
Morphological Alterations and Stress Protein Variations in Lung Biopsies Obtained from Autopsies of COVID-19 Subjects

 ,
,  , , ,
, , ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Histological Analysis
2.3. Immunohistochemistry
2.4. Immunofluorescence
2.5. Statistical Analyses
3. Results
3.1. Histological Analysis
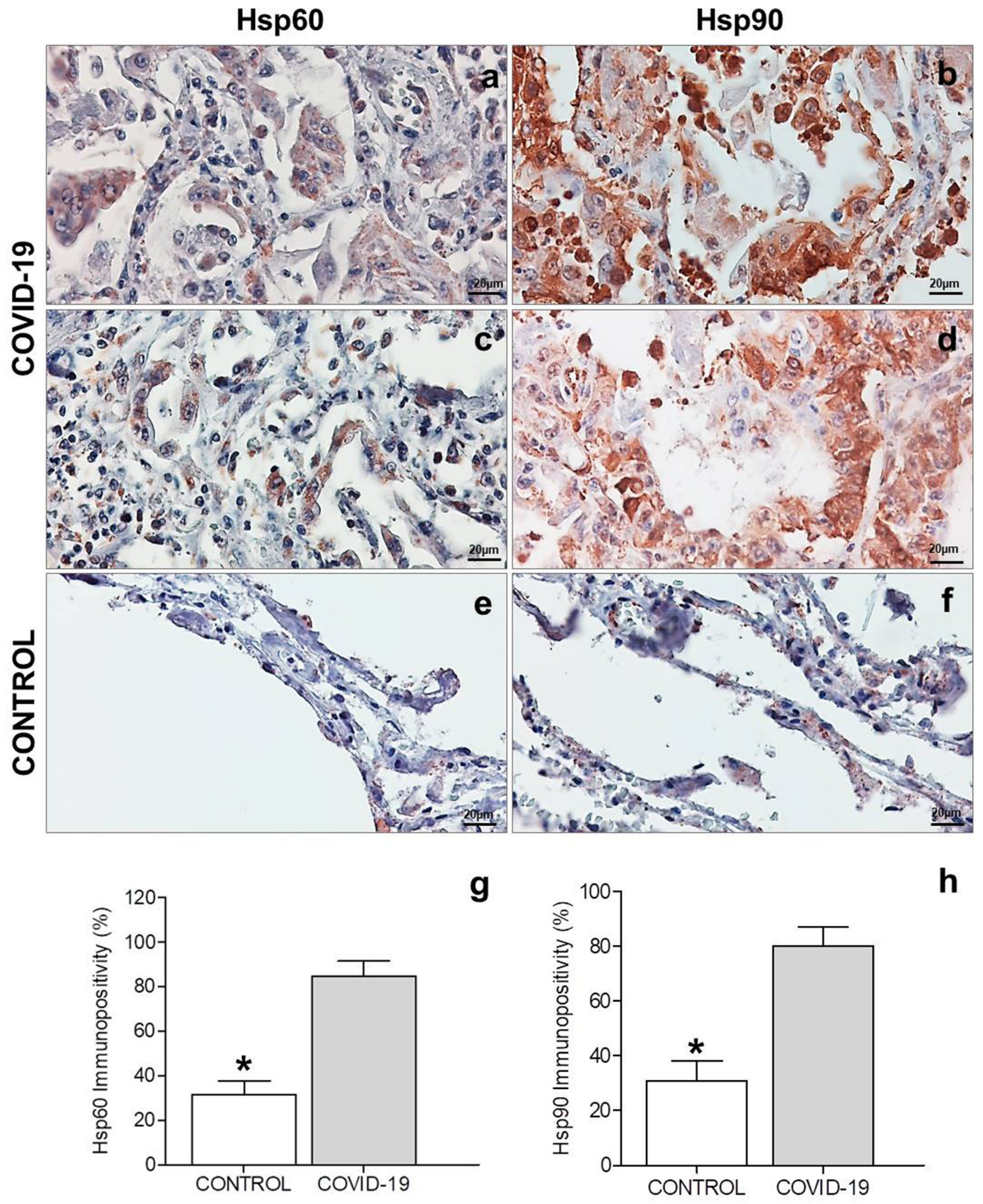
3.2. Immunohistochemistry
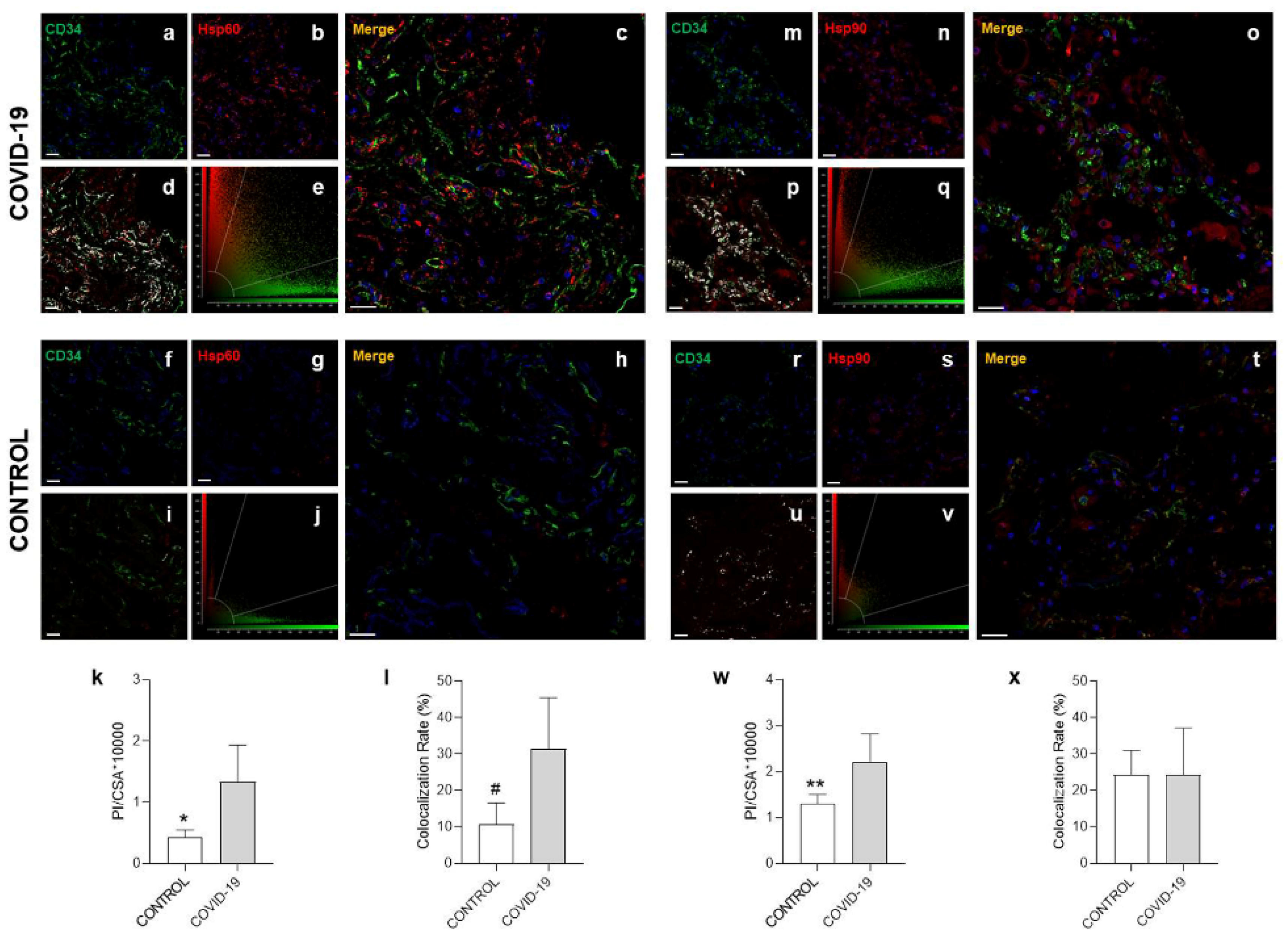
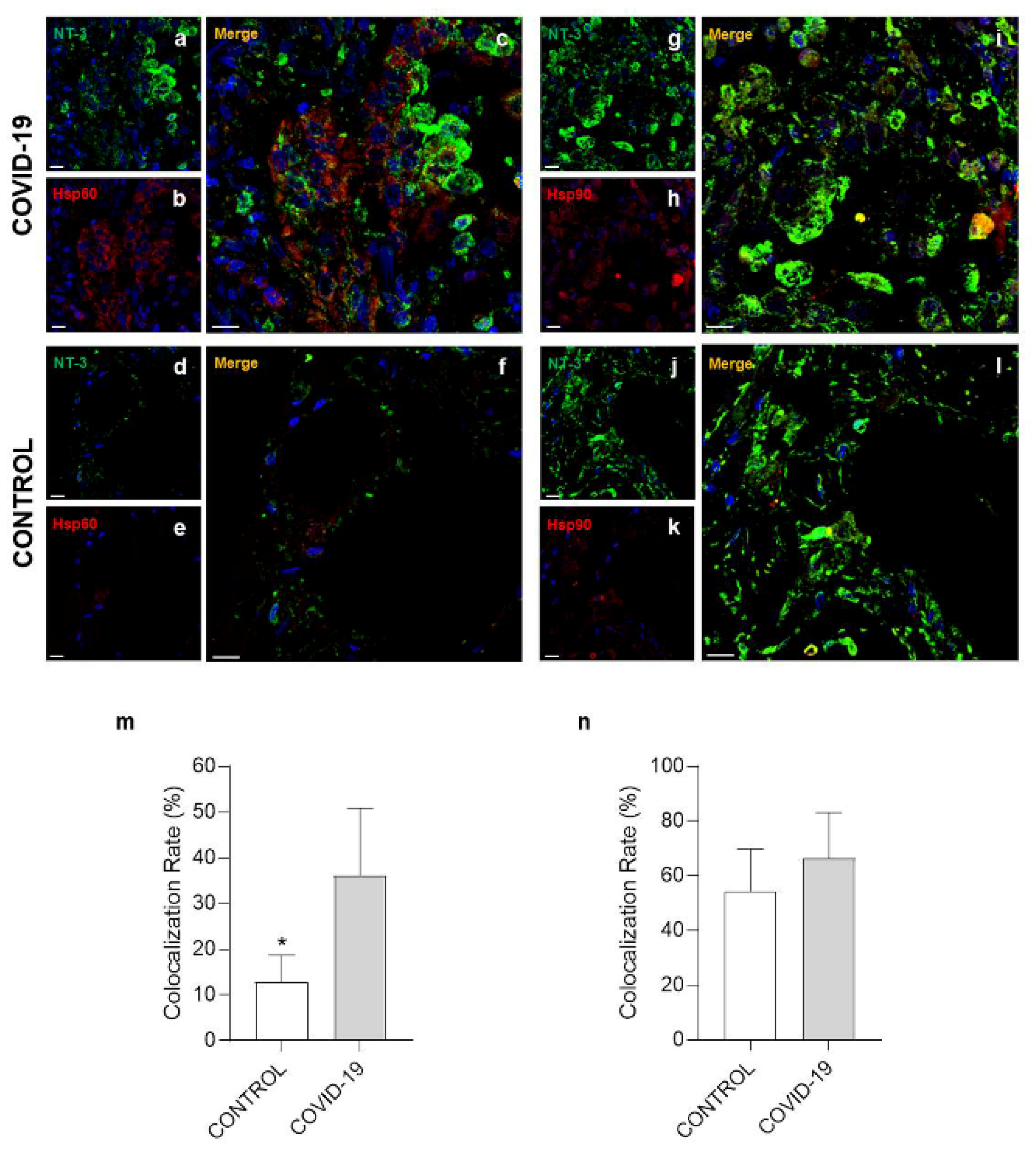
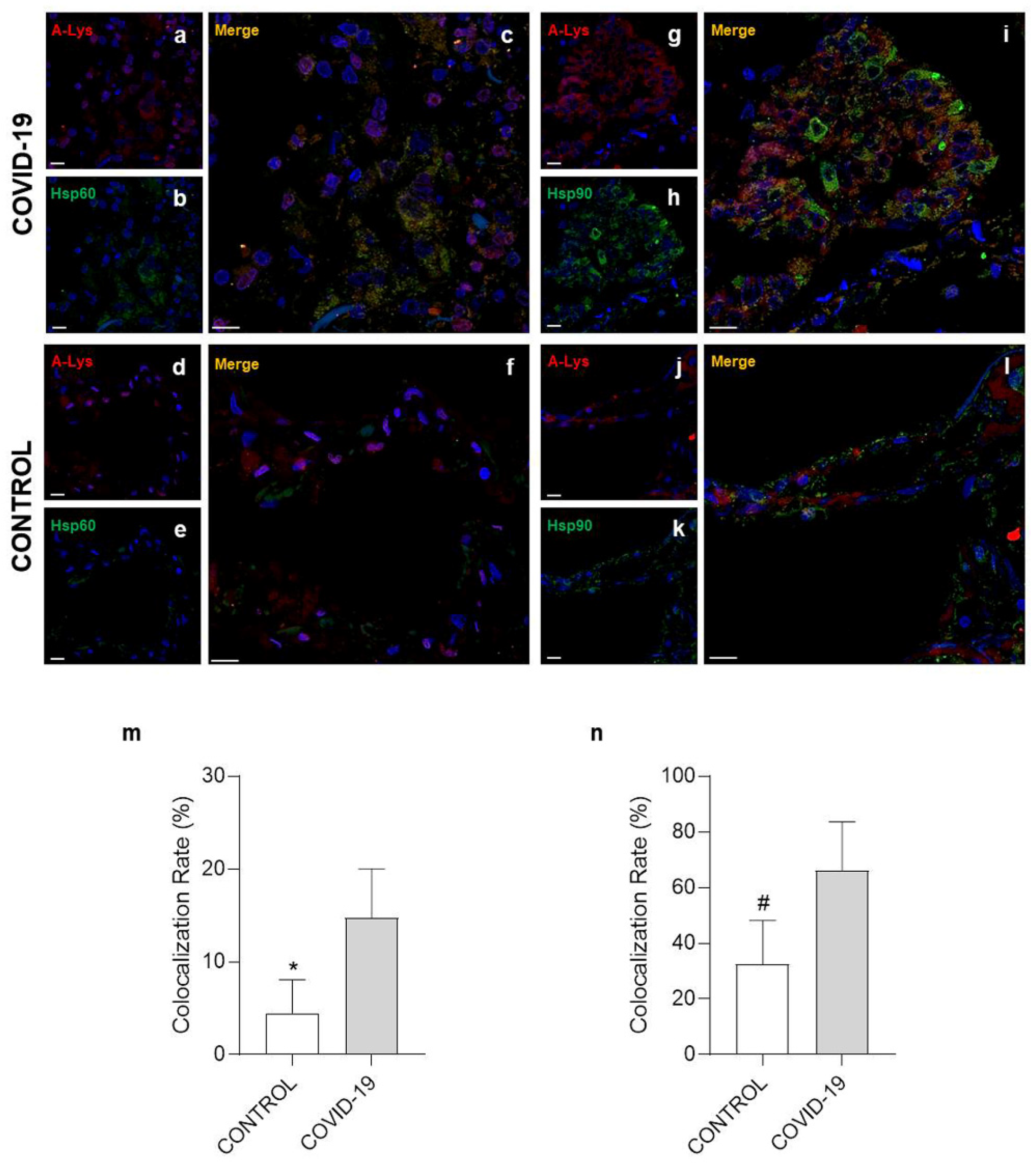
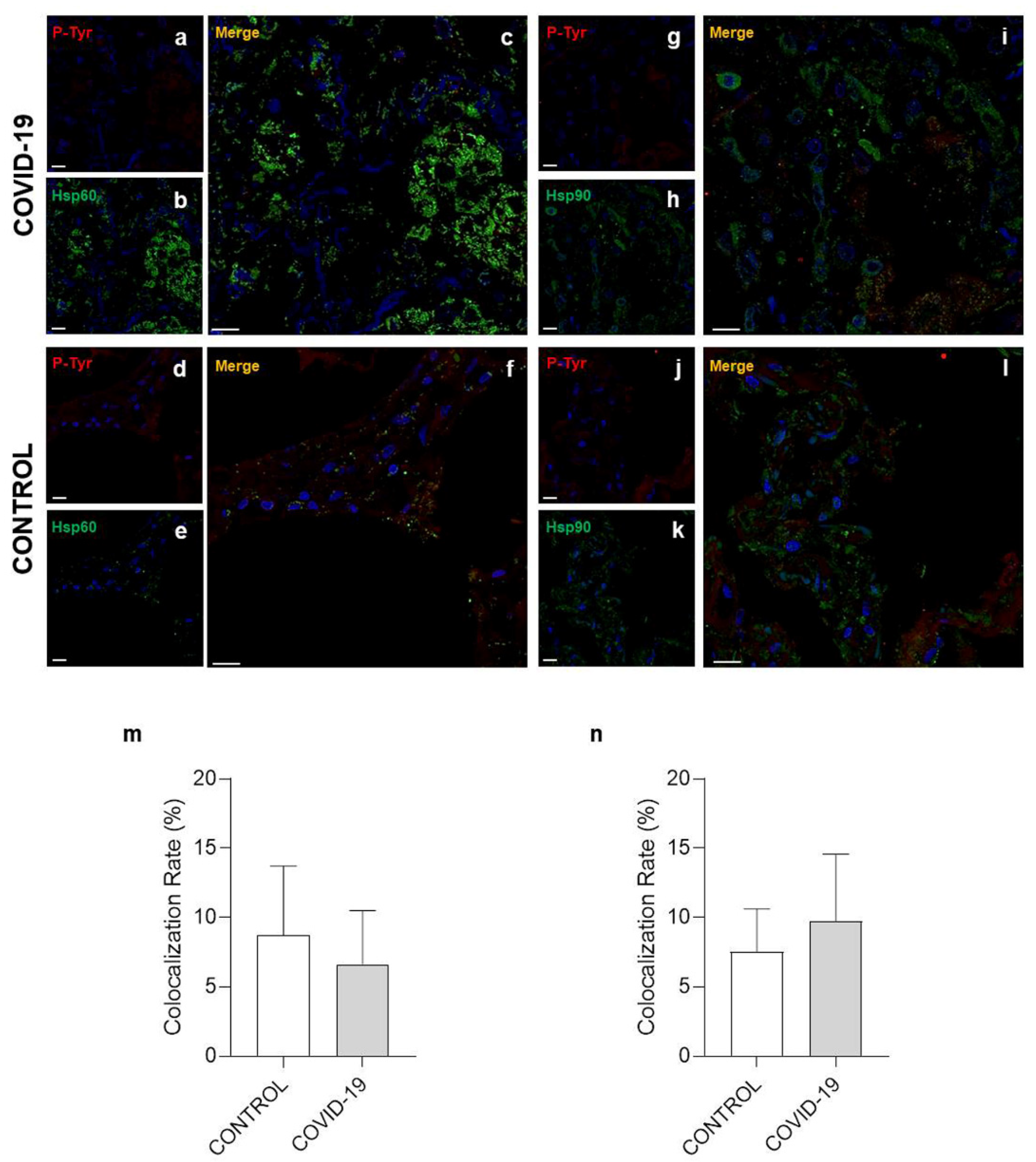
3.3. Confocal Microscopy Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bchetnia, M.; Girard, C.; Duchaine, C.; Laprise, C. The outbreak of the novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2): A review of the current global status. J. Infect. Public Health 2020, 13, 1601–1610. [Google Scholar] [CrossRef]
- Cappello, F.; Marino Gammazza, A.; Dieli, F.; de Macario, E.C.; Macario, A.J. Does SARS-CoV-2 Trigger Stress-Induced Autoimmunity by Molecular Mimicry? A Hypothesis. J. Clin. Med. 2020, 9, 2038. [Google Scholar] [CrossRef]
- Pellicori, P.; Doolub, G.; Wong, C.M.; Lee, K.S.; Mangion, K.; Ahmad, M.; Berry, C.; Squire, I.; Lambiase, P.D.; Lyon, A.; et al. COVID-19 and its cardiovascular effects: A systematic review of prevalence studies. Cochrane Database Syst. Rev. 2021, 2021, CD013879. [Google Scholar] [CrossRef]
- Ejaz, H.; Alsrhani, A.; Zafar, A.; Javed, H.; Junaid, K.; Abdalla, A.E.; Abosalif, K.O.; Ahmed, Z.; Younas, S. COVID-19 and comorbidities: Deleterious impact on infected patients. J. Infect. Public Health 2020, 13, 1833–1839. [Google Scholar] [CrossRef] [PubMed]
