
Molecular Characteristics and Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) 1 in Taiwan during 2019–2020

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Amplification of the ORF5 Gene and Phylogenetic Analysis
2.3. Identification of Selected Sites of the ORF5 Gene
2.4. Animal Experiment
2.5. Statistics
3. Results
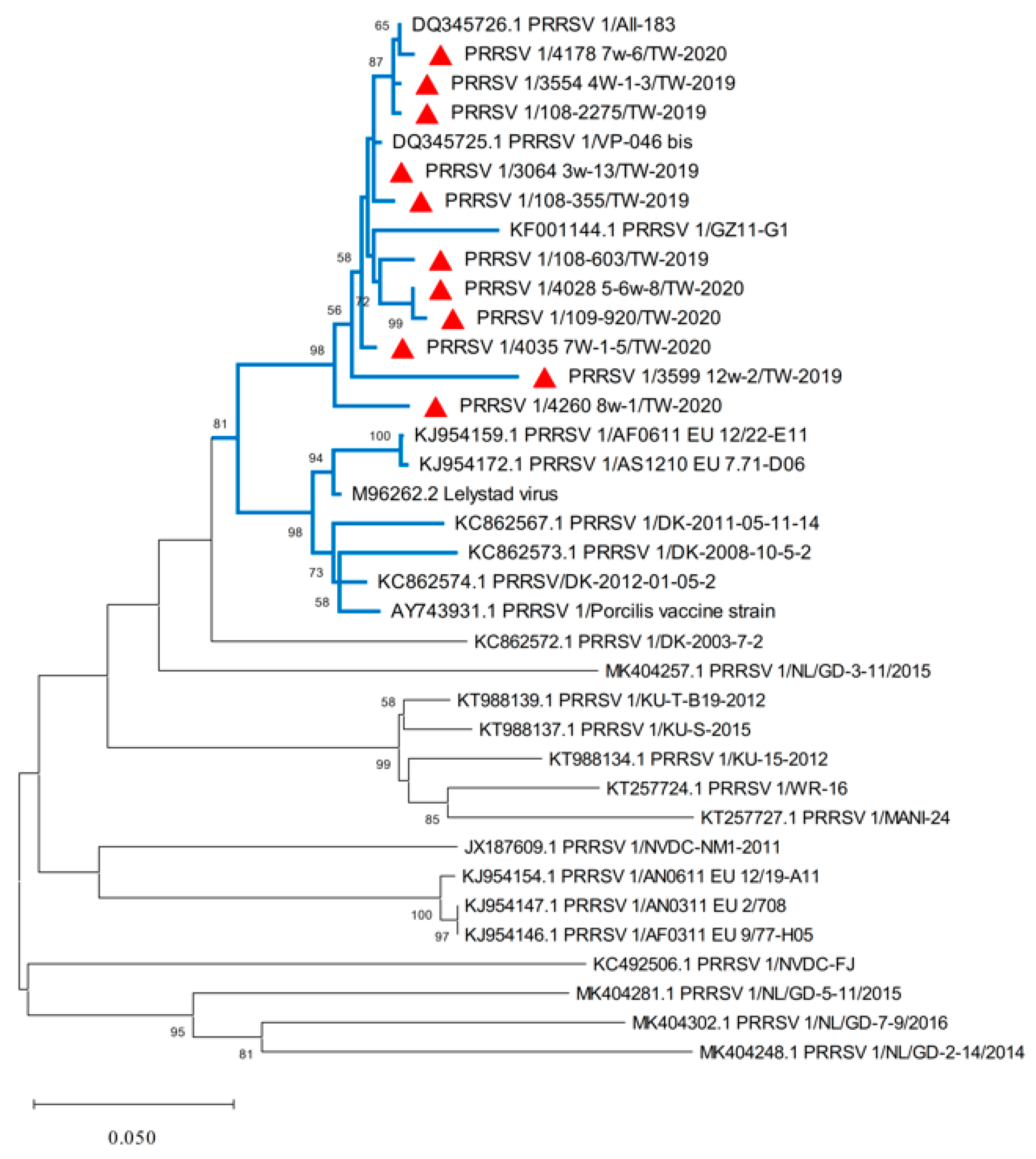
3.1. PRRSV 1 Nucleotide Sequencing and Phylogenetic Analysis of the ORF5 Gene
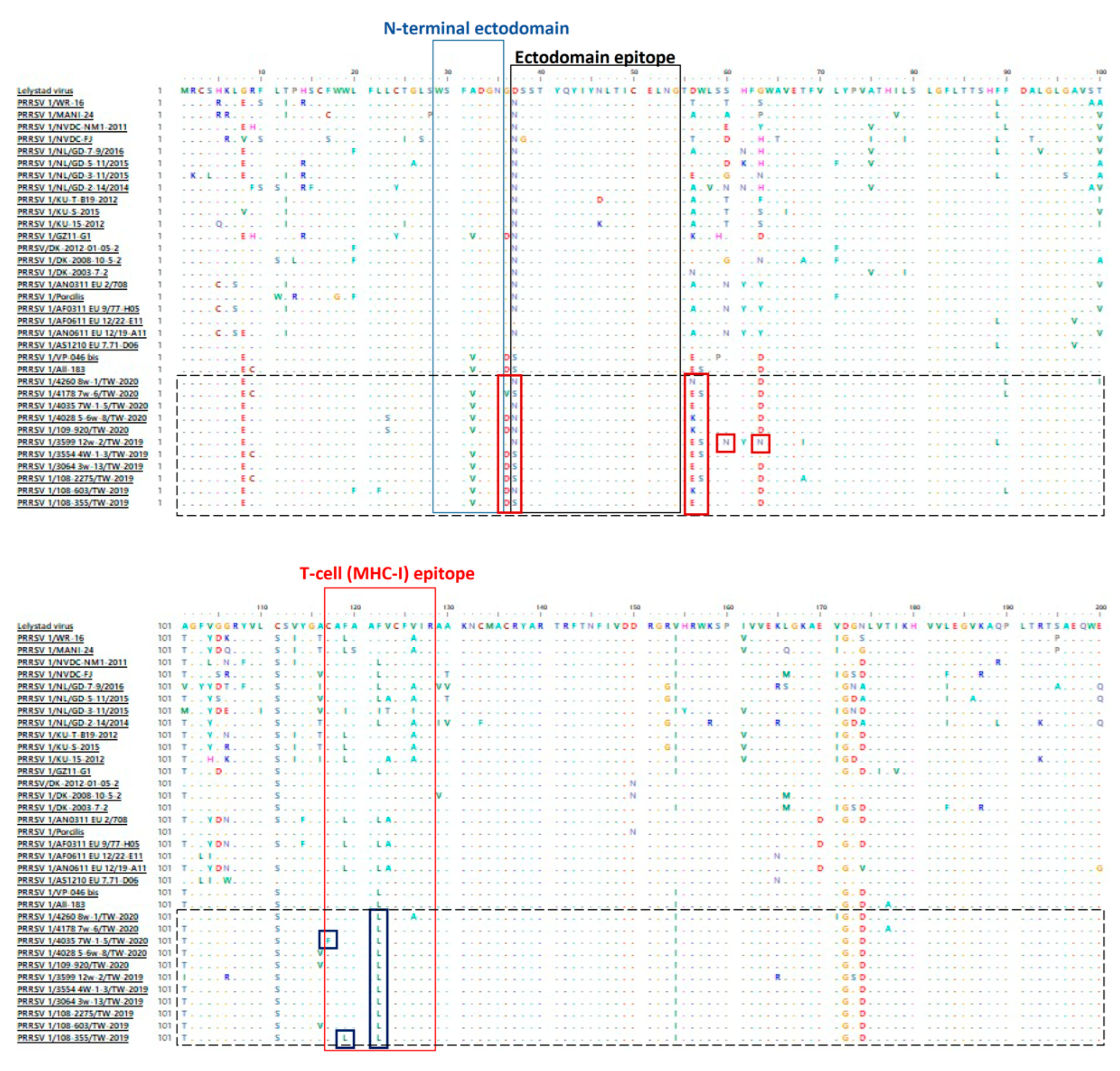
3.2. Amino Acid Analysis of ORF5 of PRRSV 1
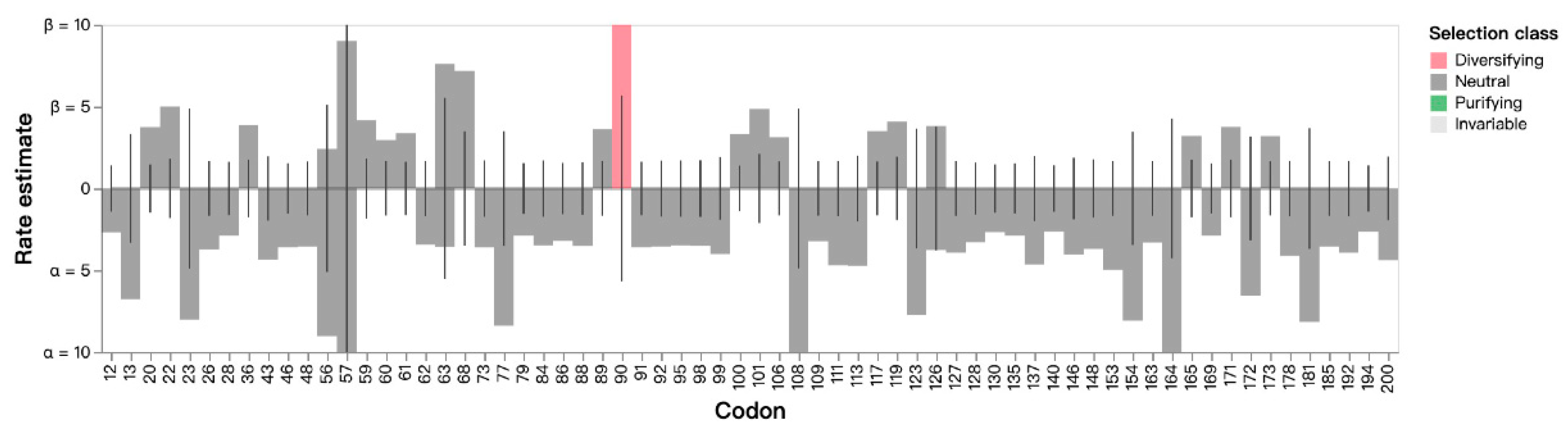
3.3. Estimation of Non-Synonymous and Synonymous Substitutions of the ORF5 Gene
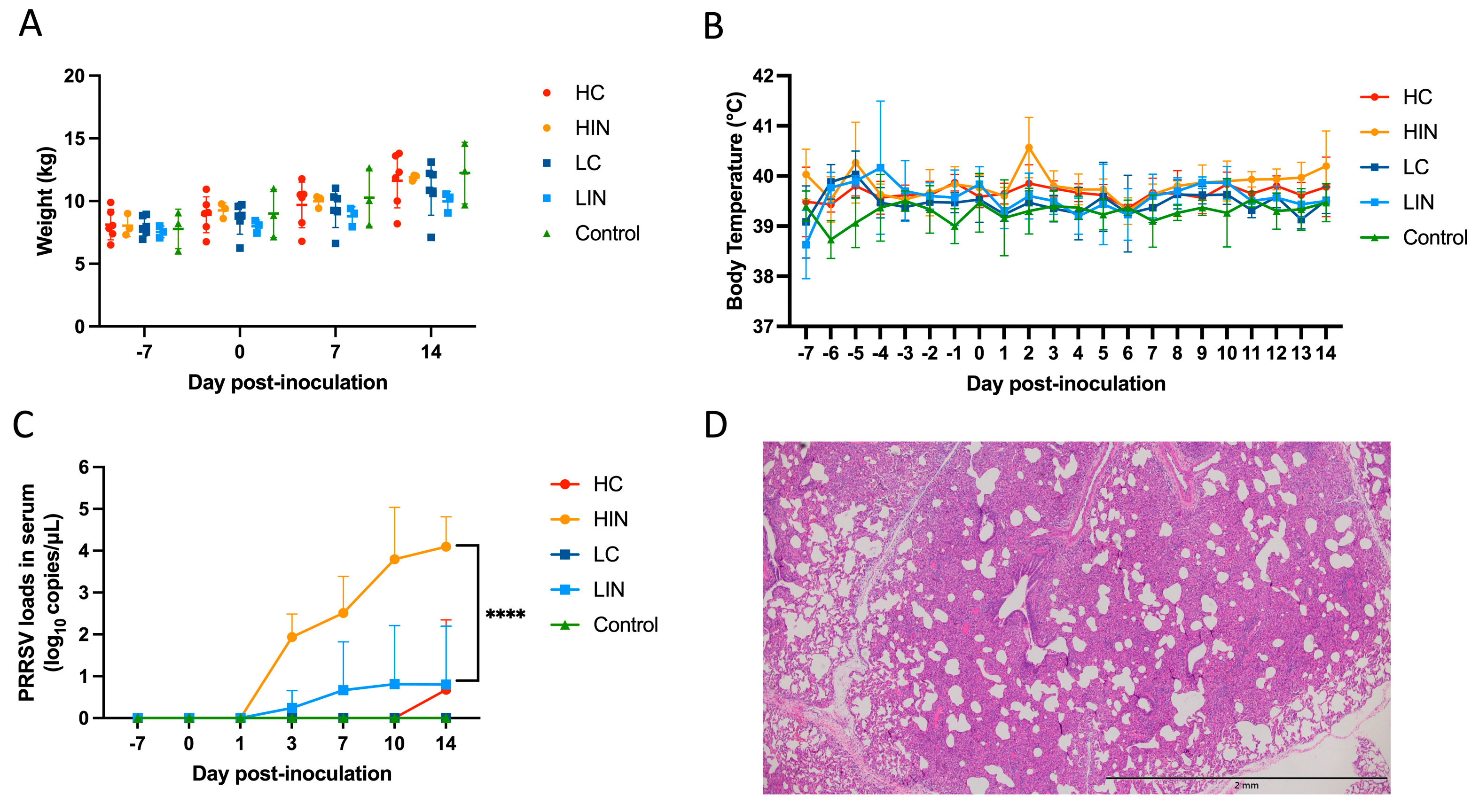
3.4. Pathogenicity of Virulent NPUST2789 Challenge in Nursery Pigs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F. Mystery Swine Disease in The Netherlands: The Isolation of Lelystad Virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; Mccullough, S.; Morrison, R.B.; Joo, H.S.; et al. Isolation of Swine Infertility and Respiratory Syndrome Virus (Isolate ATCC VR-2332) in North America and Experimental Reproduction of the Disease in Gnotobiotic Pigs. J. Vet. Diagn. Invest. 1992, 4, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Rossow, K.D. Porcine Reproductive and Respiratory Syndrome. Vet. Pathol. 1998, 35, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Darwich, L.; Gimeno, M.; Sibila, M.; Diaz, I.; de la Torre, E.; Dotti, S.; Kuzemtseva, L.; Martin, M.; Pujols, J.; Mateu, E. Genetic and Immunobiological Diversities of Porcine Reproductive and Respiratory Syndrome Genotype I Strains. Vet. Microbiol. 2011, 150, 49–62. