y.; Shin, G.E.; Ko, M.K.; Kim, S.H.; Lee, K.K.; Kim, W.I. Temporal lineage dynamics of the ORF5 gene of porcine reproductive and respiratory syndrome virus in Korea in 2014–2019. Arch. Virol. 2021, 166, 2803–2815. [Google Scholar] [CrossRef]
- Fukunaga, W.; Hayakawa-Sugaya, Y.; Koike, F.; Van Diep, N.; Kojima, I.; Yoshida, Y.; Suda, Y.; Masatani, T.; Ozawa, M. Newly-designed primer pairs for the detection of type 2 porcine reproductive and respiratory syndrome virus genes. J. Virol. Methods 2021, 291, 114071. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Li, W.; Zhao, J.; Gong, B.; Sun, Q.; Tang, Y.D.; Xiang, L.; Leng, C.; Peng, J.; et al. Novel characteristics of Chinese NADC34-like PRRSV during 2020–2021. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Trevisan, G.; Li, G.; Moura, C.A.A.; Coleman, K.; Thomas, P.; Zhang, J.; Gauger, P.; Zeller, M.; Linhares, D. Complete Coding Genome Sequence of a Novel Porcine Reproductive and Respiratory Syndrome Virus 2 Restriction Fragment Length Polymorphism 1-4-4 Lineage 1C Variant Identified in Iowa, USA. Microbiol. Resour. Announc. 2021, 10, e0044821. [Google Scholar] [CrossRef]
- Li, Y.; Xu, G.; Du, X.; Xu, L.; Ma, Z.; Li, Z.; Feng, Y.; Jiao, D.; Guo, W.; Xiao, S. Genomic characteristics and pathogenicity of a new recombinant strain of porcine reproductive and respiratory syndrome virus. Arch. Virol. 2021, 166, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Ye, M.; Li, S.; Huang, Y.; Zhou, R.; Yu, X.; Tian, K.; Zhu, J. Emergence of a novel highly pathogenic recombinant virus from three lineages of porcine reproductive and respiratory syndrome virus 2 in China 2017. Transbound. Emerg. Dis. 2018, 65, 1775–1785. [Google Scholar] [CrossRef]
- Zhang, H.; Leng, C.; Ding, Y.; Zhai, H.; Li, Z.; Xiang, L.; Zhang, W.; Liu, C.; Li, M.; Chen, J.; et al. Characterization of newly emerged NADC30-like strains of porcine reproductive and respiratory syndrome virus in China. Arch. Virol. 2019, 164, 401–411. [Google Scholar] [CrossRef]
- Brockmeier, S.L.; Loving, C.L.; Vorwald, A.C.; Kehrli, M.E., Jr.; Baker, R.B.; Nicholson, T.L.; Lager, K.M.; Miller, L.C.; Faaberg, K.S. Genomic sequence and virulence comparison of four Type 2 porcine reproductive and respiratory syndrome virus strains. Virus. Res. 2012, 169, 212–221. [Google Scholar] [CrossRef]
- Kwon, T.; Yoo, S.J.; Park, J.W.; Kang, S.C.; Park, C.K.; Lyoo, Y.S. Genomic characteristics and pathogenicity of natural recombinant porcine reproductive and respiratory syndrome virus 2 harboring genes of a Korean field strain and VR-2332-like strain. Virology 2019, 530, 89–98. [Google Scholar] [CrossRef]
- Jeong, J.; Kim, S.; Park, C.; Park, K.H.; Kang, I.; Park, S.J.; Chae, C. Commercial porcine reproductive and respiratory syndrome virus (PRRSV)-2 modified live virus vaccine against heterologous single and dual Korean PRRSV-1 and PRRSV-2 challenge. Vet. Rec. 2018, 182, 485. [Google Scholar] [CrossRef]
- Oh, T.; Kim, H.; Park, K.H.; Jeong, J.; Kang, I.; Yang, S.; Chae, C. Effectiveness of a commercial porcine reproductive and respiratory syndrome virus (PRRSV) subunit vaccine against heterologous PRRSV-1 and PRRSV-2 challenge in late-term pregnant gilts. Can. J. Vet. Res. 2019, 83, 248–254. [Google Scholar]