- Tadic, M.; Saeed, S.; Grassi, G.; Taddei, S.; Mancia, G.; Cuspidi, C. Hypertension and COVID-19: Ongoing Controversies. Front. Cardiovasc. Med. 2021, 8, 639222. [Google Scholar] [CrossRef]
- Rastogi, A.; Gupta, S.K.; Lakshmi, P.V.M.; Kaur, M. Role of self-care in COVID-19 pandemic for people living with comorbidities of diabetes and hypertension. J. Fam. Med. Prim. Care 2020, 9, 5495–5501. [Google Scholar] [CrossRef] [PubMed]
- Aghili, S.M.M.; Ebrahimpur, M.; Arjmand, B.; Shadman, Z.; Sani, M.P.; Qorbani, M.; Larijani, B.; Payab, M. Obesity in COVID-19 era, implications for mechanisms, comorbidities, and prognosis: A review and meta-analysis. Int. J. Obes. 2021, 45, 998–1016. [Google Scholar] [CrossRef]
- Xu, B.-J.; Chen, J.; Chen, X.; Liu, X.-W.; Fang, S.; Shu, Q.; Hu, L.; Shi, S.-S.; Du, L.-Z.; Tan, L.-H. High shear stress-induced pulmonary hypertension alleviated by endothelial progenitor cells independent of autophagy. World J. Pediatr. 2015, 11, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Salibe-Filho, W.; Araujo, T.L.S.; Melo, E.G.; Coimbra, L.B.C.T.; Lapa, M.S.; Acencio, M.M.P.; Freitas-Filho, O.; Capelozzi, V.L.; Teixeira, L.R.; Fernandes, C.J.C.S.; et al. Shear stress-exposed pulmonary artery endothelial cells fail to upregulate HSP70 in chronic thromboembolic pulmonary hypertension. PLoS ONE 2020, 15, e0242960. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Orekhov, A.N.; Bobryshev, Y.V. Endothelial Barrier and Its Abnormalities in Cardiovascular Disease. Front. Physiol. 2015, 6, 365. [Google Scholar] [CrossRef]
- Esper, R.J.; Vilariño, J.O.; Machado, R.A.; Paragano, A. Endothelial Dysfunction in Normal and Abnormal Glucose Metabolism. Thromb. Coron. Heart Dis. 2008, 45, 17–43. [Google Scholar] [CrossRef]