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to Taxonomy and the International Code of Virus Classification and Nomenclature Ratified by the International Committee on Taxonomy of Viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Kang, I.; Jeong, J.; Oh, T.; Park, K.H.; Park, S.-J.; Ham, H.J.; Jin, G.-R.; Lee, B.-H.; Chae, C. A Comparison of Two Commercially Available Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Modified-Live Virus Vaccines Analyzing the Growth Performance in 1-Day-Old Vaccinated Swine Located on Endemic Farms Co-Circulating PRRSV-1 and PRRSV-2. J. Vet. Med. Sci. 2020, 82, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Chung, W.B.; Lin, W.M.; Yang, P.C.; Weng, C.N.; Chiu, Y.T.; Chang, W.F.; Chu, R.M. Porcine Reproductive and Respiratory Syndrome (PRRS) in Taiwan. I: Viral Isolation. J. Chin. Soc. Vet. Sci. 1993, 19, 268–276. [Google Scholar]
- Lin, W.H.; Kaewprom, K.; Wang, S.Y.; Lin, C.F.; Yang, C.Y.; Chiou, M.T.; Lin, C.N. Outbreak of Porcine Reproductive and Respiratory Syndrome Virus 1 in Taiwan. Viruses 2020, 12, 316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bào, Y.; Ng, T.F.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and Expansion of the Nidoviral Family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Mardassi, H.; Mounir, S.; Dea, S. Molecular Analysis of the ORFs 3 to 7 of Porcine Reproductive and Respiratory Syndrome Virus, Quebec Reference Strain*. Arch. Virol. 1995, 140, 1405–1418. [Google Scholar] [CrossRef]
- Kim, W.-I.; Kim, J.-J.; Cha, S.-H.; Wu, W.-H.; Cooper, V.; Evans, R.; Choi, E.-J.; Yoon, K.-J. Significance of Genetic Variation of PRRSV ORF5 in Virus Neutralization and Molecular Determinants Corresponding to Cross Neutralization among PRRS Viruses. Vet. Microbiol. 2013, 162, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Wissink, E.H.J.; van Wijk, H.A.R.; Kroese, M.V.; Weiland, E.; Meulenberg, J.J.M.; Rottier, P.J.M.; van Rijn, P.A. The Major Envelope Protein, GP5, of a European Porcine Reproductive and Respiratory Syndrome Virus Contains a Neutralization Epitope in Its N-Terminal Ectodomain. J. Gen. Virol. 2003, 84, 1535–1543. [Google Scholar] [CrossRef]
