
Report of One-Year Prospective Surveillance of SARS-CoV-2 in Dogs and Cats in France with Various Exposure Risks: Confirmation of a Low Prevalence of Shedding, Detection and Complete Sequencing of an Alpha Variant in a Cat

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samplings
2.2. Viral RNA Extraction
2.3. SARS-CoV-2 RNA Detection Using qRT-PCR
2.4. Sequencing
2.5. Amplification of Genomic Material
2.6. ONT Library Preparation and MinION Sequencing
2.7. Genome Assembly and Phylogenetic Inference
2.8. Serological Testing
2.8.1. Microsphere Immunoassay
2.8.2. Neutralization Activity Measurement
3. Results
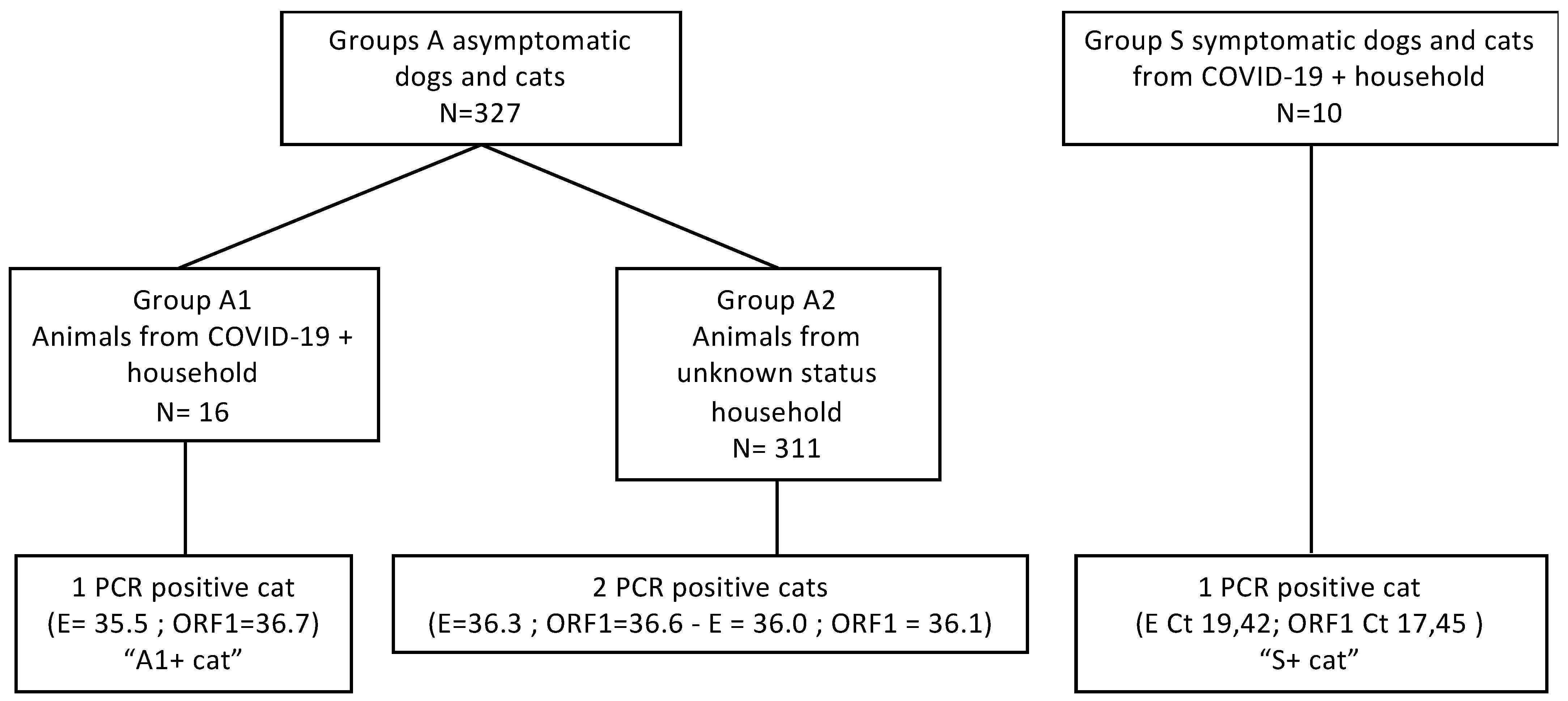
3.1. Prospective Survey
3.2. A1+ Cat
3.3. S+ Cat
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Dedication
References
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS–coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Martina, B.E.E.; Haagmans, B.L.; Kuiken, T.; Fouchier, R.; Rimmelzwaan, G.F.; Van Amerongen, G.; Peiris, J.S.M.; Lim, W.; Osterhaus, A.D.M.E. SARS virus infection of cats and ferrets. Nature 2003, 425, 915. [Google Scholar] [CrossRef]
- USDA APHIS. Cases of SARS-CoV-2 in Animals in the United States. Available online: https://www.aphis.usda.gov/aphis/dashboards/tableau/sars-dashboard (accessed on 27 July 2021).