- John, A.; Kizhakkedath, P.; Al-Gazali, L.; Ali, B.R. Defective cellular trafficking of the bone morphogenetic protein receptor type II by mutations underlying familial pulmonary arterial hypertension. Gene 2015, 561, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Erwin, P.A.; Mitchell, D.; Sartoretto, J.; Marletta, M.A.; Michel, T. Subcellular Targeting and Differential S-Nitrosylation of Endothelial Nitric-oxide Synthase. J. Biol. Chem. 2006, 281, 151–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz, P.; Garvin, J.L. Trafficking and activation of eNOS in epithelial cells. Acta Physiol. Scand. 2003, 179, 107–114. [Google Scholar] [CrossRef]
- Bavisotto, C.C.; Alberti, G.; Vitale, A.M.; Paladino, L.; Campanella, C.; Rappa, F.; Gorska, M.; De Macario, E.C.; Cappello, F.; Macario, A.J.L.; et al. Hsp60 Post-translational Modifications: Functional and Pathological Consequences. Front. Mol. Biosci. 2020, 7, 95. [Google Scholar] [CrossRef]
- Chiu, W.-T.; Shen, S.-C.; Yang, L.-Y.; Chow, J.-M.; Wu, C.-Y.; Chen, Y.-C. Inhibition of HSP90-dependent telomerase activity in amyloid β-induced apoptosis of cerebral endothelial cells. J. Cell. Physiol. 2011, 226, 2041–2051. [Google Scholar] [CrossRef]
- Wick, G.; Jakic, B.; Buszko, M.; Wick, M.C.; Grundtman, C. The role of heat shock proteins in atherosclerosis. Nat. Rev. Cardiol. 2014, 11, 516–529. [Google Scholar] [CrossRef]
- Kreutmayer, S.B.; Messner, B.; Knoflach, M.; Henderson, B.; Niederegger, H.; Böck, G.; van der Zee, R.; Wick, G.; Bernhard, D. Dynamics of heat shock protein 60 in endothelial cells exposed to cigarette smoke extract. J. Mol. Cell. Cardiol. 2011, 51, 777–780. [Google Scholar] [CrossRef] [Green Version]
