
Changing Dynamics of SARS-CoV-2: A Global Challenge

 , , ,
, , ,  ,
,
Abstract
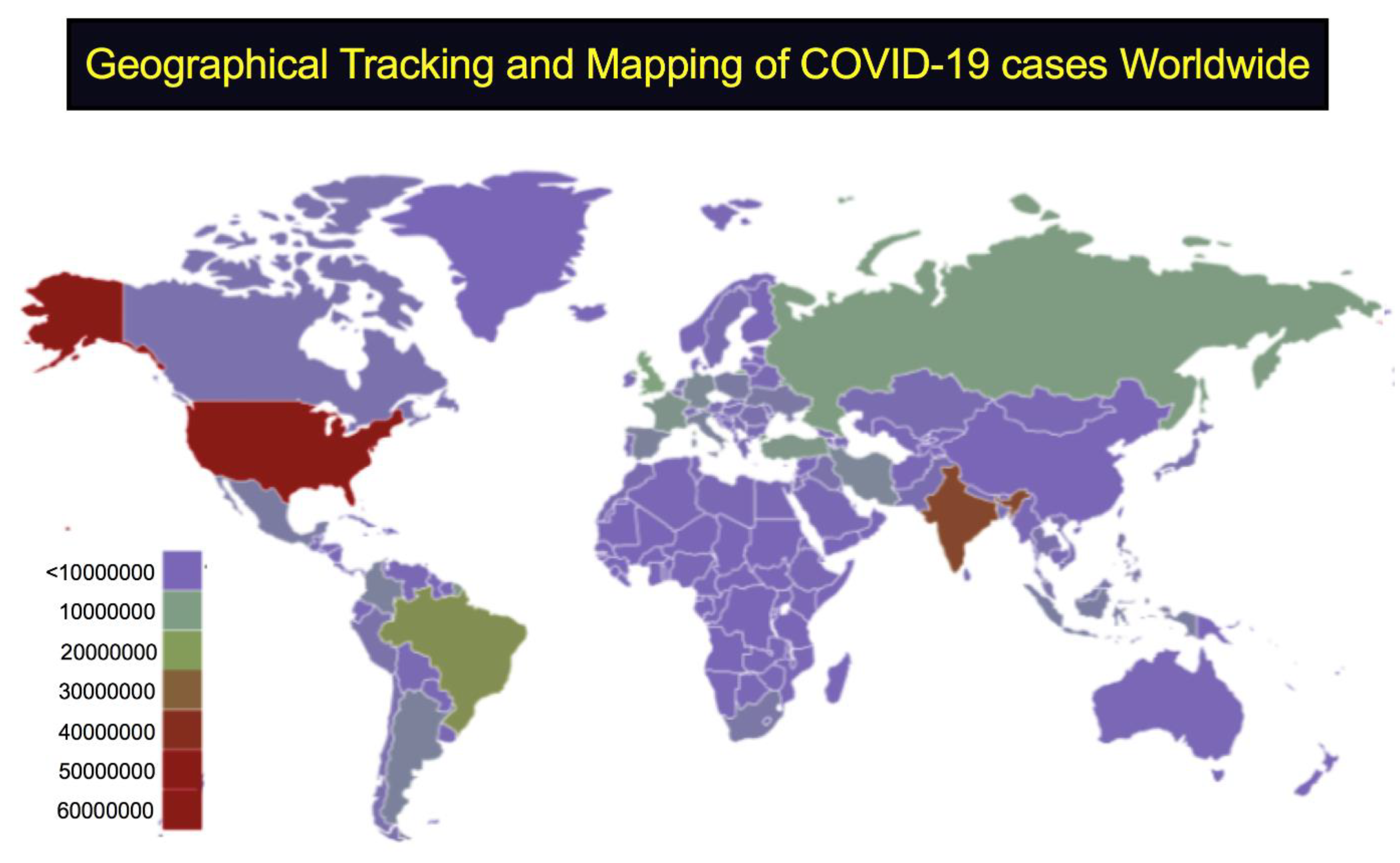
:1. Introduction
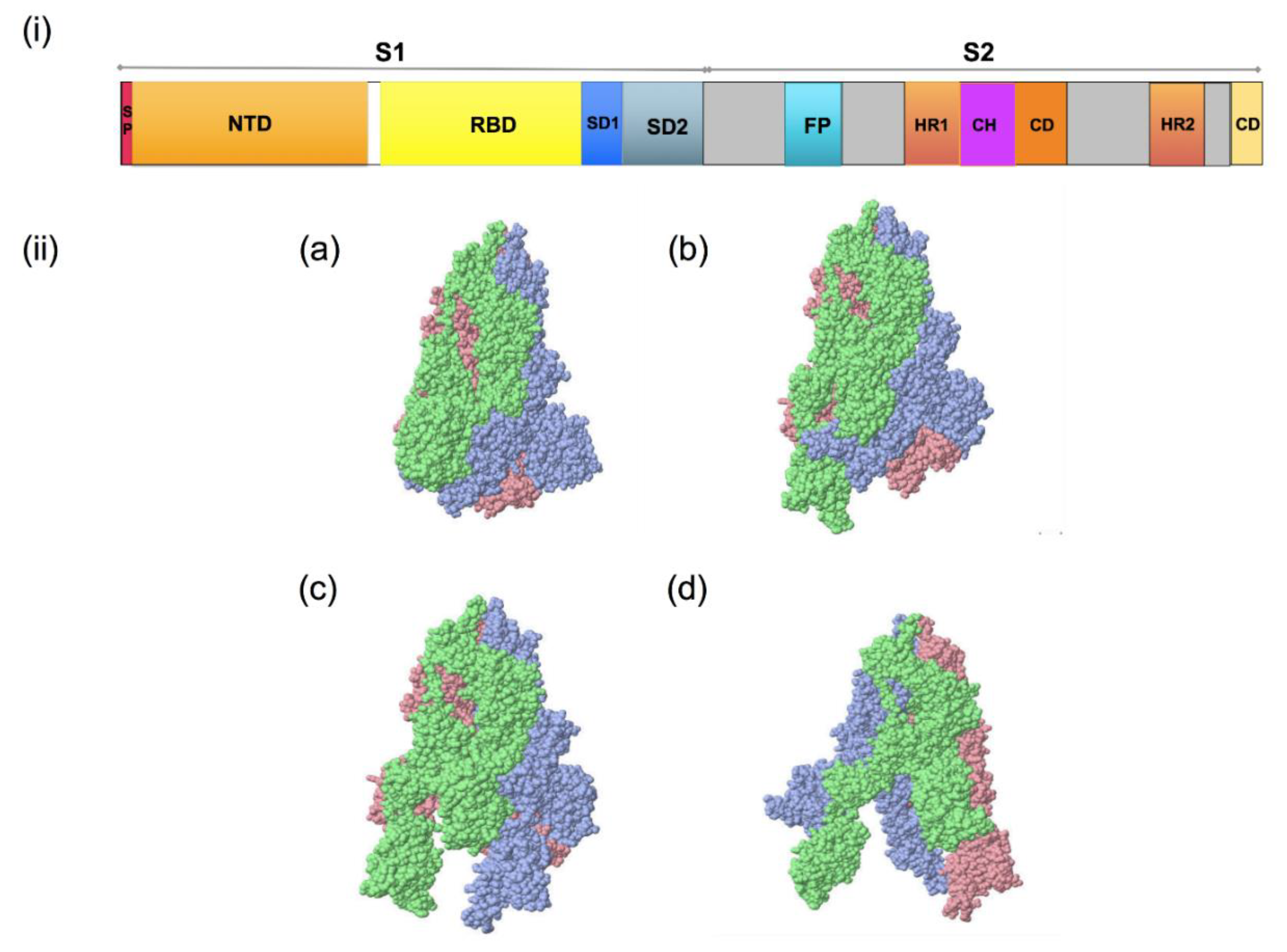
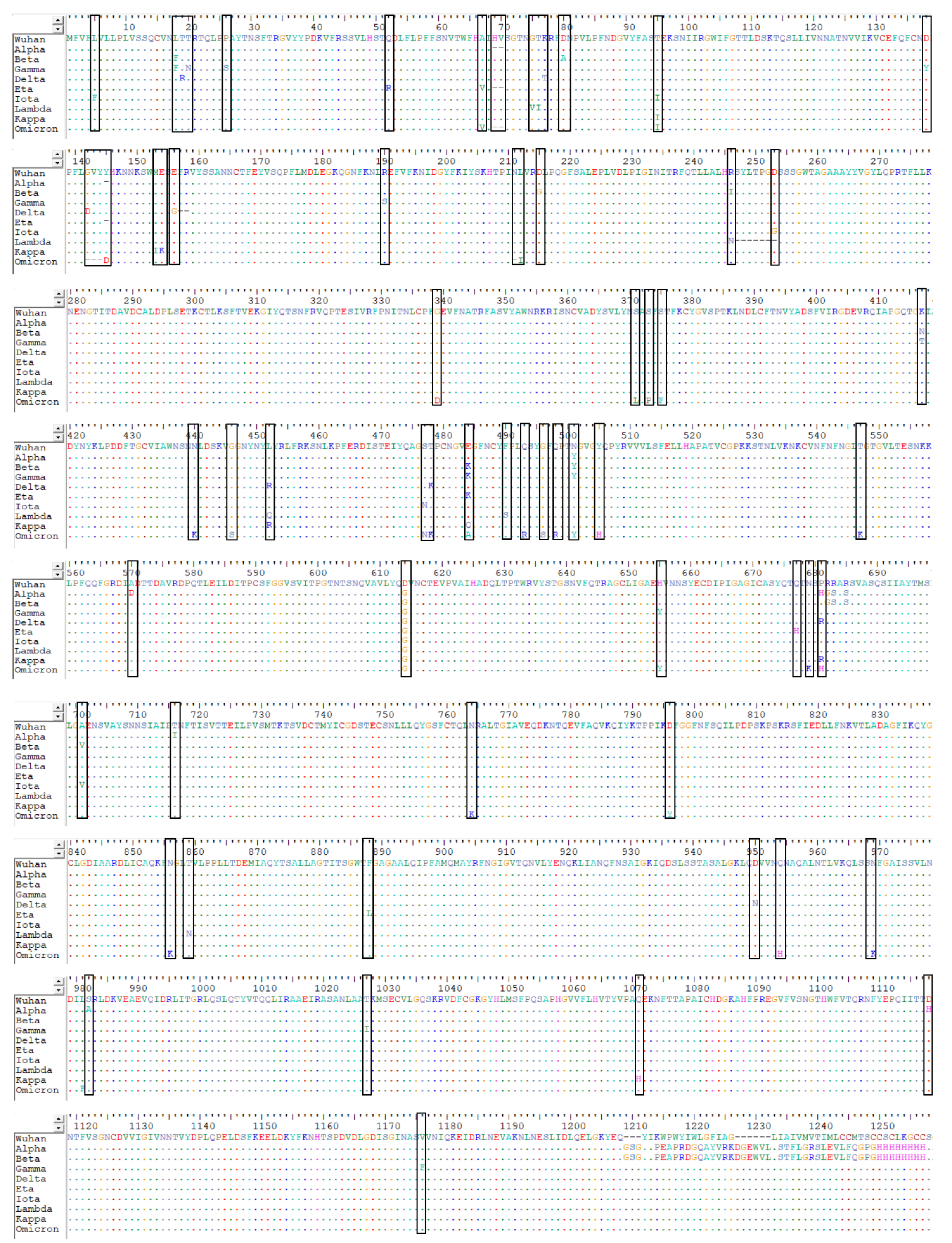
2. Structure and Sequence of SARS-CoV-2
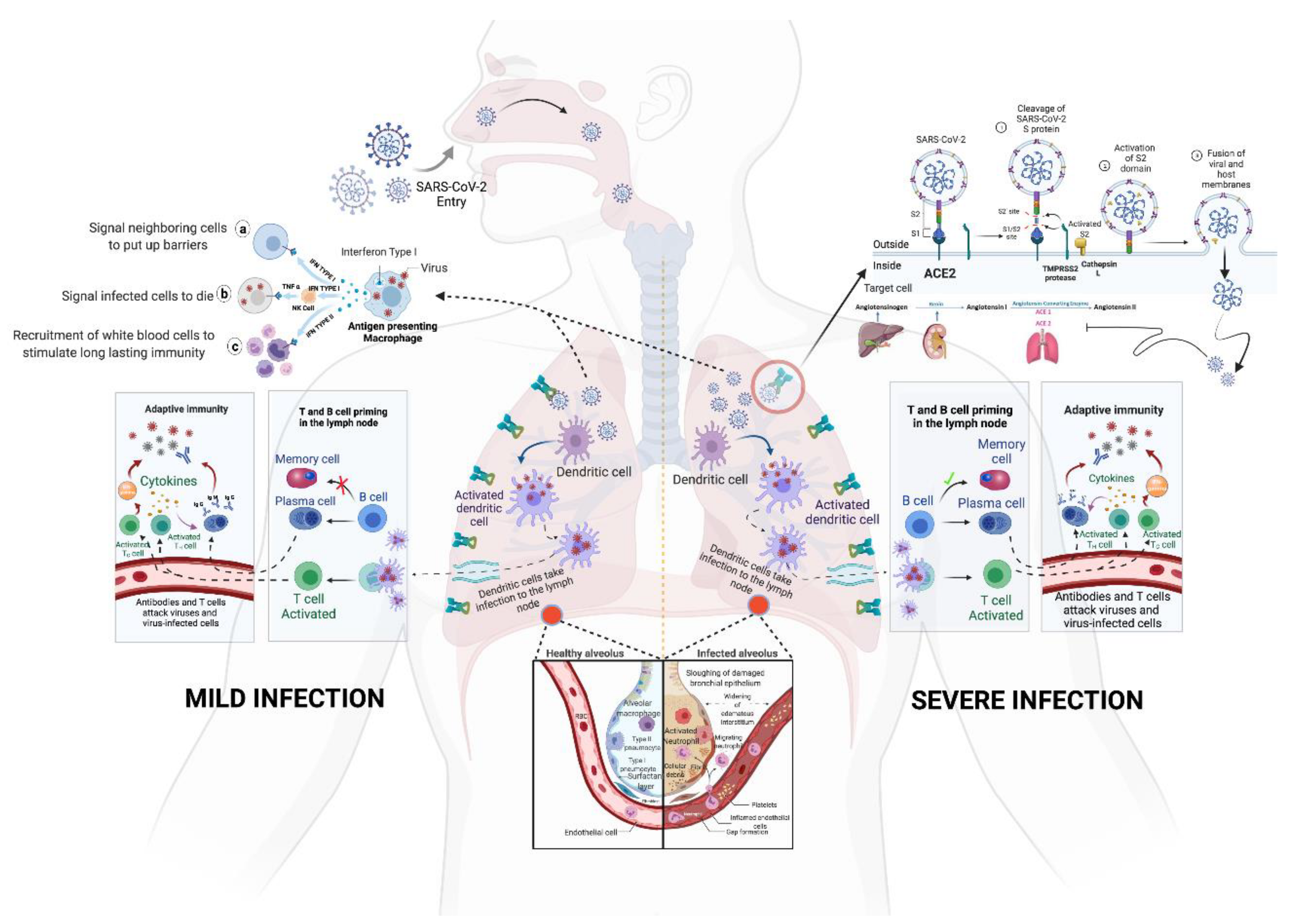
3. Immunopathology of SARS-CoV-2
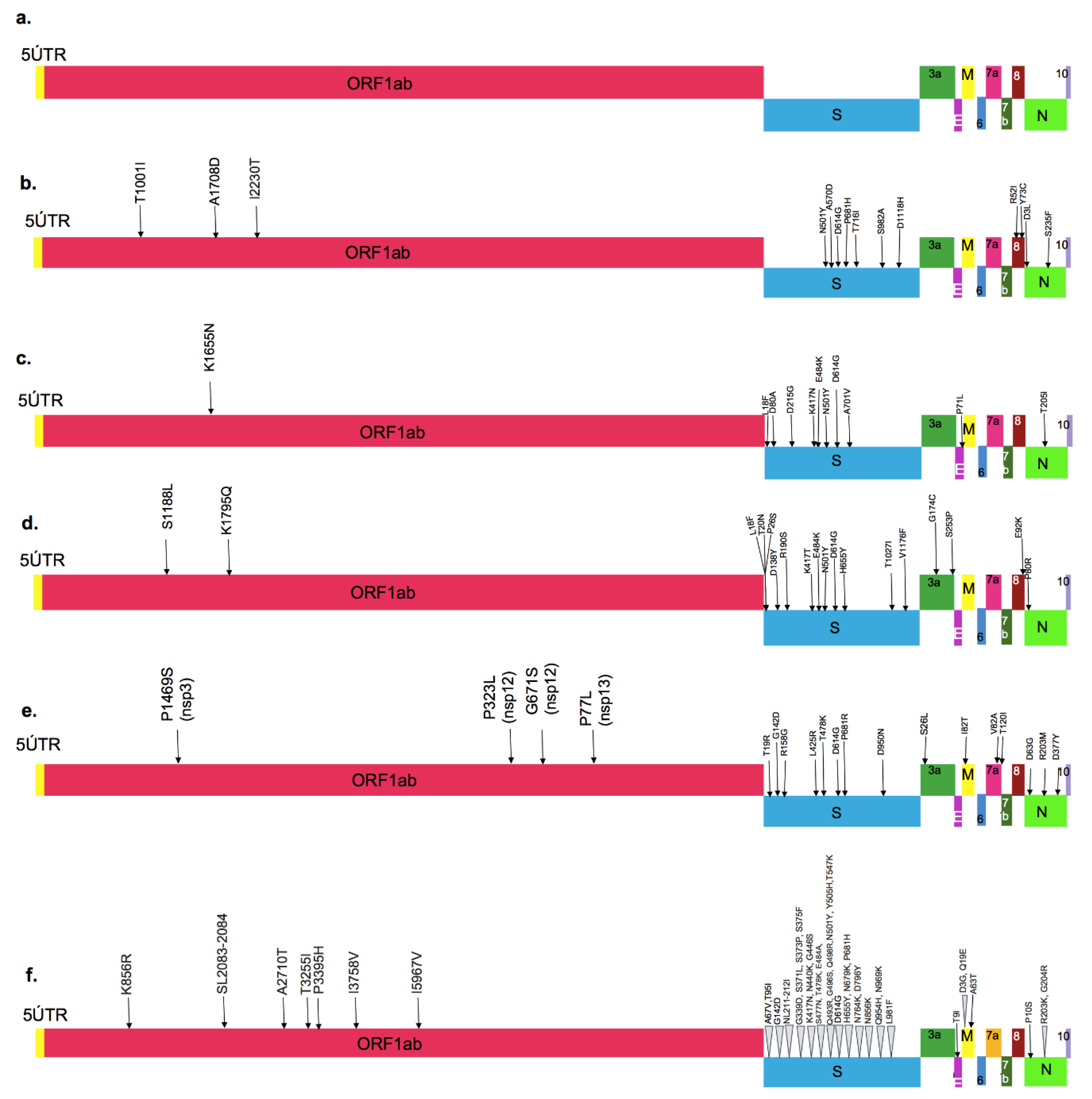
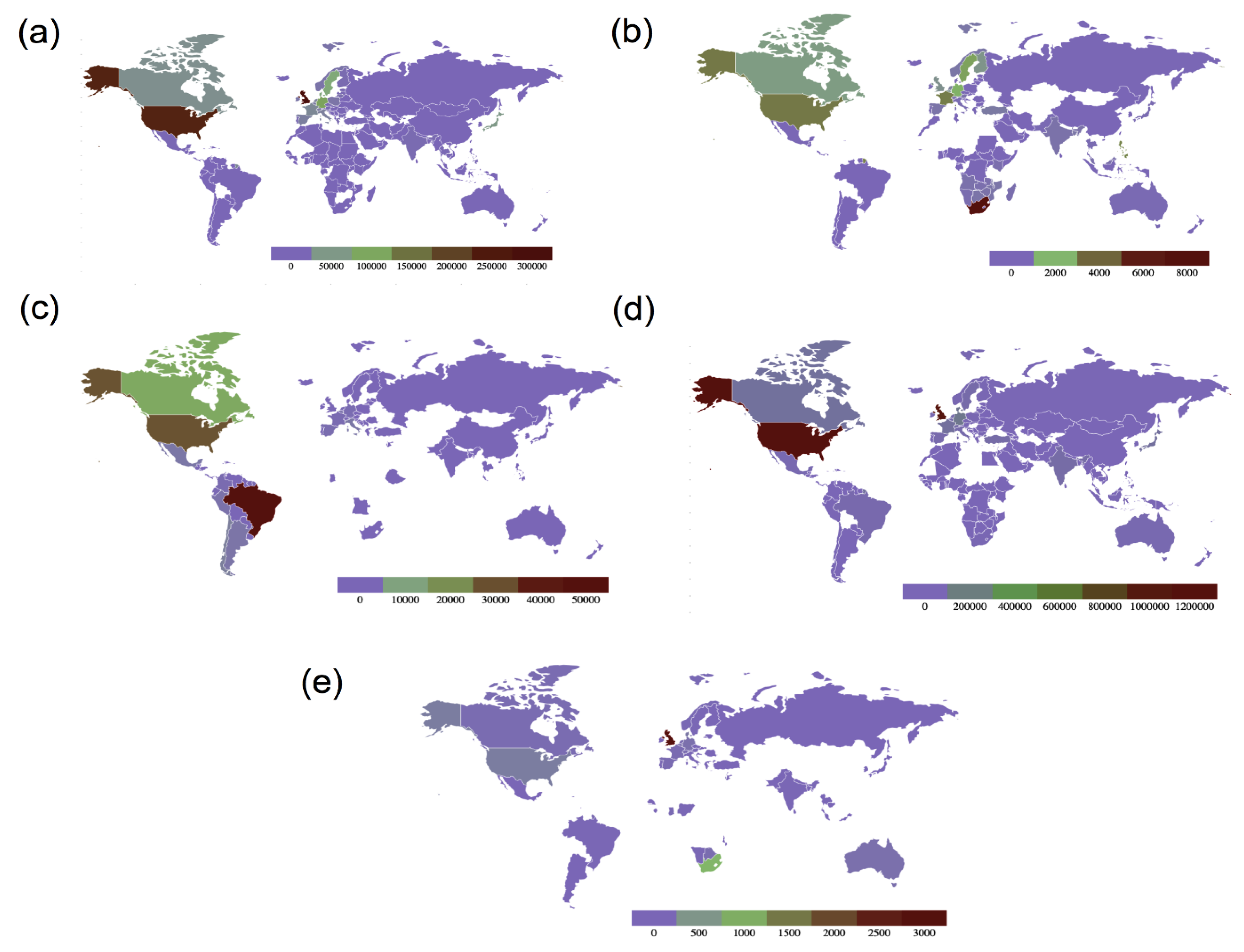
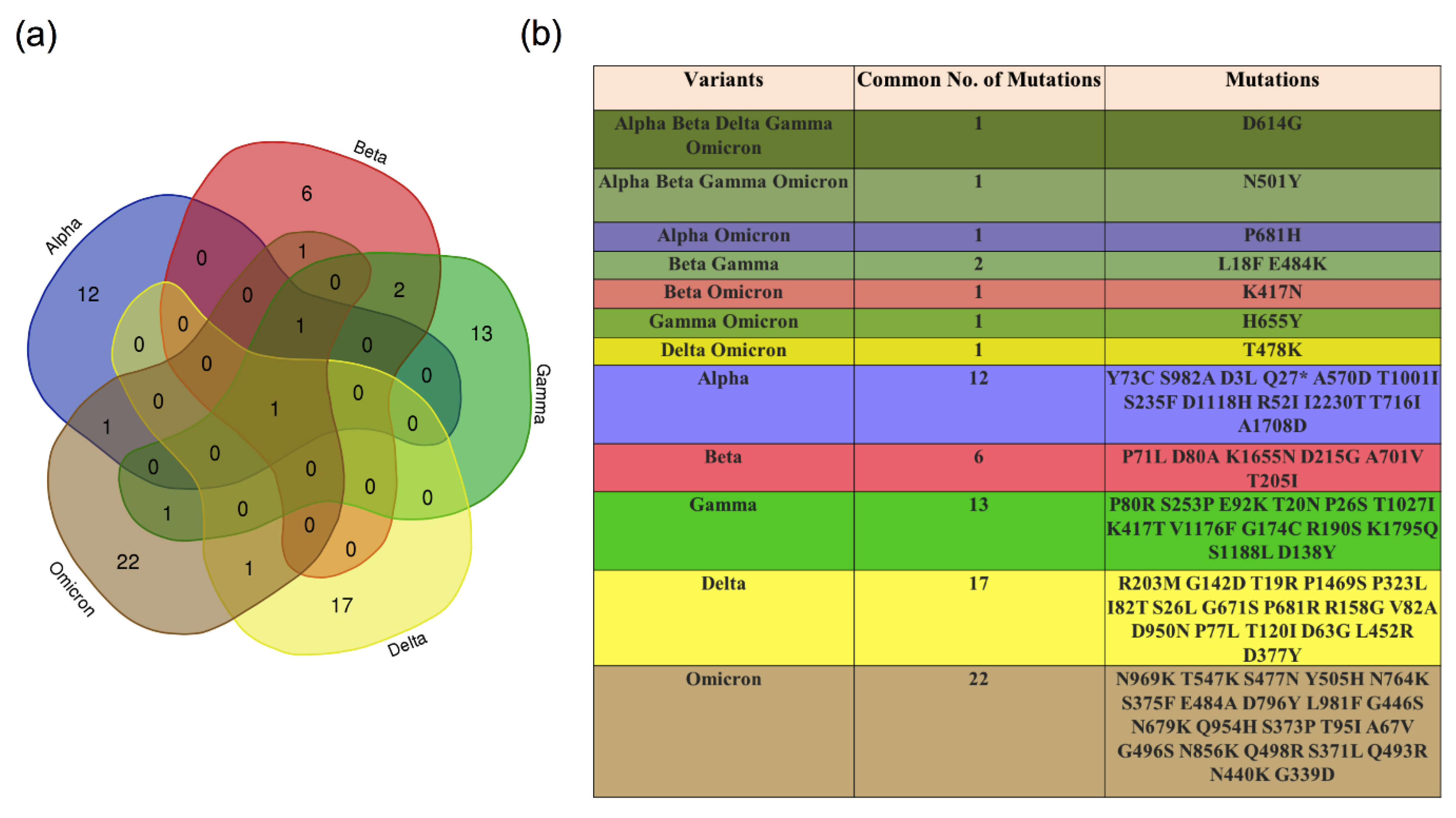
4. Variants of SARS-CoV-2
4.1. The D614G Variant