- Plagemann, P.G.W. GP5 Ectodomain Epitope of Porcine Reproductive and Respiratory Syndrome Virus, Strain Lelystad Virus. Virus Res. 2004, 102, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Díaz, I.; Pujols, J.; Ganges, L.; Gimeno, M.; Darwich, L.; Domingo, M.; Mateu, E. In Silico Prediction and Ex Vivo Evaluation of Potential T-Cell Epitopes in Glycoproteins 4 and 5 and Nucleocapsid Protein of Genotype-I (European) of Porcine Reproductive and Respiratory Syndrome Virus. Vaccine 2009, 27, 5603–5611. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.L.X.; Kwon, B.; Yoon, K.-J.; Laegreid, W.W.; Pattnaik, A.K.; Osorio, F.A. Immune Evasion of Porcine Reproductive and Respiratory Syndrome Virus through Glycan Shielding Involves Both Glycoprotein 5 as Well as Glycoprotein 3. J. Virol. 2011, 85, 5555–5564. [Google Scholar] [CrossRef] [Green Version]
- Ansari, I.H.; Kwon, B.; Osorio, F.A.; Pattnaik, A.K. Influence of N-Linked Glycosylation of Porcine Reproductive and Respiratory Syndrome Virus GP5 on Virus Infectivity, Antigenicity, and Ability To Induce Neutralizing Antibodies. J. Virol. 2006, 80, 3994–4004. [Google Scholar] [CrossRef] [Green Version]
- Wissink, E.H.J.; Kroese, M.V.; Maneschijn-Bonsing, J.G.; Meulenberg, J.J.M.; van Rijn, P.A.; Rijsewijk, F.A.M.; Rottier, P.J.M. Significance of the Oligosaccharides of the Porcine Reproductive and Respiratory Syndrome Virus Glycoproteins GP2a and GP5 for Infectious Virus Production. J. Gen. Virol. 2004, 85, 3715–3723. [Google Scholar] [CrossRef]