- Fritz, M.; Rosolen, B.; Krafft, E.; Becquart, P.; Elguero, E.; Vratskikh, O.; Denolly, S.; Boson, B.; Vanhomwegen, J.; Gouilh, M.A.; et al. High prevalence of SARS-CoV-2 antibodies in pets from COVID-19+ households. One Health 2020, 11, 100192. [Google Scholar] [CrossRef]
- Patterson, E.I.; Elia, G.; Grassi, A.; Giordano, A.; Desario, C.; Medardo, M.; Smith, S.L.; Anderson, E.R.; Prince, T.; Patterson, G.T.; et al. Evidence of exposure to SARS-CoV-2 in cats and dogs from households in Italy. Nat. Commun. 2020, 11, 1–5. [Google Scholar] [CrossRef]
- Hamer, S.; Pauvolid-Corrêa, A.; Zecca, I.; Davila, E.; Auckland, L.; Roundy, C.; Tang, W.; Torchetti, M.; Killian, M.; Jenkins-Moore, M.; et al. SARS-CoV-2 Infections and viral isolations among serially tested cats and dogs in households with infected owners in Texas, USA. Viruses 2021, 13, 938. [Google Scholar] [CrossRef] [PubMed]
- Neira, V.; Brito, B.; Agüero, B.; Berrios, F.; Valdés, V.; Gutierrez, A.; Ariyama, N.; Espinoza, P.; Retamal, P.; Holmes, E.C.; et al. A household case evidences shorter shedding of SARS-CoV-2 in naturally infected cats compared to their human owners. Emerg. Microbes Infect. 2021, 10, 376–383. [Google Scholar] [CrossRef]
- Pagani, G.; Lai, A.; Bergna, A.; Rizzo, A.; Stranieri, A.; Giordano, A.; Paltrinieri, S.; Lelli, D.; Decaro, N.; Rusconi, S.; et al. Human-to-cat SARS-CoV-2 transmission: Case report and full-genome sequencing from an infected pet and its owner in Northern Italy. Pathogens 2021, 10, 252. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.-I.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; et al. Transmission of SARS-CoV-2 in domestic cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Homepage—FEDIAF. Available online: https://fediaf.org/ (accessed on 6 August 2021).
- Zhao, S.; Schuurman, N.; Li, W.; Wang, C.; Smit, L.A.; Broens, E.M.; Wagenaar, J.A.; van Kuppeveld, F.J.; Bosch, B.-J.; Egberink, H. Serologic screening of severe acute respiratory syndrome coronavirus 2 infection in cats and dogs during first coronavirus disease wave, The Netherlands. Emerg. Infect. Dis. 2021, 27, 1362–1370. [Google Scholar] [CrossRef]
- Stevanovic, V.; Tabain, I.; Vilibic-Cavlek, T.; Maljkovic, M.M.; Benvin, I.; Hruskar, Z.; Kovac, S.; Smit, I.; Miletic, G.; Hadina, S.; et al. The emergence of SARS-CoV-2 within the dog population in croatia: Host factors and clinical outcome. Viruses 2021, 13, 1430. [Google Scholar] [CrossRef]
- Michelitsch, A.; Schön, J.; Hoffmann, D.; Beer, M.; Wernike, K. The second wave of SARS-CoV-2 circulation—Antibody detection in the domestic cat population in germany. Viruses 2021, 13, 1009. [Google Scholar] [CrossRef] [PubMed]
- Calvet, G.A.; Pereira, S.A.; Ogrzewalska, M.; Pauvolid-Corrêa, A.; Resende, P.C.; Tassinari, W.D.S.; Costa, A.D.P.; Keidel, L.O.; da Rocha, A.S.B.; da Silva, M.F.B.; et al. Investigation of SARS-CoV-2 infection in dogs and cats of humans diagnosed with COVID-19 in Rio de Janeiro, Brazil. PLoS ONE 2021, 16, e0250853. [Google Scholar] [CrossRef] [PubMed]
- Dias, H.G.; Resck, M.E.B.; Caldas, G.C.; Resck, A.F.; da Silva, N.V.; dos Santos, A.M.V.; Sousa, T.d.C.; Ogrzewalska, M.H.; Siqueira, M.M.; Pauvolid-Corrêa, A.; et al. Neutralizing antibodies for SARS-CoV-2 in stray animals from Rio de Janeiro, Brazil. PLoS ONE 2021, 16, e0248578. [Google Scholar] [CrossRef]