4.2. Variants of Concern
4.2.1. The Alpha Variant (B.1.1.7)
4.2.2. The Beta Variant (B.1.351)
4.2.3. The Gamma Variant (P.1 Variant; a Descendent of B.1.1.28)
4.2.4. The Delta Variant (B.1.617.2)
4.2.5. The Omicron Variant (B.1.1.529)
4.3. Variants of Interest
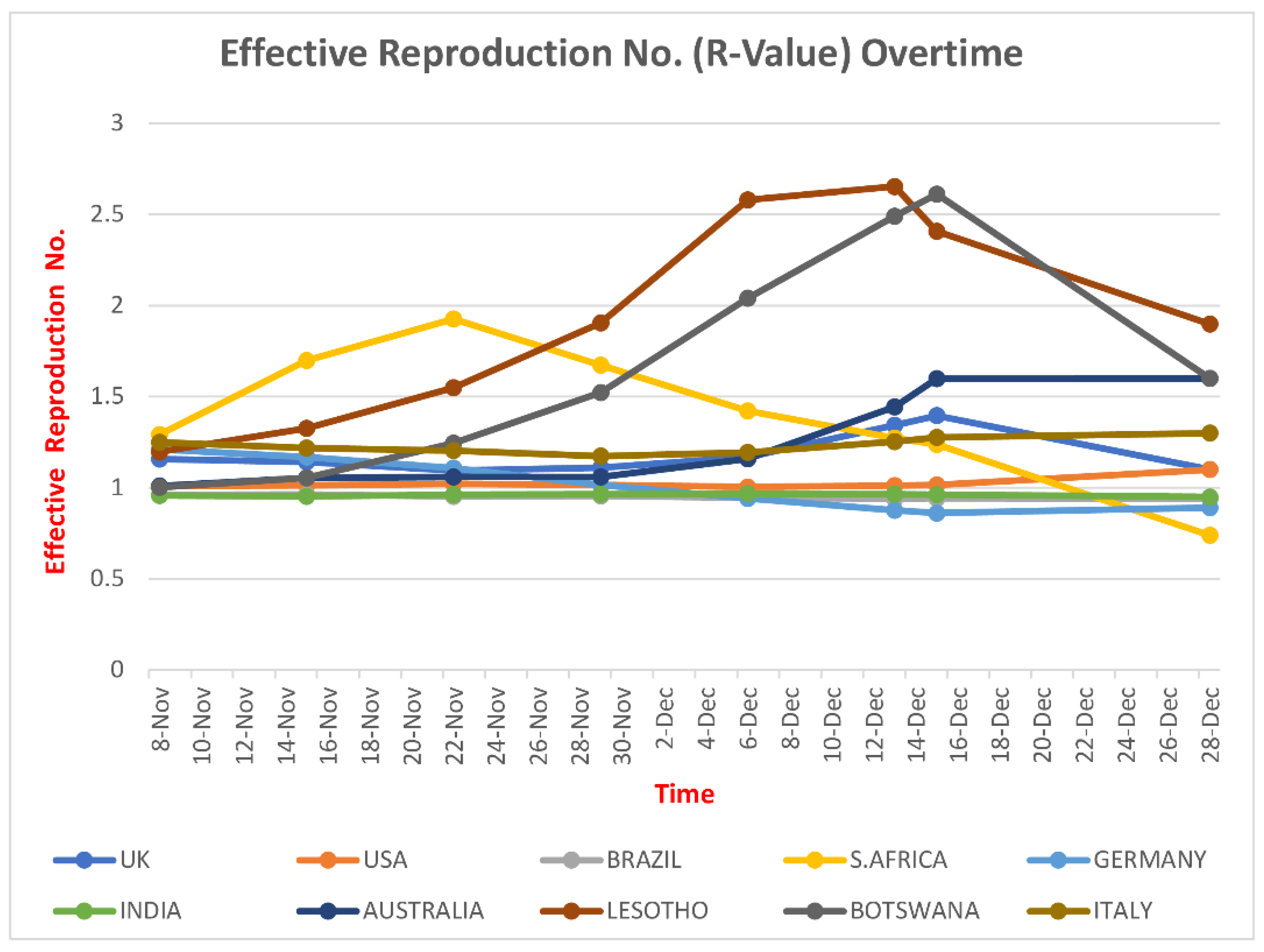
5. Infectious Potential of SARS-CoV-2
6. Impact of Variants on the Vaccines
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
- WHO. Coronavirus (COVID-19) Dashboard: https://covid19.who.int/ (accessed on 30 December 2021).
- Viral Zone. SARS-CoV-2 circulating variants: https://viralzone.expasy.org/9556 (accessed on 30 December 2021).
- Global Initiative on Sharing All Influenza Data (GISAID). EpiCoV: https://www.epicov.org/epi3/frontend#3a1b04 (accessed on 30 December 2021).
- CMMID nCoV working group: https://epiforecasts.io/covid/posts/global/ (accessed on 30 December 2021).
- CoV Lineage: Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool-Áine O’Toole, Emily Scher, Anthony Underwood, Ben Jackson, Verity Hill, John T McCrone, Rachel Colquhoun, Chris Ruis, Khalil Abu-Dahab, Ben Taylor, Corin Yeats, Louis Du Plessis, Daniel Maloney, Nathan Medd, Stephen W Attwood, David M Aanensen, Edward C Holmes, Oliver G Pybus, Andrew Rambaut (2021) Virus Evolution DOI:10.1093/ve/veab064 https://cov-lineages.org/ (accessed on 30 December 2021).
- CDC (Centre of Disease Control and Prevention): https://www.cdc.gov/ (accessed on 30 December 2021).
- NCBI Virus—https://www.ncbi.nlm.nih.gov/labs/virus/vssi/#/ (accessed on 30 December 2021).