- Wei, Z.; Lin, T.; Sun, L.; Li, Y.; Wang, X.; Gao, F.; Liu, R.; Chen, C.; Tong, G.; Yuan, S. N-Linked Glycosylation of GP5 of Porcine Reproductive and Respiratory Syndrome Virus Is Critically Important for Virus Replication In Vivo. J. Virol. 2012, 86, 9941–9951. [Google Scholar] [CrossRef] [Green Version]
- Stadejek, T.; Oleksiewicz, M.B.; Potapchuk, D.; Podgórska, K. Porcine Reproductive and Respiratory Syndrome Virus Strains of Exceptional Diversity in Eastern Europe Support the Definition of New Genetic Subtypes. J. Gen. Virol. 2006, 87, 1835–1841. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.Y.; Hon, C.C.; Hui, R.K.H.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C.C. Molecular Epidemiology of PRRSV: A Phylogenetic Perspective. Virus Res. 2010, 154, 7–17. [Google Scholar] [CrossRef]
- Balka, G.; Podgórska, K.; Brar, M.S.; Bálint, Á.; Cadar, D.; Celer, V.; Dénes, L.; Dirbakova, Z.; Jedryczko, A.; Márton, L.; et al. Genetic Diversity of PRRSV 1 in Central Eastern Europe in 1994-2014: Origin and Evolution of the Virus in the Region. Sci. Rep. 2018, 8, 7811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimman, T.G.; Cornelissen, L.A.; Moormann, R.J.; Rebel, J.M.J.; Stockhofe-Zurwieden, N. Challenges for Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) Vaccinology. Vaccine 2009, 27, 3704–3718. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ge, X.; Yang, H. Porcine Reproductive and Respiratory Syndrome Modified Live Virus Vaccine: A “Leaky” Vaccine with Debatable Efficacy and Safety. Vaccines 2021, 9, 362. [Google Scholar] [CrossRef] [PubMed]