- Iseki, H.; Morozumi, T.; Takagi, M.; Kawashima, K.; Shibahara, T.; Uenishi, H.; Tsunemitsu, H. Genomic sequence and virulence evaluation of MN184A-like porcine reproductive and respiratory syndrome virus in Japan. Microbiol. Immunol. 2016, 60, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Ye, C.; Chang, X.B.; Jiang, C.G.; Wang, S.J.; Cai, X.H.; Tong, G.Z.; Tian, Z.J.; Shi, M.; An, T.Q. Importation and Recombination Are Responsible for the Latest Emergence of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus in China. J. Virol. 2015, 89, 10712–10716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.K.; Zhou, X.; Zhai, J.Q.; Li, B.; Wei, C.H.; Dai, A.L.; Yang, X.Y.; Luo, M.L. Emergence of a novel highly pathogenic porcine reproductive and respiratory syndrome virus in China. Transbound. Emerg. Dis. 2017, 64, 2059–2074. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhou, X.; Zhai, J.; Wei, C.; Dai, A.; Yang, X.; Luo, M. Recombination in JXA1-R vaccine and NADC30-like strain of porcine reproductive and respiratory syndrome viruses. Vet. Microbiol. 2017, 204, 110–120. [Google Scholar] [CrossRef]
- Han, G.; Lei, K.; Xu, H.; He, F. Genetic characterization of a novel recombinant PRRSV2 from lineage 8, 1 and 3 in China with significant variation in replication efficiency and cytopathic effects. Transbound. Emerg. Dis. 2020, 67, 1574–1584. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, J.; Bai, X.; Ji, G.; Yan, H.; Li, Y.; Wang, Y.; Tan, F.; Xiao, Y.; Li, X.; et al. Pathogenicity comparison between highly pathogenic and NADC30-like porcine reproductive and respiratory syndrome virus. Arch. Virol. 2016, 161, 2257–2261. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, B.; Xu, L.; Jin, H.; Ge, X.; Guo, X.; Han, J.; Yang, H. Efficacy evaluation of three modified-live virus vaccines against a strain of porcine reproductive and respiratory syndrome virus NADC30-like. Vet. Microbiol. 2017, 207, 108–116. [Google Scholar] [CrossRef]
- Wang, H.M.; Liu, Y.G.; Tang, Y.D.; Liu, T.X.; Zheng, L.L.; Wang, T.Y.; Liu, S.G.; Wang, G.; Cai, X.H. A natural recombinant PRRSV between HP-PRRSV JXA1-like and NADC30-like strains. Transbound. Emerg. Dis. 2018, 65, 1078–1086. [Google Scholar] [CrossRef]
- Bian, T.; Sun, Y.; Hao, M.; Zhou, L.; Ge, X.; Guo, X.; Han, J.; Yang, H. A recombinant type 2 porcine reproductive and respiratory syndrome virus between NADC30-like and a MLV-like: Genetic characterization and pathogenicity for piglets. Infect. Genet. Evol. 2017, 54, 279–286. [Google Scholar] [CrossRef]
- Zhou, L.; Kang, R.; Yu, J.; Xie, B.; Chen, C.; Li, X.; Xie, J.; Ye, Y.; Xiao, L.; Zhang, J.; et al. Genetic Characterization and Pathogenicity of a Novel Recombined Porcine Reproductive and Respiratory Syndrome Virus 2 among Nadc30-Like, Jxa1-Like, and Mlv-Like Strains. Viruses 2018, 10, 551. [Google Scholar] [CrossRef] [Green Version]