- Wick, C. Tolerization against atherosclerosis using heat shock protein 60. Cell Stress Chaperon 2015, 21, 201–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, F. Is COVID-19 a proteiform disease inducing also molecular mimicry phenomena? Cell Stress Chaperon 2020, 25, 381–382. [Google Scholar] [CrossRef]
- Pomara, C.; Volti, G.L.; Cappello, F. COVID-19 Deaths: Are We Sure It Is Pneumonia? Please, Autopsy, Autopsy, Autopsy! J. Clin. Med. 2020, 9, 1259. [Google Scholar] [CrossRef] [PubMed]
- Angileri, F.; Legare, S.; Gammazza, A.M.; de Macario, E.C.; Macario, A.J.; Cappello, F. Molecular mimicry may explain multi-organ damage in COVID-19. Autoimmun. Rev. 2020, 19, 102591. [Google Scholar] [CrossRef] [PubMed]
- Angileri, F.; Légaré, S.; Gammazza, A.M.; De Macario, E.C.; Macario, A.J.L.; Cappello, F. Is molecular mimicry the culprit in the autoimmune haemolytic anaemia affecting patients with COVID-19? Br. J. Haematol. 2020, 190, 92. [Google Scholar] [CrossRef]
- Cappello, F. COVID-19 and molecular mimicry: The Columbus’ egg? J. Clin. Neurosci. 2020, 77, 246. [Google Scholar] [CrossRef]
- Marino, G.A.; Légaré, S.; Lo Bosco, G.; Fucarino, A.; Angileri, F.; Conway de Macario, E.; Macario, A.J.; Cappello, F. Human molecular chaperones share with SARS-CoV-2 antigenic epitopes potentially capable of eliciting autoimmunity against endothelial cells: Possible role of molecular mimicry in COVID-19. Cell Stress Chaperones 2020, 25, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Pomara, C.; Volti, G.L.; Cappello, F. The Post-Lockdown Era: What Is Next in Italy? Front. Pharmacol. 2020, 11, 1074. [Google Scholar] [CrossRef]
- Pomara, C.; Salerno, M.; Sessa, F.; Esposito, M.; Barchitta, M.; Ledda, C.; Grassi, P.; Liberto, A.; Mattaliano, A.; Rapisarda, V.; et al. Safe Management Strategies in Clinical Forensic Autopsies of Confirmed COVID-19 Cases. Diagnostics 2021, 11, 457. [Google Scholar] [CrossRef]