- CoVariants (c) 2020–2021 (Enabled by GISAID)—https://covariants.org/ (accessed on 30 December 2021).
- Reactome—https://reactome.org/ (accessed on 30 December 2021).
- Investigation of SARS-CoV-2 variants of concern: Variant risk assessments—gov.uk—(accessed on 30 December 2021).
- https://www.gov.uk/government/publications/investigation-of-sars-cov-2-variants-of-concern-variant-risk-assessments (accessed on 30 December 2021).
Acknowledgments
Conflicts of Interest
Abbreviations
| SARS-CoV-2 | Severe Acute Respiratory Syndrome Coronavirus 2 |
| COVID-19 | Coronavirus Disease 2019 |
| VOC | Variant of Concern |
| VOI | Variant of Interest |
| ORF | Open Reading Frame |
| Nsp | Nonstructural protein |
| hACE-2 | human Angiotensin-Converting Enzyme 2 |
| RBD | Receptor Binding Domain |
| Aa | Amino Acid |
| NTDs | N Terminal Domains |
| CTDs | C Terminal Domains |
| Nt | Nucleotide (s) |
| MERS-CoV | Middle East respiratory syndrome—Coronavirus |
| SARS-CoV | Severe acute respiratory syndrome—Coronavirus |
| R0 | Basic reproduction number |
| Rt | Effective reproduction number |
| r | Epidemic growth rate |
| dt | Doubling time |
References
- Li, C.; Yang, Y.; Ren, L. Genetic evolution analysis of 2019 novel coronavirus and coronavirus from other species. Infect. Genet. Evol. 2020, 82, 104285. [Google Scholar] [CrossRef] [PubMed]
- Phartyal, R.; Verma, M. SARS-CoV-2 Mutations: An Insight. In Human Viruses: Diseases, Treatments and Vaccines—The New Insights; Ahmad, S.I., Ed.; Springer: Cham, Switzerland, 2021; pp. 551–563. ISBN 978-3-030-71165-8. [Google Scholar]
- Fathi, N.; Rezaei, N. Lymphopenia in COVID-19: Therapeutic opportunities. Cell Biol. Int. 2020, 44, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Thakur, V.; Kaur, P.; Khan, A.; Kulshrestha, S.; Kumar, P. Blood clots in COVID-19 patients: Simplifying the curious mystery. Med. Hypotheses 2021, 146, 110371. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Patidar, R.; Younis, K.; Desai, P.; Hosein, Z.; Padda, I.; Mangat, J.; Altaf, M. Comorbidity and its Impact on Patients with COVID-19. SN Compr. Clin. Med. 2020, 2, 1069–1076. [Google Scholar] [CrossRef]
- How to Redesign COVID Vaccines so They Protect against Variants. Available online: https://www.nature.com/articles/d41586-021-00241-6 (accessed on 10 October 2021).
- Gupta, V.; Haider, S.; Verma, M.; Singhvi, N.; Ponnusamy, K.; Malik, M.Z.; Verma, H.; Kumar, R.; Sood, U.; Hira, P.; et al. Comparative Genomics and Integrated Network Approach Unveiled Undirected Phylogeny Patterns, Co-mutational Hot Spots, Functional Cross Talk, and Regulatory Interactions in SARS-CoV-2. MSystems 2021, 6, e00030-21. [Google Scholar] [CrossRef]
- Kumar, R.; Verma, H.; Singhvi, N.; Sood, U.; Gupta, V.; Singh, M.; Kumari, R.; Hira, P.; Nagar, S.; Talwar, C.; et al. Comparative Genomic Analysis of Rapidly Evolving SARS-CoV-2 Reveals Mosaic Pattern of Phylogeographical Distribution. MSystems 2021, 5, e00505-20. [Google Scholar] [CrossRef]
- Boni, M.F.; Lemey, P.; Jiang, X.; Lam, T.T.-Y.; Perry, B.W.; Castoe, T.A.; Rambaut, A.; Robertson, D.L. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat. Microbiol. 2020, 5, 1408–1417. [Google Scholar] [CrossRef]
- Redondo, N.; Zaldívar-López, S.; Garrido, J.J.; Montoya, M. SARS-CoV-2 Accessory Proteins in Viral Pathogenesis: Knowns and Unknowns. Front. Immunol. 2021, 12, 708264. [Google Scholar] [CrossRef]
- Harrison, A.G.; Lin, T.; Wang, P. Mechanisms of SARS-CoV-2 Transmission and Pathogenesis. Trends Immunol. 2020, 41, 1100–1115. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]
- Soh, S.M.; Kim, Y.; Kim, C.; Jang, U.S.; Lee, H.-R. The rapid adaptation of SARS-CoV-2–rise of the variants: Transmission and resistance. J. Microbiol. 2021, 59, 807–818. [Google Scholar] [CrossRef]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Hasöksüz, M.; Kiliç, S.; Saraç, F. Coronaviruses and SARS-COV-2. Turk. J. Med. Sci. 2020, 50, 549–556. [Google Scholar] [CrossRef]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Turoňová, B.; Sikora, M.; Schürmann, C.; Hagen, W.J.H.; Welsch, S.; Blanc, F.E.C.; Von Bülow, S.; Gecht, M.; Bagola, K.; Hörner, C.; et al. In situ structural analysis of SARS-CoV-2 spike reveals flexibility mediated by three hinges. Science 2020, 370, 203–208. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.; Xu, W.; Liu, S. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Tang, T.; Bidon, M.; Jaimes, J.A.; Whittaker, G.R.; Daniel, S. Coronavirus membrane fusion mechanism offers a potential target for antiviral development. Antivir. Res. 2020, 178, 104792. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904. [Google Scholar] [CrossRef]
- Gobeil, S.M.; Janowska, K.; McDowell, S.; Mansouri, K.; Parks, R.; Stalls, V.; Kopp, M.F.; Manne, K.; Saunders, K.; Edwards, R.J.; et al. Effect of natural mutations of SARS-CoV-2 on spike structure, conformation, and antigenicity. Science 2021, 373, eabi6226. [Google Scholar] [CrossRef]
- Majumdar, P.; Niyogi, S. ORF3a mutation associated with higher mortality rate in SARS-CoV-2 infection. Epidemiol. Infect. 2020, 148, e262. [Google Scholar] [CrossRef] [PubMed]
- Pachetti, M.; Marini, B.; Benedetti, F.; Giudici, F.; Mauro, E.; Storici, P.; Masciovecchio, C.; Angeletti, S.; Ciccozzi, M.; Gallo, R.C.; et al. Emerging SARS-CoV-2 mutation hot spots include a novel RNA-dependent-RNA polymerase variant. J. Transl. Med. 2020, 18, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Alfajaro, M.M.; DeWeirdt, P.C.; Hanna, R.E.; Lu-Culligan, W.J.; Cai, W.L.; Strine, M.S.; Zhang, S.-M.; Graziano, V.R.; Schmitz, C.O.; et al. Genome-wide CRISPR Screens Reveal Host Factors Critical for SARS-CoV-2 Infection. Cell 2021, 184, 76–91.e13. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Wang, M.-L.; Chien, C.-S.; Yarmishyn, A.A.; Yang, Y.-P.; Lai, W.-Y.; Luo, Y.-H.; Lin, Y.-T.; Chen, Y.-J.; Chang, P.-C.; et al. Highlight of Immune Pathogenic Response and Hematopathologic Effect in SARS-CoV, MERS-CoV, and SARS-Cov-2 Infection. Front. Immunol. 2020, 11, 1022. [Google Scholar] [CrossRef]
- Daniloski, Z.; Jordan, T.X.; Wessels, H.-H.; Hoagland, D.A.; Kasela, S.; Legut, M.; Maniatis, S.; Mimitou, E.P.; Lu, L.; Geller, E.; et al. Identification of Required Host Factors for SARS-CoV-2 Infection in Human Cells. Cell 2021, 184, 92–105.e16. [Google Scholar] [CrossRef]
- Torbati, E.; Krause, K.L.; Ussher, J.E. The Immune Response to SARS-CoV-2 and Variants of Concern. Viruses 2021, 13, 1911. [Google Scholar] [CrossRef]