- Sui, X.; Guo, X.; Jia, H.; Wang, X.; Lin, W.; Li, M.; Gao, X.; Wu, J.; Jiang, Y.; Willems, L.; et al. Genomic sequence and virulence of a novel NADC30-like porcine reproductive and respiratory syndrome virus isolate from the Hebei province of China. Microb. Pathog. 2018, 125, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Kang, R.; Zhang, Y.; Yu, J.; Xie, B.; Chen, C.; Li, X.; Chen, B.; Liang, L.; Zhu, J.; et al. Emergence of two novel recombinant porcine reproductive and respiratory syndrome viruses 2 (lineage 3) in Southwestern China. Vet. Microbiol. 2019, 232, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xia, M.; Wang, W.; Ju, D.; Cao, L.; Wu, B.; Wang, X.; Wu, Y.; Song, N.; Hu, J.; et al. An Attenuated Highly Pathogenic Chinese PRRS Viral Vaccine Confers Cross Protection to Pigs against Challenge with the Emerging PRRSV NADC30-Like Strain. Virol. Sin. 2018, 33, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Dai, A.; Fan, J.; Li, Y.; Chen, A.; Zhou, X.; Luo, M.; Yang, X.; Liu, J. Efficacy of Type 2 PRRSV vaccine against challenge with the Chinese lineage 1 (NADC30-like) PRRSVs in pigs. Sci. Rep. 2019, 9, 10781. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Q.; Cao, Z.; Tang, Y.D.; Xia, D.; Wang, G.; Shan, H. Recent Advances in Porcine Reproductive and Respiratory Syndrome Virus NADC30-Like Research in China: Molecular Characterization, Pathogenicity, and Control. Front. Microbiol. 2021, 12, 791313. [Google Scholar] [CrossRef]
- Chae, C. Commercial PRRS Modified-Live Virus Vaccines. Vaccines 2021, 9, 185. [Google Scholar] [CrossRef]
- Li, C.; Liu, Z.; Chen, K.; Qian, J.; Hu, Y.; Fang, S.; Sun, Z.; Zhang, C.; Huang, L.; Zhang, J.; et al. Efficacy of the Synergy Between Live-Attenuated and Inactivated PRRSV Vaccines Against a NADC30-Like Strain of Porcine Reproductive and Respiratory Syndrome Virus in 4-Week Piglets. Front. Vet. Sci. 2022, 9, 812040. [Google Scholar] [CrossRef]
- Li, L.; Chen, J.; Cao, Z.; Cao, Y.; Guo, Z.; Tong, W.; Zhou, Y.; Li, G.; Jiang, Y.; Liu, C.; et al. Recombinant Bivalent Live Vectored Vaccine Against Classical Swine Fever and HP-PRRS Revealed Adequate Heterogeneous Protection Against NADC30-Like Strain. Front. Microbiol. 2021, 12, 822749. [Google Scholar] [CrossRef]
- Renukaradhya, G.J.; Meng, X.J.; Calvert, J.G.; Roof, M.; Lager, K.M. Inactivated and subunit vaccines against porcine reproductive and respiratory syndrome: Current status and future direction. Vaccine 2015, 33, 3065–3072. [Google Scholar] [CrossRef]
- Wang, H.; Xu, Y.; Feng, W. Porcine Reproductive and Respiratory Syndrome Virus: Immune Escape and Application of Reverse Genetics in Attenuated Live Vaccine Development. Vaccines 2021, 9, 480. [Google Scholar] [CrossRef]
- Bai, X.; Wang, Y.; Xu, X.; Sun, Z.; Xiao, Y.; Ji, G.; Li, Y.; Tan, F.; Li, X.; Tian, K. Commercial vaccines provide limited protection to NADC30-like PRRSV infection. Vaccine 2016, 34, 5540–5545. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.F.; Zhou, L.; Bian, T.; Tian, X.X.; Ren, W.K.; Lu, C.; Zhang, L.; Li, X.L.; Cui, M.S.; Yang, H.C.; et al. Efficacy evaluation of two commercial modified-live virus vaccines against a novel recombinant type 2 porcine reproductive and respiratory syndrome virus. Vet. Microbiol. 2018, 216, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jiang, P.; Song, Z.; Lv, L.; Li, L.; Bai, J. Pathogenicity and antigenicity of a novel NADC30-like strain of porcine reproductive and respiratory syndrome virus emerged in China. Vet. Microbiol. 