- Kosakovsky Pond, S.L.; Frost, S.D.W. Not So Different After All: A Comparison of Methods for Detecting Amino Acid Sites Under Selection. Mol. Biol. Evol. 2005, 22, 1208–1222. [Google Scholar] [CrossRef] [Green Version]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A Fast, Unconstrained Bayesian AppRoximation for Inferring Selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.; Shank, S.D.; Spielman, S.J.; Li, M.; Muse, S.V.; Kosakovsky Pond, S.L. Datamonkey 2.0: A Modern Web Application for Characterizing Selective and Other Evolutionary Processes. Mol. Biol. Evol. 2018, 35, 773–777. [Google Scholar] [CrossRef] [Green Version]
- Prieto, C.; Vázquez, A.; Núñez, J.I.; Álvarez, E.; Simarro, I.; Castro, J.M. Influence of Time on the Genetic Heterogeneity of Spanish Porcine Reproductive and Respiratory Syndrome Virus Isolates. Vet. J. 2009, 180, 363–370. [Google Scholar] [CrossRef]
- Nguyen, V.G.; Kim, H.K.; Moon, H.J.; Park, S.J.; Chung, H.C.; Choi, M.K.; Park, B.K. A Bayesian Phylogeographical Analysis of Type 1 Porcine Reproductive and Respiratory Syndrome Virus (PRRSV). Transbound Emerg. Dis. 2014, 61, 537–545. [Google Scholar] [CrossRef]
- Sun, Q.; Xu, H.; Li, C.; Gong, B.; Li, Z.; Tian, Z.-J.; Zhang, H. Emergence of a Novel PRRSV-1 Strain in Mainland China: A Recombinant Strain Derived from the Two Commercial Modified Live Viruses Amervac and DV. Front. Vet. Sci. 2022, 9, 974743. [Google Scholar] [CrossRef]
- Smith, N.; Power, U.F.; McKillen, J. Phylogenetic Analysis of Porcine Reproductive and Respiratory Syndrome Virus Isolates from Northern Ireland. Arch. Virol. 2018, 163, 2799–2804. [Google Scholar] [CrossRef] [Green Version]
- Kvisgaard, L.K.; Hjulsager, C.K.; Kristensen, C.S.; Lauritsen, K.T.; Larsen, L.E. Genetic and Antigenic Characterization of Complete Genomes of Type 1 Porcine Reproductive and Respiratory Syndrome Viruses (PRRSV) Isolated in Denmark over a Period of 10 Years. Virus Res. 2013, 178, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Kim, H.; Kang, B.; Yeom, M.; Han, S.; Moon, H.; Park, S.; Kim, H.; Song, D.; Park, B. Prevalence and Phylogenetic Analysis of the Isolated Type I Porcine Reproductive and Respiratory Syndrome Virus from 2007 to 2008 in Korea. Virus Genes 2010, 40, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Amonsin, A.; Tatsanakit, A.; Damrongwatanapokin, S. Genetics and Geographical Variation of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) in Thailand. Vet. Microbiol. 2004, 101, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Yu, J.E.; Shin, J.-E.; Kang, A.; Kim, W.-I.; Lee, C.; Lee, J.; Cho, I.-S.; Choe, S.-E.; Cha, S.-H. Geographic Distribution and Molecular Analysis of Porcine Reproductive and Respiratory Syndrome Viruses Circulating in Swine Farms in the Republic of Korea between 2013 and 2016. BMC Vet. Res. 2018, 14, 160. [Google Scholar] [CrossRef] [PubMed]
- Zhai, S.-L.; Lin, T.; Zhou, X.; Pei, Z.-F.; Wei, Z.-Z.; Zhang, H.; Wen, X.-H.; Chen, Q.-L.; Lv, D.-H.; Wei, W.-K. Phylogeographic Analysis of Porcine Reproductive and Respiratory Syndrome Virus 1 in Guangdong Province, Southern China. Arch. Virol. 2018, 163, 2443–2449. [Google Scholar] [CrossRef]
- Dortmans, J.C.F.M.; Buter, G.J.; Dijkman, R.; Houben, M.; Duinhof, T.F. Molecular Characterization of Type 1 Porcine Reproductive and Respiratory Syndrome Viruses (PRRSV) Isolated in the Netherlands from 2014 to 2016. PLoS ONE 2019, 14, e0218481. [Google Scholar] [CrossRef]
- Martín-Valls, G.E.; Cortey, M.; Allepuz, A.; Illas, F.; Tello, M.; Mateu, E. Introduction of a PRRSV-1 Strain of Increased Virulence in a Pig Production Structure in Spain: Virus Evolution and Impact on Production. Porc. Health Manag. 2023, 9, 1. [Google Scholar] [CrossRef]