- Cheng, Y.-R.; Li, X.; Zhao, X.; Lin, H. Cell Entry of Animal Coronaviruses. Viruses 2021, 13, 1977. [Google Scholar] [CrossRef]
- Shah, V.K.; Firmal, P.; Alam, A.; Ganguly, D.; Chattopadhyay, S. Overview of Immune Response During SARS-CoV-2 Infection: Lessons From the Past. Front. Immunol. 2020, 11, 1949. [Google Scholar] [CrossRef]
- Catanzaro, M.; Fagiani, F.; Racchi, M.; Corsini, E.; Govoni, S.; Lanni, C. Immune response in COVID-19: Addressing a pharmacological challenge by targeting pathways triggered by SARS-CoV-2. Signal Transduct. Target. Ther. 2020, 5, 84. [Google Scholar] [CrossRef]
- De Wit, E.; van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Graham, M.S.; Sudre, C.H.; May, A.; Antonelli, M.; Murray, B.; Varsavsky, T.; Kläser, K.; Canas, L.S.; Molteni, E.; Modat, M.; et al. Changes in symptomatology, reinfection, and transmissibility associated with the SARS-CoV-2 variant B.1.1.7: An ecological study. Lancet Public Health 2021, 6, e335–e345. [Google Scholar] [CrossRef]
- Mehra, R.; Kepp, K.P. Structure and Mutations of SARS-CoV-2 Spike Protein: A Focused Overview. ACS Infect. Dis. 2022, 8, 29–58. [Google Scholar] [CrossRef]
- Harvey, W.T.; Carabelli, A.M.; Jackson, B.; Gupta, R.K.; Thomson, E.C.; Harrison, E.M.; Ludden, C.; Reeve, R.; Rambaut, A.; Peacock, S.J.; et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 2021, 19, 409–424. [Google Scholar] [CrossRef]
- Winger, A.; Caspari, T. The Spike of Concern—The Novel Variants of SARS-CoV-2. Viruses 2021, 13, 1002. [Google Scholar] [CrossRef]
- Luan, B.; Wang, H.; Huynh, T. Molecular Mechanism of the N501Y Mutation for Enhanced Binding between SARS-CoV-2’s Spike Protein and Human ACE2 Receptor. BioRxiv 2021, BioRxiv:2021.01.04.425316. [Google Scholar]
- Chakraborti, P.K.; Matange, N.; Nandicoori, V.K.; Singh, Y.; Tyagi, J.S.; Visweswariah, S.S. Signalling mechanisms in Mycobacteria. Tuberculosis 2011, 91, 432–440. [Google Scholar] [CrossRef]
- Barton, M.I.; MacGowan, S.A.; Kutuzov, M.A.; Dushek, O.; Barton, G.J.; van der Merwe, P.A. Effects of common mutations in the SARS-CoV-2 Spike RBD and its ligand, the human ACE2 receptor on binding affinity and kinetics. eLife 2021, 10, e70658. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Mercatelli, D.; Rakhimov, A.; Giorgi, F.M. Preliminary report on severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) Spike mutation T478K. J. Med. Virol. 2021, 93, 5638–5643. [Google Scholar] [CrossRef]
- Cherian, S.; Potdar, V.; Jadhav, S.; Yadav, P.; Gupta, N.; Das, M.; Rakshit, P.; Singh, S.; Abraham, P.; Panda, S.; et al. Convergent Evolution of SARS-CoV-2 Spike Mutations, L452R, E484Q and P681R, in the Second Wave of COVID-19 in Maharashtra, India. BioRxiv 2021, BioRxiv:2021.04.22.440932. [Google Scholar]
- Teo, S.P. Review of COVID-19 mRNA Vaccines: BNT162b2 and mRNA-1273. J. Pharm. Pract. 2021, 8971900211009650. [Google Scholar] [CrossRef] [PubMed]
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation, and Treatment of Coronavirus (COVID-19). In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Isabel, S.; Graña-Miraglia, L.; Gutierrez, J.M.; Bundalovic-Torma, C.; Groves, H.E.; Isabel, M.R.; Eshaghi, A.; Patel, S.N.; Gubbay, J.B.; Poutanen, T.; et al. Evolutionary and structural analyses of SARS-CoV-2 D614G spike protein mutation now documented worldwide. Sci. Rep. 2020, 10, 14031. [Google Scholar] [CrossRef] [PubMed]
- Bian, L.; Gao, F.; Zhang, J.; He, Q.; Mao, Q.; Xu, M.; Liang, Z. Effects of SARS-CoV-2 variants on vaccine efficacy and response strategies. Expert Rev. Vaccines 2021, 20, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.B.; Zhang, L.; Farzan, M.; Choe, H. Functional importance of the D614G mutation in the SARS-CoV-2 spike protein. Biochem. Biophys. Res. Commun. 2021, 538, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Daniloski, Z.; Jordan, T.X.; Ilmain, J.K.; Guo, X.; Bhabha, G.; tenOever, B.R.; Sanjana, N.E. The Spike D614G mutation increases SARS-CoV-2 infection of multiple human cell types. eLife 2021, 10, e65365. [Google Scholar] [CrossRef]
- Shorthouse, D.; Hall, B.A. SARS-CoV-2 Variants are Selecting for Spike Protein Mutations that Increase Protein Stability. J. Chem. Inf. Modeling 2021, 61, 4152–4155. [Google Scholar] [CrossRef]
- Laiton-Donato, K.; Franco-Muñoz, C.; Álvarez-Díaz, D.A.; Ruiz-Moreno, H.A.; Usme-Ciro, J.A.; Prada, D.A.; Reales-González, J.; Corchuelo, S.; Herrera-Sepúlveda, M.T.; Naizaque, J.; et al. Characterization of the Emerging B.1.621 Variant of Interest of SARS-CoV-2. Infect. Genet. Evol. 2021, 95, 105038. [Google Scholar] [CrossRef]
- Leung, K.; Shum, M.H.; Leung, G.M.; Lam, T.T.; Wu, J.T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Eurosurveillance 2021, 26, 2002106. [Google Scholar] [CrossRef]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; de Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The Biological and Clinical Significance of Emerging SARS-CoV-2 Variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Tian, D.; Sun, Y.; Zhou, J.; Ye, Q. The Global Epidemic of SARS-CoV-2 Variants and Their Mutational Immune Escape. J. Med. Virol. 2021, 84, 847–857. [Google Scholar] [CrossRef]
- Abdool Karim, S.S.; de Oliveira, T. New SARS-CoV-2 Variants—Clinical, Public Health, and Vaccine Implications. N. Engl. J. Med. 2021, 384, 1866–1868. [Google Scholar] [CrossRef]
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N.; et al. Detection of a SARS-CoV-2 variant of concern in South Africa. Nature 2021, 592, 438–443. [Google Scholar] [CrossRef]
- Gan, H.H.; Twaddle, A.; Marchand, B.; Gunsalus, K.C. Structural Modeling of the SARS-CoV-2 Spike/Human ACE2 Complex Interface can Identify High-Affinity Variants Associated with Increased Transmissibility. J. Mol. Biol. 2021, 433, 167051. [Google Scholar] [CrossRef]
- Naveca, F.; Nascimento, V.; Souza, V.; de Lima Corado, A.; Nascimento, F.; Mejía, M.; Brandão, M.J.; da Silva, A.V.; Benzaken, A.S.; Silva, G.; et al. The SARS-CoV-2 Variant Delta Displaced the Variants Gamma and Gamma Plus in Amazonas, Brazil. Available online: https://virological.org/t/the-sars-cov-2-variant-delta-displaced-the-variants-gamma-and-gamma-plus-in-amazonas-brazil/765 (accessed on 30 December 2021).
- Roy, B.; Roy, H. The Delta Plus variant of COVID-19: Will it be the worst nightmare in the SARS-CoV-2 pandemic? J. Biomed. Sci. 2021, 8, 1–2. [Google Scholar] [CrossRef]
- Delta Variant is Causing a Surge in Covid Cases Globally—Quartz India. Available online: https://qz.com/india/2036052/delta-variant-is-causing-a-surge-in-covid-cases-globally/ (accessed on 30 December 2021).
