



The Bittersweet Symphony of COVID-19: Associations between TAS1Rs and TAS2R38 Genetic Variations and COVID-19 Symptoms
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Patients Cohort and Clinical Evaluation
2.3. Sample Collection and Genetic Analysis
2.4. Statistical Analysis
3. Results
3.1. COVID-19 Patient Clinical Evaluation
3.2. TAS2R38, TAS1R2, and TAS1R3 Haplotype/Genotype Distribution
3.3. Genetic Variation within TAS1R2, TAS1R3 and COVID-19 Symptoms
3.4. Association between the TAS2R38 Haplotype and COVID-19 Symptoms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parsa, S.; Mogharab, V.; Ebrahimi, M.; Ahmadi, S.R.; Shahi, B.; Mehramiz, N.J.; Foroughian, M.; Zarenezhad, M.; Kalani, N.; Abdi, M.H.; et al. COVID-19 as a Worldwide Selective Event and Bitter Taste Receptor Polymorphisms: An Ecological Correlational Study. Int. J. Biol. Macromol. 2021, 177, 204–210. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiao, B.; Qu, L.; Yang, D.; Liu, R. The Development of COVID-19 Treatment. Front. Immunol. 2023, 14, 1125246. [Google Scholar] [CrossRef] [PubMed]
- Mair, M.; Singhavi, H.; Pai, A.; Singhavi, J.; Gandhi, P.; Conboy, P.; Baker, A.; Das, S. A Meta-Analysis of 67 Studies with Presenting Symptoms and Laboratory Tests of COVID-19 Patients. Laryngoscope 2021, 131, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Mastrangelo, A.; Bonato, M.; Cinque, P. Smell and Taste Disorders in COVID-19: From Pathogenesis to Clinical Features and Outcomes. Neurosci. Lett. 2021, 748, 135694. [Google Scholar] [CrossRef] [PubMed]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the Human Innate Immune System. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Palmer, J.N.; Adappa, N.D.; Cohen, N.A. The Role of Bitter and Sweet Taste Receptors in Upper Airway Immunity. Curr. Allergy Asthma Rep. 2015, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Cohen, N.A. Taste Receptors in Innate Immunity. Cell. Mol. Life Sci. 2015, 72, 217–236. [Google Scholar] [CrossRef]
- Liszt, K.I.; Wang, Q.; Farhadipour, M.; Segers, A.; Thijs, T.; Nys, L.; Deleus, E.; van der Schueren, B.; Gerner, C.; Neuditschko, B.; et al. Human Intestinal Bitter Taste Receptors Regulate Innate Immune Responses and Metabolic Regulators in Obesity. J. Clin. Investig. 2022, 132, e144828. [Google Scholar] [CrossRef]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human Receptors for Sweet and Umami Taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef]
- Lu, P.; Zhang, C.H.; Lifshitz, L.M.; ZhuGe, R. Extraoral Bitter Taste Receptors in Health and Disease. J. General. Physiol. 2017, 149, 181–197. [Google Scholar] [CrossRef]
- Lee, R.J.; Cohen, N.A. Bitter and Sweet Taste Receptors in the Respiratory Epithelium in Health and Disease. J. Mol. Med. 2014, 92, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Hollenhorst, M.I.; Krasteva-Christ, G. The Journal of Physiology Chemosensory Cells in the Respiratory Tract as Crucial Regulators of Innate Immune Responses. J. Physiol. 2023, 601, 1555–1572. [Google Scholar] [CrossRef] [PubMed]