- Spada, E.; Vitale, F.; Bruno, F.; Castelli, G.; Reale, S.; Perego, R.; Baggiani, L.; Proverbio, D. A pre- and during pandemic survey of Sars-Cov-2 infection in stray colony and shelter cats from a high endemic area of Northern Italy. Viruses 2021, 13, 618. [Google Scholar] [CrossRef] [PubMed]
- Dileepan, M.; Di, D.; Huang, Q.; Ahmed, S.; Heinrich, D.; Ly, H.; Liang, Y. Seroprevalence of SARS-CoV-2 (COVID-19) exposure in pet cats and dogs in Minnesota, USA. Virulence 2021, 12, 1597–1609. [Google Scholar] [CrossRef]
- Jara, L.M. Evidence of neutralizing antibodies against SARS-CoV-2 in domestic cats living with owners with a history of COVID-19 in Lima—Peru. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, H.; Gao, J.; Huang, K.; Yang, Y.; Hui, X.; He, X.; Li, C.; Gong, W.; Zhang, Y.; et al. A serological survey of SARS-CoV-2 in cat in Wuhan. Emerg. Microbes Infect. 2020, 9, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/emergencies/emergency-health-kits/trauma-emergency-surgery-kit-who-tesk-2019/tracking-SARS-CoV-2-variants (accessed on 3 August 2021).
- Ferasin, L.; Fritz, M.; Ferasin, H.; Becquart, P.; Legros, V.; Leroy, E.M. Myocarditis in naturally infected pets with the British variant of COVID-19. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with mul-tiplex MinION sequencing. Microb Genom. 2017, 3, e000132. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-tree 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Michael, H.T.; Waterhouse, T.; Estrada, M.; Seguin, M.A. Frequency of respiratory pathogens and SARS-CoV -2 in canine and feline samples submitted for respiratory testing in early 2020. J. Small Anim. Pr. 2021, 62, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, R.; Beltramo, C.; Magris, G.; Peletto, S.; Acutis, P.; Bozzetta, E.; Radovic, S.; Zappulla, F.; Porzio, A.M.; Gennero, M.S.; et al. First detection of an Italian human-to-cat outbreak of SARS-CoV-2 Alpha variant—lineage B.1.1.7. One Health 2021, 13, 100295. [Google Scholar] [CrossRef]
- Garigliany, M.; Van Laere, A.-S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; Van Der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 natural transmission from human to cat, belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Ruiz-Arrondo, I.; Portillo, A.; Palomar, A.M.; Santibáñez, S.; Santibáñez, P.; Cervera, C.; Oteo, J.A. Detection of SARS-CoV-2 in pets living with COVID-19 owners diagnosed during the COVID-19 lockdown in Spain: A case of an asymptomatic cat with SARS-CoV-2 in Europe. Transbound. Emerg. Dis. 2020, 68, 973–976. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Cheng, C.-Y.; Chen, C.-P.; Cheng, S.-H.; Chang, S.-Y.; Hsueh, P.-R. A case of transient existence of SARS-CoV-2 RNA in the respiratory tract with the absence of anti-SARS-CoV-2 antibody response. Int. J. Infect. Dis. 2020, 96, 464–466. [Google Scholar] [CrossRef]
- Alonso, F.D.O.M.; Sabino, B.D.; Guimarães, M.A.A.M.; Varella, R.B. Recurrence of SARS-CoV-2 infection with a more severe case after mild COVID-19, reversion of RT-qPCR for positive and late antibody response: Case report. J. Med. Virol. 2020, 93, 655–656. [Google Scholar] [CrossRef]