- Gupta, N.; Kaur, H.; Yadav, P.D.; Mukhopadhyay, L.; Sahay, R.R.; Kumar, A.; Nyayanit, D.A.; Shete, A.M.; Patil, S.; Majumdar, T.; et al. Clinical Characterization and Genomic Analysis of Samples from COVID-19 Breakthrough Infections during the Second Wave among the Various States of India. Viruses 2021, 13, 1782. [Google Scholar] [CrossRef]
- Kandeel, M.; Mohamed, M.E.M.; Abd El-Lateef, H.M.; Venugopala, K.N.; El-Beltagi, H.S. Omicron variant genome evolution and phylogenetics. J. Med. Virol. 2022, 94, 1627–1632. [Google Scholar] [CrossRef]
- Gao, S.-J.; Guo, H.; Luo, G. Omicron variant (B.1.1.529) of SARS-CoV-2, a global urgent public health alert! J. Med. Virol. 2022, 94, 1225. [Google Scholar] [CrossRef]
- Severin Lupala, C.; Ye, Y.; Chen, H.; Su, X.; Liu, H. Mutations in RBD of SARS-CoV-2 Omicron Variant Result Stronger Binding to Human ACE2 Protein. Biochem. Biophys. Res. Commun. 2022, 590, 34–41. [Google Scholar] [CrossRef]
- Golcuk, M.; Yildiz, A.; Gur, M. The Omicron Variant Increases the Interactions of SARS-CoV-2 Spike Glycoprotein with ACE2. bioRxiv 2021, bioRxiv:2021.12.06.471377. [Google Scholar]
- Wilhelm, A.; Widera, M.; Grikscheit, K.; Toptan, T.; Schenk, B.; Pallas, C.; Metzler, M.; Kohmer, N.; Hoehl, S.; Helfritz, F.; et al. Reduced Neutralization of SARS-CoV-2 Omicron Variant by Vaccine Sera and Monoclonal Antibodies. medRxiv 2021, medRxiv:2021.12.07.21267432. [Google Scholar]
- Ranjan, P.; Neha; Devi, C.; Devar, K.A.; Das, P. The influence of New SARS-CoV-2 Variant Omicron (B.1.1.529) on Vaccine Efficacy, Its Correlation to Delta Variants: A Computational Approach. bioRxiv 2021, bioRxiv:2021.12.06.471215. [Google Scholar]
- Callaway, E. Omicron likely to weaken COVID vaccine protection. Nature 2021, 600, 367–368. [Google Scholar] [CrossRef]
- Saxena, S.K.; Kumar, S.; Ansari, S.; Paweska, J.T.; Maurya, V.K.; Tripathi, A.K.; Abdel-Moneim, A.S. Characterization of the novel SARS-CoV-2 Omicron (B.1.1.529) Variant of Concern and its global perspective. J. Med. Virol. 2022, 94, 1738–1744. [Google Scholar] [CrossRef]
- Del Águila-Mejía, J.; Wallmann, R.; Calvo-Montes, J.; Rodríguez-Lozano, J.; Valle-Madrazo, T.; Aginagalde-Llorente, A. Secondary Attack Rate, Transmission and Incubation Periods, and Serial Interval of SARS-CoV-2 Omicron Variant, Spain. Emerg. Infect. Dis. 2022, 28, 1224–1228. [Google Scholar] [CrossRef]
- Liu, Y.; Rocklöv, J. The reproductive number of the Delta variant of SARS-CoV-2 is far higher compared to the ancestral SARS-CoV-2 virus. J. Travel Med. 2021, 28, taab124. [Google Scholar] [CrossRef]
- Inglesby, T.V. Public Health Measures and the Reproduction Number of SARS-CoV-2. JAMA 2020, 323, 2186–2187. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.N.; Shah, J.; Shah, J.N. Quarantine, isolation and lockdown: In context of COVID-19. J. Patan Acad. Health Sci. 2020, 7, 48–57. [Google Scholar] [CrossRef]
- Halley, J.M.; Vokou, D.; Pappas, G.; Sainis, I. Evolving SARS-CoV-2 Variants and Mutational Cascades. medRxiv 2021, medRxiv:2021.04.03.21254871. Available online: https://www.medrxiv.org/content/10.1101/2021.04.03.21254871v1 (accessed on 10 October 2021).
- Tregoning, J.S.; Flight, K.E.; Higham, S.L.; Wang, Z.; Pierce, B.F. Progress of the COVID-19 vaccine effort: Viruses, vaccines and variants versus efficacy, effectiveness and escape. Nat. Rev. Immunol. 2021, 21, 626–636. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383.e9. [Google Scholar] [CrossRef]
- Knoll, M.D.; Wonodi, C. Oxford-AstraZeneca COVID-19 vaccine efficacy. Lancet 2021, 397, 72–74. [Google Scholar] [CrossRef]
- Mahase, E. Covid-19: Moderna applies for US and EU approval as vaccine trial reports 94.1% efficacy. BMJ 2020, 371, m4709. [Google Scholar] [CrossRef] [PubMed]
- Sadoff, J.; Gray, G.; Vandebosch, A.; Cárdenas, V.; Shukarev, G.; Grinsztejn, B.; Goepfert, P.A.; Truyers, C.; Fennema, H.; Spiessens, B.; et al. Safety and Efficacy of Single-Dose Ad26.COV2.S Vaccine against Covid-19. N. Engl. J. Med. 2021, 384, 2187–2201. [Google Scholar] [CrossRef] [PubMed]
- Heath, P.T.; Galiza, E.P.; Baxter, D.N.; Boffito, M.; Browne, D.; Burns, F.; Chadwick, D.R.; Clark, R.; Cosgrove, C.; Galloway, J.; et al. Safety and Efficacy of NVX-CoV2373 Covid-19 Vaccine. N. Engl. J. Med. 2021, 385, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Costa Clemens, S.A.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: A pooled analysis of four randomised trials. Lancet 2021, 397, 881–891. [Google Scholar] [CrossRef]
- Chang, X.; Augusto, G.S.; Liu, X.; Kündig, T.M.; Vogel, M.; Mohsen, M.O.; Bachmann, M.F. BNT162b2 mRNA COVID-19 vaccine induces antibodies of broader cross-reactivity than natural infection, but recognition of mutant viruses is up to 10-fold reduced. Allergy 2021, 76, 2895–2998. [Google Scholar] [CrossRef]
- Immune Imprinting Causes Varied Patterns of Protection against COVID-19 Variants. Available online: https://www.imperial.ac.uk/news/232396/immune-imprinting-causes-varied-patterns-protection/ (accessed on 31 December 2021).
- Khan, W.H.; Hashmi, Z.; Goel, A.; Ahmad, R.; Gupta, K.; Khan, N.; Alam, I.; Ahmed, F.; Ansari, M.A. COVID-19 Pandemic and Vaccines Update on Challenges and Resolutions. Front. Cell. Infect. Microbiol. 2021, 11, 690621. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Butt, A.A. Effectiveness of the BNT162b2 Covid-19 Vaccine against the B.1.1.7 and B.1.351 Variants. N. Engl. J. Med. 2021, 385, 187–189. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- mRNA-1273 COVID-19 Vaccine Effectiveness against the B.1.1.7 and B.1.351 Variants and Severe COVID-19 Disease in Qatar. Nature Medicine. Available online: https://www.nature.com/articles/s41591-021-01446-y (accessed on 28 December 2021).
- AstraZeneca’s COVID-19 Vaccine Shows Effectiveness against Indian Variants of SARS-CoV-2 Virus. Available online: https://www.astrazeneca.com/media-centre/articles/2021/astrazenecas-covid-19-vaccine-shows-effectiveness-against-indian-variants-of-sars-cov-2-virus.html (accessed on 28 December 2021).
- Sanderson, K. COVID Vaccines Protect against Delta, but Their Effectiveness Wanes. Nature 2021. [Google Scholar] [CrossRef]
- Zinatizadeh, M.R.; Zarandi, P.K.; Zinatizadeh, M.; Yousefi, M.H.; Amani, J.; Rezaei, N. Efficacy of mRNA, adenoviral vector, and perfusion protein COVID-19 vaccines. Biomed. Pharmacother. 2022, 146, 112527. [Google Scholar] [CrossRef]
- Johnson & Johnson Announces Real-World Evidence and Phase 3 Data Confirming Strong and Long-Lasting Protection of Single-Shot COVID-19 Vaccine in the U.S. Available online: https://www.jnj.com/johnson-johnson-announces-real-world-evidence-and-phase-3-data-confirming-strong-and-long-lasting-protection-of-single-shot-covid-19-vaccine-in-the-u-s (accessed on 28 December 2021).