- Pomara, C.; Fineschi, V. Forensic and Clinical Forensic Autopsy. In An Atlas and Handbook 2020, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Barone, R.; Macaluso, F.; Sangiorgi, C.; Campanella, C.; Gammazza, A.M.; Moresi, V.; Coletti, D.; De Macario, E.C.; Macario, A.J.; Cappello, F.; et al. Skeletal muscle Heat shock protein 60 increases after endurance training and induces peroxisome proliferator-activated receptor gamma coactivator 1 α1 expression. Sci. Rep. 2016, 6, 19781. [Google Scholar] [CrossRef] [Green Version]
- Santos, A.L.; Lindner, A.B. Protein Posttranslational Modifications: Roles in Aging and Age-Related Disease. Oxid. Med. Cell. Longev. 2017, 2017, 5716409. [Google Scholar] [CrossRef] [Green Version]
- Bösmüller, H.; Matter, M.; Fend, F.; Tzankov, A. The pulmonary pathology of COVID-19. Virchows Arch. 2021, 478, 137–150. [Google Scholar] [CrossRef]
- Satturwar, S.; Fowkes, M.; Farver, C.; Wilson, A.M.; Eccher, A.; Girolami, I.; Pujadas, E.; Bryce, C.; Salem, F.; El Jamal, S.M.; et al. Postmortem Findings Associated With SARS-CoV-2: Systematic Review and Meta-analysis. Am. J. Surg. Pathol. 2021, 45, 587–603. [Google Scholar] [CrossRef]
- Bradley, B.T.; Maioli, H.; Johnston, R.; Chaudhry, I.; Fink, S.L.; Xu, H.; Najafian, B.; Deutsch, G.; Lacy, J.M.; Williams, T.; et al. Histopathology and ultrastructural findings of fatal COVID-19 infections in Washington State: A case series. Lancet 2020, 396, 320–332. [Google Scholar] [CrossRef]
- Konopka, K.E.; Nguyen, T.; Jentzen, J.M.; Rayes, O.; Schmidt, C.J.; Wilson, A.M.; Farver, C.F.; Myers, J.L. Diffuse alveolar damage (DAD) resulting from coronavirus disease 2019 Infection is Morphologically Indistinguishable from Other Causes of DAD. Histopathology 2020, 77, 570–578. [Google Scholar] [CrossRef]