- Nan, Y.; Wu, C.; Gu, G.; Sun, W.; Zhang, Y.-J.; Zhou, E.-M. Improved Vaccine against PRRSV: Current Progress and Future Perspective. Front. Microbiol. 2017, 8, 1635. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Farm | Collection Time | Location | Scale of Herd | Type of Vaccine 1 | Age in Weeks | Animal Condition 2 | Quantitation of PRRSV 1/PRRSV 2 Loads 3 | Sample |
|---|---|---|---|---|---|---|---|---|
| Farm A | January 2019 | Pingtung | 1400 sows | Live attenuated PRRSV 1 | 3 | Sick | 1.10 × 106/− 4 | NPUST3064 |
| Farm B | March 2019 | Pingtung | 170 sows | Live attenuated PRRSV 1 | 8 | Sick | 5.85 × 106/− | NPUST108-355 |
| Farm C | April 2019 | Kaohsiung | 200 sows | Live attenuated PRRSV 1 | 6–7 | Sick | 1.87 × 106/− | NPUST108-603 |
| Farm D | August 2019 | Kaohsiung | 140 sows | Live attenuated PRRSV 1 and Modified live PRRSV 2 | 4 | Normal | 1.90 × 104/− | NPUST3554 |
| Farm E | August 2019 | Pingtung | 480 sows | Modified live PRRSV 2 | 12 | Normal | 4.40 × 104/− | NPUST3599 |
| Farm F | November 2019 | Miaoli | 400 sows | Live attenuated PRRSV 1 | 5 | Sick | 1.77 × 106/− | NPUST108-2275 |
| Farm G | February 2020 | Yunlin | 900 sows | Live attenuated PRRSV 1 | 5–6 | Sick | 2.26 × 105/− | NPUST4028 |
| Farm H | February 2020 | Pingtung | 250 sows | Live attenuated PRRSV 1 | 7 | Normal | 7.1 × 104/5.2 × 103 | NPUST4035 |
| Farm I | March 2020 | Yunlin | 200 sows | Live attenuated PRRSV 1 | 7 | Normal | 1.7 × 104/− | NPUST4178 |
| Farm J | March 2020 | Hsinchu | 200 sows | Non provided | 8 | Normal | 4.7 × 104/− | NPUST4260 |
| Farm K | April 2020 | Yunlin | 1100 hogs | Modified live PRRSV 2 | 5 | Sick | 6.64 × 107/− | NPUST109-920 |
| Name of Primers | Sequences |
|---|---|
| EU-ORF5-F1 | 5′-TGCATTTCYTGACACCATC-3′ |
| EU-ORF5-R1 | 5′-CCCBARRAGTCGGCCRCGWGA-3′ |
| EU-ORF5-F2 | 5′-ATCYRCAATGAGGTGGGCTAC-3′ |