- Bessière, P.; Fusade-Boyer, M.; Walch, M.; Lèbre, L.; Brun, J.; Croville, G.; Boullier, S.; Cadiergues, M.-C.; Guérin, J.-L. Household cases suggest that cats belonging to owners with COVID-19 have a limited role in virus transmission. Viruses 2021, 13, 673. [Google Scholar] [CrossRef]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G. Assessing transmissibility of SARS-CoV-2 lineage B.1.1.7 in England. Nature 2021, 593, 266–269. [Google Scholar] [CrossRef]
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.B.; Russell, T.W.; Tully, D.C.; Washburne, A.D.; et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef] [PubMed]
- Hamer, S.A.; Ghai, R.R.; Zecca, I.B.; Auckland, L.D.; Roundy, C.M.; Davila, E.; Busselman, R.E.; Tang, W.; Pauvolid-Corrêa, A.; Killian, M.L.; et al. SARS-CoV-2 B.1.1.7 variant of concern detected in a pet dog and cat after exposure to a person with COVID-19, USA. Transbound. Emerg. Dis. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Arévalo, S.; Rivera, B.; Domínguez, L.; Sánchez-Vizcaíno, J. First detection of SARS-CoV-2 B.1.1.7 variant of concern in an asymptomatic dog in spain. Viruses 2021, 13, 1379. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Days Post Detection | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Test | Sample | Target | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 20 | |
| Cat A1+ | qRT-PCR | Oro-pharyngeal | E | 35.5 | 36 | - | - | - | 36 | - | - | - | - | - | - | ||
| ORF1 | 36.7 | 36 | - | - | - | 36 | - | - | - | - | - | - | |||||
| Rectal | E | - | - | - | - | - | - | - | - | - | - | - | - | ||||
| ORF1 | - | - | - | - | - | - | - | - | - | - | - | - | |||||
| Serology | Serum | N | - | - | - | ||||||||||||
| RBD | - | - | - | ||||||||||||||
| S tri | - | - | - | ||||||||||||||
| SNT | Serum | SARS-CoV-2pp | - | - | - | ||||||||||||
| Cat S+ | qRT-PCR | Oro-pharyngeal | E | 19.4 | - | ||||||||||||
| ORF1 | 17.4 | - | |||||||||||||||
| Rectal | E | 36.8 | 39.7 | ||||||||||||||
| ORF1 | 36.8 | - | |||||||||||||||
| Serology | Serum | N | - | + | |||||||||||||
| RBD | - | + | |||||||||||||||
| S tri | - | + | |||||||||||||||
| SNT | Serum | SARS-CoV-2pp | - | + | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krafft, E.; Denolly, S.; Boson, B.; Angelloz-Pessey, S.; Levaltier, S.; Nesi, N.; Corbet, S.; Leterrier, B.; Fritz, M.; Leroy, E.M.; et al. Report of One-Year Prospective Surveillance of SARS-CoV-2 in Dogs and Cats in France with Various Exposure Risks: Confirmation of a Low Prevalence of Shedding, Detection and Complete Sequencing of an Alpha Variant in a Cat. Viruses 2021, 13, 1759. https://doi.org/10.3390/v13091759
Krafft E, Denolly S, Boson B, Angelloz-Pessey S, Levaltier S, Nesi N, Corbet S, Leterrier B, Fritz M, Leroy EM, et al. Report of One-Year Prospective Surveillance of SARS-CoV-2 in Dogs and Cats in France with Various Exposure Risks: Confirmation of a Low Prevalence of Shedding, Detection and Complete Sequencing of an Alpha Variant in a Cat. Viruses. 2021; 13(9):1759. https://doi.org/10.3390/v13091759
Chicago/Turabian StyleKrafft, Emilie, Solène Denolly, Bertrand Boson, Sophie Angelloz-Pessey, Sophie Levaltier, Nicolas Nesi, Sandrine Corbet, Bryce Leterrier, Matthieu Fritz, Eric M. Leroy, and et al. 2021. "Report of One-Year Prospective Surveillance of SARS-CoV-2 in Dogs and Cats in France with Various Exposure Risks: Confirmation of a Low Prevalence of Shedding, Detection and Complete Sequencing of an Alpha Variant in a Cat" Viruses 13, no. 9: 1759. https://doi.org/10.3390/v13091759