- Almanzar, G.; Öllinger, R.; Leuenberger, J.; Onestingel, E.; Rantner, B.; Zehm, S.; Cardini, B.; van der Zee, R.; Grundtman, C.; Wick, G. Autoreactive HSP60 epitope-specific T-cells in early human atherosclerotic lesions. J. Autoimmun. 2012, 39, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Grundtman, C.; Wick, G. The autoimmune concept of atherosclerosis. Curr. Opin. Lipidol. 2011, 22, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Grundtman, C.; Kreutmayer, S.B.; Almanzar, G.; Wick, M.C.; Wick, G. Heat Shock Protein 60 and Immune Inflammatory Responses in Atherosclerosis. Arter. Thromb. Vasc. Biol. 2011, 31, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucchese, G.; Flöel, A. SARS-CoV-2 and Guillain-Barré syndrome: Molecular mimicry with human heat shock proteins as potential pathogenic mechanism. Cell Stress Chaperones 2020, 25, 731–735. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Vázquez, E.; de Castro, P. “2-6-11” motif in heat shock protein 60 and central nervous system antigens: A preliminary study in multiple sclerosis patients. J. Physiol. Biochem. 2003, 59, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nittari, G.; Pallotta, G.; Amenta, F.; Tayebati, S.K. Current pharmacological treatments for SARS-CoV-2: A narrative review. Eur. J. Pharmacol. 2020, 882, 173328. [Google Scholar] [CrossRef]
- Hajjar, L.A.; Costa, I.B.S.D.S.; Rizk, S.I.; Biselli, B.; Gomes, B.R.; Bittar, C.S.; de Oliveira, G.Q.; de Almeida, J.P.; Bello, M.V.D.O.; Garzillo, C.; et al. Intensive care management of patients with COVID-19: A practical approach. Ann. Intensiv. Care 2021, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| COVID-19 | |||||||