2016, 197, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.J.; An, T.Q.; Zhou, Y.J.; Peng, J.M.; Hu, S.P.; Wei, T.C.; Jiang, Y.F.; Xiao, Y.; Tong, G.Z. An attenuated live vaccine based on highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) protects piglets against HP-PRRS. Vet. Microbiol. 2009, 138, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Tian, Z.; An, T.; Zhou, Y.; Xiao, Y.; Jiang, Y.; Hao, X.; Zhang, S.; Peng, J.; Qiu, H.; et al. Development and application of Taq Man-MGB fluorescence quantitative RT-PCR assay for detection of porcine reproductive and respiratory syndrome virus. Chin. J. Prev. Vet. Med. 2008, 30, 944–948. [Google Scholar]
- Zhou, L.; Wang, Z.; Ding, Y.; Ge, X.; Guo, X.; Yang, H. NADC30-like Strain of Porcine Reproductive and Respiratory Syndrome Virus, China. Emerg. Infect. Dis. 2015, 21, 2256–2257. [Google Scholar] [CrossRef]
- Guo, Z.; Chen, X.X.; Li, X.; Qiao, S.; Deng, R.; Zhang, G. Prevalence and genetic characteristics of porcine reproductive and respiratory syndrome virus in central China during 2016–2017: NADC30-like PRRSVs are predominant. Microb. Pathog. 2019, 135, 103657. [Google Scholar] [CrossRef]
- Chen, P.; Tan, X.; Lao, M.; Wu, X.; Zhao, X.; Zhou, S.; Yu, J.; Zhu, J.; Yu, L.; Tong, W.; et al. The Novel PRRSV Strain HBap4-2018 with a Unique Recombinant Pattern Is Highly Pathogenic to Piglets. Virol. Sin. 2021, 36, 1611–1625. [Google Scholar] [CrossRef]
- Yu, X.; Zhou, Z.; Cao, Z.; Wu, J.; Zhang, Z.; Xu, B.; Wang, C.; Hu, D.; Deng, X.; Han, W.; et al. Assessment of the safety and efficacy of an attenuated live vaccine based on highly pathogenic porcine reproductive and respiratory syndrome virus. Clin. Vaccine Immunol. 2015, 22, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Leng, X.; Li, Z.; Xia, M.; He, Y.; Wu, H. Evaluation of the efficacy of an attenuated live vaccine against highly pathogenic porcine reproductive and respiratory syndrome virus in young pigs. Clin. Vaccine Immunol. 2012, 19, 1199–1206. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, W.; Gong, W.; Zhang, D.; She, R.; Xu, B.; Ning, Y. Comparative Respiratory Pathogenicity and Dynamic Tissue Distribution of Chinese Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus and its Attenuated Strain in Piglets. J. Comp. Pathol. 2015, 153, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Song, T.; Yu, Y.; Liu, Y.; Shi, W.; Wang, S.; Rong, F.; Dong, J.; Liu, H.; Cai, X.; et al. Immune responses in piglets infected with highly pathogenic porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2011, 142, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Leng, C.; Zhang, W.; Zhang, H.; Kan, Y.; Yao, L.; Zhai, H.; Li, M.; Li, Z.; Liu, C.; An, T.; et al. ORF1a of highly pathogenic PRRS attenuated vaccine virus plays a key role in neutralizing antibody induction in piglets and virus neutralization in vitro. Virol. J. 2017, 14, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, W.; Liu, Z.; Sun, Z.; Su, L.; Zhang, C.; Huang, L. Efficacy of two porcine reproductive and respiratory syndrome (PRRS) modified-live virus (MLV) vaccines against heterologous NADC30-like PRRS virus challenge. Vet. Microbiol. 2020, 248, 108805. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Vaccination | Number of Animals |