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Emary, K.R.W.; Golubchik, T.; Aley, P.K.; Ariani, C.V.; Angus, B.; Bibi, S.; Blane, B.; Bonsall, D.; Cicconi, P.; Charlton, S.; et al. Efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 variant of concern 202012/01 (B.1.1.7): An exploratory analysis of a randomised controlled trial. Lancet 2021, 397, 1351–1362. [Google Scholar] [CrossRef]
- Jara, A.; Undurraga, E.A.; González, C.; Paredes, F.; Fontecilla, T.; Jara, G.; Pizarro, A.; Acevedo, J.; Leo, K.; Leon, F.; et al. Effectiveness of an Inactivated SARS-CoV-2 Vaccine in Chile. N. Engl. J. Med. 2021, 385, 875–884. [Google Scholar] [CrossRef]
- Cromer, D.; Steain, M.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Kent, S.J.; Triccas, J.A.; Khoury, D.S.; Davenport, M.P. Neutralising antibody titres as predictors of protection against SARS-CoV-2 variants and the impact of boosting: A meta-analysis. Lancet Microbe 2021, 3, e52–e61. [Google Scholar] [CrossRef]
- Sapkota, B.; Saud, B.; Shrestha, R.; Al-Fahad, D.; Sah, R.; Shrestha, S.; Rodriguez-Morales, A.J. Heterologous prime–boost strategies for COVID-19 vaccines. J. Travel Med. 2021, taab191. [Google Scholar] [CrossRef]
- Li, R.; Liu, J.; Zhang, H. The challenge of emerging SARS-CoV-2 mutants to vaccine development. J. Genet. Genom. 2021, 48, 102–106. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WHO Label | Pango Lineage | Potential Emergence | Number of Countries Affected | Important Spike Mutations | Sequenced Genomes (GISAID) | Impact of Mutations | Transmissibility of Variant |
|---|---|---|---|---|---|---|---|
| Alpha | B.1.1.7 | UK, September 2020 | 170 | N501Y P681H | 11,49,443 | P681H: Slightly increases the S1–S2 cleavage [37]. N501Y: Enhances binding affinity between spike RBD domain and hACE2 receptor [38]. E484K *, L452R * | ~70% increase with respect to wild type |
| Beta | B.1.351 | South Africa, May 2020 | 112 | K417N E484K N501Y | 39,604 | K417N: Responsible for antibody-escape property of the virus [39]. N501Y: Same as above. E484K: Reduces antibody neutralisation and is responsible for increased infectivity [40]. | 20–113% increase with respect to wild type |
| Gamma | P.1 | Brazil, November 2020 | 80 | K417T E484K N501Y | 1,17,675 | K417T: Responsible for antibody escape property of the virus [39]. N501Y: Same as above. E484K: Same as above. | ~161% increase with respect to wild type |
| Delta | B.1.617.2 | India, October 2020 | 105 | L452R T478K P681R | 34,37,588 | T478K: Helps evade neutralising antibodies. Enhances viral infectivity [41]. L452R: Impairs antibody neutralization. Increases viral infectivity [39,42]. P681R: Increases the rate of S1–S2 cleavage, and hence results in enhanced transmissibility [40]. |
|
| Omicron | B.1.1.529 | Multiple countries, November 2021 | - | H655Y N679K P681H E484A Q498R N501Y | 5352 | N501Y: Same as above. H655Y: May be associated with increased transmissibility as it is proximal to the Furin cleavage site, hence aids in increased spike cleavage (CoVariants-GISAID). N679K: Associated with increased transmissibility (CoVariants-GISAID). P681H: Slightly increases the S1–S2 cleavage [37]. E484A: May be associated with immune escape (CoVariants-GISAID). Q498R: Enhances binding affinity with ACE2 receptor in combination with N501Y (CoVariants-GISAID). | - |
| WHO Label | Pango Lineage | Emergence | RBD Mutations | Other Spike Mutations |
|---|---|---|---|---|
| Eta * | B.1.525 | Multiple countries, December 2020 | E484K | O52R, Q677H, F888L, D614G, A67V |
| Iota * | B.1.526 | United States of America, November 2020 | E484K, S447N | L5F, T95I, D253G, D614G, A701V |
| Kappa * | B.1.617.1 | India, October 2020 | E484Q, L452R, | G142D, D614G, T95I, E154K, Q1071H, M153I |
| Lambda | C.37 | Peru, December 2020 | L452Q | G75V, T76I F490S, D614G T859N |
| Mu | B.1.621 | Colombia, January 2021 | R346K, E484K, N501Y | T95I, Y144T, Y145S, P618H |
| Name of Vaccine | Vaccine Type | Efficacy | Effectiveness against Variants (B.1.1.7/alpha (United Kingdom), B.1.351/beta (South Africa), P.1/gamma (Brazil), B.1.617/delta (India)) | |
|---|---|---|---|---|
| First Dose | Second Dose | |||
| Pfizer-BioNTech (BNT162b2) | mRNA | 92.6% | 95.0% | Alpha (B.1.1.7)–87.0% Beta (B.1.351)–75.0% Gamma (P.1)–82.0% Delta (B.1.617.2)–95.0% |
| Moderna (mRNA-1273) | mRNA | 81.6% | 94.1% | Alpha (B.1.1.7)–81.6% Beta (B.1.351)–95.7% Gamma (P.1)–89.0% Delta (B.1.617.2)–95.0% |
| AstraZeneca, University of Oxford (AZD1222) | Adenovirus-based | 90.0% | 62.1% | Alpha (B.1.1.7)–70.4% Beta (B.1.351)–77.0% Gamma (P.1)–82.0% Delta (B.1.617.2)–67.0% |
| Johnson & Johnson (Ad26.COV2. S) | Non-replicating viral vector | 66.0% | 73.1% | Alpha (B.1.1.7)–74.0% Beta (B.1.351)–52.0% Gamma (P.1)–66.0% Delta (B.1.617.2)–60.0% |
| BBIBP-CorV | Inactivated virus vaccine | 79.0% | Efficacious in preventing PCR-confirmed COVID-19 in adults. No vaccine effectiveness studies yet to inform protection against variants of concern (CDC, 2021) | |
| Novavax (NVX-CoV2373) | Protein-based vaccine | 83.4% | 89.7% | Alpha (B.1.1.7)–86.3% Beta (B.1.351)–51.0% Non-B.1.1.7–96.0% |
| Covishield (ChAdOx1 nCoV-19) | Adenovirus-based vaccine | 64.0% | 70.4% | Alpha (B.1.1.7)–70.4% Non-B.1.1.7–81.5% Gamma (P.1)–N.A Delta (B.1.617.2)–87.0% |
| CoronaVac | Inactivated virus vaccine | 50.0% | 50.0–91.0% | More data are needed on its efficacy against emerging SARS-CoV-2 variants |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chugh, A.; Khurana, N.; Verma, K.; Sehgal, I.; Rolta, R.; Vats, P.; Phartyal, R.; Salaria, D.; Kaushik, N.; Choi, E.H.; et al. Changing Dynamics of SARS-CoV-2: A Global Challenge. Appl. Sci. 2022, 12, 5546. https://doi.org/10.3390/app12115546
Chugh A, Khurana N, Verma K, Sehgal I, Rolta R, Vats P, Phartyal R, Salaria D, Kaushik N, Choi EH, et al. Changing Dynamics of SARS-CoV-2: A Global Challenge. Applied Sciences. 2022; 12(11):5546. https://doi.org/10.3390/app12115546
Chicago/Turabian StyleChugh, Ananya, Nimisha Khurana, Kangna Verma, Ishita Sehgal, Rajan Rolta, Pranjal Vats, Rajendra Phartyal, Deeksha Salaria, Neha Kaushik, Eun Ha Choi, and et al. 2022. "Changing Dynamics of SARS-CoV-2: A Global Challenge" Applied Sciences 12, no. 11: 5546. https://doi.org/10.3390/app12115546