| Case | Age | Sex | Days in Hospital | Thorax Radiological Findings | Clinical Personal History | Intubation | Initial Clinical Presentation |
| 1 | 61 | F | 23 | Ground glass opacities | Hypertension, major depressive disorder | Yes | Confusional state, anemia |
| 2 | 79 | F | 19 | Ground glass opacities | Cardiovascular disease, COPD, chronic kidney disease | Yes | Fever, anemia, dyspnea/tachypnea |
| 3 | 72 | F | 10 | Hypertension, cardiovascular disease, COPD, smoker, diabetes mellitus, dyslipidemia, hypothyroidism | Yes | Cardiac arrest | |
| 4 | 68 | F | 2 | Ground glass opacities | Diabetes mellitus, rheumatoid arthritis, polymyalgia | Yes | Fever, dyspnea/tachypnea, diarrhea, anemia |
| 5 | 70 | M | 1, Before admission in hospital for emergency, medical consultation. | No | Fever; asthenia, respiratory failure, disorientation | ||
| 6 | 42 | M | 0, Medical consultation, no hospitalization | Alcoholism, pancreatitis | No | Fever, asthenia, respiratory failure, disorientation | |
| Control | |||||||
| Case | Age | Sex | Cause of Death | ||||
| 1 | 46 | F | Death in a road accident | ||||
| 2 | 68 | M | Death by firearm | ||||
| 3 | 66 | F | Death from brain hemorrhage | ||||
| 4 | 54 | F | Death by suicide | ||||
| 5 | 70 | M | Death by suicide | ||||
| 6 | 72 | M | Death in a road accident | ||||
| Method | Antigen | Type and Source | Catalog Number | Supplier | Dilution |
|---|---|---|---|---|---|
| IHC | CKAE1AE3 | Mouse monoclonal | CM 011 A,B,C | BIOCARE medical | 1:100 |
| IHC | CK7 | Mouse monoclonal | CM 061 A,B,C | BIOCARE medical | 1:100 |
| IHC/ IF | CD34 | Mouse monoclonal | CM 084 A,B,C | BIOCARE medical | 1:100/ 1:50 |
| IHC | CD61 | Mouse monoclonal | ACI 3139 A,C | BIOCARE medical | 1:100 |