|---|---|---|
| High-dose group of 3B1 | 2 × 106.0TCID50 | 5 (100; 101;102; 103; 104) |
| High-dose group of 3B2 | 2 × 106.5TCID50 | 5 (105; 106; 107; 108; 109) |
| High-dose group of 3B3 | 2 × 106.2TCID50 | 5 (110; 111; 112; 113; 114) |
| Repeated-dose group of 3B1 | 2 × 105.0TCID50 +2 × 105.0TCID50 | 5 (70; 71; 73; 74; 75) |
| Repeated-dose group of 3B2 | 2 × 105.5TCID50 +2 × 105.5TCID50 | 5 (77; 78; 79; 80; 81) |
| Repeated-dose group of 3B3 | 2 × 105.2TCID50 +2 × 105.2TCID50 | 5 (92; 93; 94; 95; 96) |
| Groups | Corresponding Groups | Number of Animals | Vaccination | Challenge |
|---|---|---|---|---|
| Group A1 | SD-R vaccine-treated and SD-challenge group | 5 (031; 032;035; 037; 038) | 2 × 105.0TCID50 per pig (SD-R) | 4 × 105.0TCID50 per pig (SD) |
| Group B1 | SD-challenge group | 5 (081; 082; 083; 084; 085) | DMEM | |
| Group C1 | Negative control group | 3 (033; 034; 036) | DMEM | |
| Group A2 | SD-R vaccine-treated and HLJWK108-1711-challenge group | 5 (073; 074; 075; 077; 079) | 2 × 105.0TCID50 per pig (SD-R) | 4 × 105.0TCID50 per pig (HLJWK108-1711) |
| Group B2 | HLJWK108-1711-challenge group | 5 (057; 058; 059; 078; 080) | DMEM | |
| Group C2 | Negative control group | 3 (076; 088; 090) | DMEM |
| Gene | Nucleotide Similarity (%) | Deduced Amino Acid Similarity (%) | Gene | Nucleotide Similarity (%) | Deduced Amino Acid Similarity (%) |
|---|---|---|---|---|---|
| Whole genome | 89.9 | / | Nsp8 | 94.8 | 93.3 |
| skeleton section of NADC30 | 91.4 | / | Nsp9 | 89.2 | 97.2 |
| 5′UTR | 91.5 | / | Nsp10 | 94.0 | 97.3 |
| 3′UTR | 97.3 | / | Nsp11 | 93.1 | 95.1 |
| Nsp1α | 88.5 | 93.3 | Nsp12 | 93.7 | 96.1 |
| Nsp1β | 82.7 | 80.2 | ORF2a | 94.0 | 92.6 |
| Nsp2 | 87.0 | 84.9 | ORF2b | 97.7 | 100 |
| Nsp3 | 90.4 | 93.0 | ORF3 | 86.8 | 84.3 |
| Nsp4 | 82.8 | 92.6 | ORF4 | 94.6 | 96.1 |
| Nsp5 | 84.1 | 91.8 | ORF5 | 93.5 | 93.0 |
| Nsp6 | 93.8 | 100 | ORF5a | 96.4 | 95.7 |
| Nsp7α | 91.1 | 90.6 | ORF6 | 95.6 | 98.9 |
| Nsp7β | 90.9 | 90.9 | ORF7 | 94.9 | 93.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Xiang, L.; Xu, H.; Li, C.; Tang, Y.-D.; Gong, B.; Zhang, W.; Zhao, J.; Song, S.; Peng, J.; et al. Lineage 1 Porcine Reproductive and Respiratory Syndrome Virus Attenuated Live Vaccine Provides Broad Cross-Protection against Homologous and Heterologous NADC30-Like Virus Challenge in Piglets. Vaccines 2022, 10, 752. https://doi.org/10.3390/vaccines10050752
Zhang H, Xiang L, Xu H, Li C, Tang Y-D, Gong B, Zhang W, Zhao J, Song S, Peng J, et al. Lineage 1 Porcine Reproductive and Respiratory Syndrome Virus Attenuated Live Vaccine Provides Broad Cross-Protection against Homologous and Heterologous NADC30-Like Virus Challenge in Piglets. Vaccines. 2022; 10(5):752. https://doi.org/10.3390/vaccines10050752
Chicago/Turabian StyleZhang, Hongliang, Lirun Xiang, Hu Xu, Chao Li, Yan-Dong Tang, Bangjun Gong, Wenli Zhang, Jing Zhao, Shuaijie Song, Jinmei Peng, and et al. 2022. "Lineage 1 Porcine Reproductive and Respiratory Syndrome Virus Attenuated Live Vaccine Provides Broad Cross-Protection against Homologous and Heterologous NADC30-Like Virus Challenge in Piglets" Vaccines 10, no. 5: 752. https://doi.org/10.3390/vaccines10050752