| EU-ORF5-R2 | 5′-ACYTTNAGGGCRTADATCAT-3′ |
| Year | Number | Positive (%) | PRRSV 1 Only (%) | PRRSV 2 Only (%) | PRRSV 1 + PRRSV 2 (%) |
|---|---|---|---|---|---|
| 2019 | 8852 | 2864 (32.4) | 108 (3.8) | 2747 (95.9) | 9 (0.3) |
| 2020 | 6182 | 2600 (42.1) | 50 (1.9) | 2528 (97.2) | 22 (0.9) |
| Total | 15,034 | 5464 (36.3) | 158 (2.9) | 5275 (96.5) | 31 (0.6) |
| Codon Site | Mean Posterior Beta-Alpha | Negative Selection | Positive Selection | |
|---|---|---|---|---|
| Prob [Alpha > Beta] a | Prob [Alpha < Beta] b | Bayes Factor [Alpha < Beta] c | ||
| 77 | −12.955 | 0.974 | 0.017 | 0.023 |
| 90 | 9.578 | 0.018 | 0.961 | 31.945 |
| 108 | −17.870 | 0.984 | 0.009 | 0.012 |
| 123 | −14.816 | 0.979 | 0.014 | 0.018 |
| 164 | −20.986 | 0.988 | 0.007 | 0.010 |
| 181 | −13.273 | 0.972 | 0.018 | 0.024 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsueh, F.-C.; Kuo, K.-L.; Hsu, F.-Y.; Wang, S.-Y.; Chiu, H.-J.; Wu, M.-T.; Lin, C.-F.; Huang, Y.-H.; Chiou, M.-T.; Lin, C.-N. Molecular Characteristics and Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) 1 in Taiwan during 2019–2020. Life 2023, 13, 843. https://doi.org/10.3390/life13030843
Hsueh F-C, Kuo K-L, Hsu F-Y, Wang S-Y, Chiu H-J, Wu M-T, Lin C-F, Huang Y-H, Chiou M-T, Lin C-N. Molecular Characteristics and Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) 1 in Taiwan during 2019–2020. Life. 2023; 13(3):843. https://doi.org/10.3390/life13030843
Chicago/Turabian StyleHsueh, Fu-Chun, Kun-Lin Kuo, Feng-Yang Hsu, Sheng-Yuan Wang, Hsien-Jen Chiu, Meng-Tien Wu, Chuen-Fu Lin, Yu-Han Huang, Ming-Tang Chiou, and Chao-Nan Lin. 2023. "Molecular Characteristics and Pathogenicity of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) 1 in Taiwan during 2019–2020" Life 13, no. 3: 843. https://doi.org/10.3390/life13030843