| IHC | CD68 | Mouse monoclonal | ACI 3139 A,C | BIOCARE medical | 1:100 |
| IHC | Ki67 | Mouse monoclonal | API 3156 AA, H | BIOCARE medical | 1:100 |
| IHC/ IF | Hsp60 | Rabbit polyclonal | sc-13966 | Santa Cruz Biotechnology | 1:300/ 1:50 |
| IHC/ IF | Hsp90 | Mouse monoclonal | sc-59577 | Santa Cruz Biotechnology | 1:100/ 1:50 |
| IF | Hsp60 | Mouse monoclonal | ab13532 | Abcam | 1:50 |
| IF | Hsp90 | Rabbit polyclonal | ab13495 | Abcam | 1:50 |
| IF | Acetylate-lisine | Rabbit polyclonal | #9441 | Cell Signaling Technology | 1:50 |
| IF | 3-nitrotyrosine | Mouse monoclonal | ab61392 | Abcam | 1:50 |
| IF | Phosphotyrosine | Rabbit polyclonal | ab179530 | Abcam | 1:50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barone, R.; Marino Gammazza, A.; Paladino, L.; Pitruzzella, A.; Spinoso, G.; Salerno, M.; Sessa, F.; Pomara, C.; Cappello, F.; Rappa, F. Morphological Alterations and Stress Protein Variations in Lung Biopsies Obtained from Autopsies of COVID-19 Subjects. Cells 2021, 10, 3136. https://doi.org/10.3390/cells10113136
Barone R, Marino Gammazza A, Paladino L, Pitruzzella A, Spinoso G, Salerno M, Sessa F, Pomara C, Cappello F, Rappa F. Morphological Alterations and Stress Protein Variations in Lung Biopsies Obtained from Autopsies of COVID-19 Subjects. Cells. 2021; 10(11):3136. https://doi.org/10.3390/cells10113136
Chicago/Turabian StyleBarone, Rosario, Antonella Marino Gammazza, Letizia Paladino, Alessandro Pitruzzella, Giulio Spinoso, Monica Salerno, Francesco Sessa, Cristoforo Pomara, Francesco Cappello, and Francesca Rappa. 2021. "Morphological Alterations and Stress Protein Variations in Lung Biopsies Obtained from Autopsies of COVID-19 Subjects" Cells 10, no. 11: 3136. https://doi.org/10.3390/cells10113136