- Kamila, T.; Agnieszka, K. An Update on Extra-Oral Bitter Taste Receptors. J. Transl. Med. 2021, 19, 440. [Google Scholar]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and Sweet Taste Receptors Regulate Human Upper Respiratory Innate Immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Bethineedi, L.D.; Baghsheikhi, H.; Soltani, A.; Mafi, Z.; Samieefar, N.; Seraj, S.S.; Tabari, M.A.K. Human T2R38 Bitter Taste Receptor Expression and COVID-19: From Immunity to Prognosis. Avicenna J. Med. Biotechnol. 2023, 15, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M.; Adappa, N.D.; Palmer, J.N.; Lee, R.J.; Cohen, N.A. Taste Receptors: Regulators of Sinonasal Innate Immunity. Laryngoscope Investig. Otolaryngol. 2016, 1, 88–95. [Google Scholar] [CrossRef]
- Douglas, J.E.; Cohen, N.A. Taste Receptors Mediate Sinonasal Immunity and Respiratory Disease. Int. J. Mol. Sci. 2017, 18, 437. [Google Scholar] [CrossRef]
- Åkerström, S.; Gunalan, V.; Keng, C.T.; Tan, Y.J.; Mirazimi, A. Dual Effect of Nitric Oxide on SARS-CoV Replication: Viral RNA Production and Palmitoylation of the S Protein Are Affected. Virology 2009, 395, 1–9. [Google Scholar] [CrossRef]
- Risso, D.; Carmagnola, D.; Morini, G.; Pellegrini, G.; Canciani, E.; Antinucci, M.; Henin, D.; Dellavia, C. Distribution of TAS2R38 Bitter Taste Receptor Phenotype and Haplotypes among COVID-19 Patients. Sci. Rep. 2022, 12, 7381. [Google Scholar] [CrossRef]
- Farquhar, D.R.; Kovatch, K.J.; Palmer, J.N.; Shofer, F.S.; Adappa, N.D.; Cohen, N.A. Phenylthiocarbamide Taste Sensitivity Is Associated with Sinonasal Symptoms in Healthy Adults. Int. Forum Allergy Rhinol. 2015, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Xiong, G.; Kofonow, J.M.; Chen, B.; Lysenko, A.; Jiang, P.; Abraham, V.; Doghramji, L.; Adappa, N.D.; Palmer, J.N.; et al. T2R38 Taste Receptor Polymorphisms Underlie Susceptibility to Upper Respiratory Infection. J. Clin. Investig. 2012, 122, 4145–4159. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.A. The Genetics of the Bitter Taste Receptor T2R38 in Upper Airway Innate Immunity and Implications for Chronic Rhinosinusitis. Laryngoscope 2017, 127, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Fushan, A.A.; Simons, C.T.; Slack, J.P.; Manichaikul, A.; Drayna, D. Allelic Polymorphism within the TAS1R3 Promoter Is Associated with Human Taste Sensitivity to Sucrose. Curr. Biol. 2009, 19, 1288–1293. [Google Scholar] [CrossRef] [PubMed]
- Eny, K.M.; Wolever, T.M.S.; Corey, P.N.; El-Sohemy, A. Genetic Variation in TAS1R2 (Ile191Val) Is Associated with Consumption of Sugars in Overweight and Obese Individuals in 2 Distinct Populations. Am. J. Clin. Nutr. 2010, 92, 1501–1510. [Google Scholar] [CrossRef]
- Barham, H.P.; Taha, M.A.; Broyles, S.T.; Stevenson, M.M.; Zito, B.A.; Hall, C.A. Association between Bitter Taste Receptor Phenotype and Clinical Outcomes among Patients with COVID-19. JAMA Netw. Open 2021, 4, e2111410. [Google Scholar] [CrossRef]
- Risso, D.S.; Mezzavilla, M.; Pagani, L.; Robino, A.; Morini, G.; Tofanelli, S.; Carrai, M.; Campa, D.; Barale, R.; Caradonna, F.; et al. Global Diversity in the TAS2R38 Bitter Taste Receptor: Revisiting a Classic Evolutionary PROPosal. Sci. Rep. 2016, 6, 25506. [Google Scholar] [CrossRef]
- Boscolo-Rizzo, P.; Tirelli, G.; Meloni, P.; Hopkins, C.; Madeddu, G.; De Vito, A.; Gardenal, N.; Valentinotti, R.; Tofanelli, M.; Borsetto, D.; et al. Coronavirus Disease 2019 (COVID-19)–Related Smell and Taste Impairment with Widespread Diffusion of Severe Acute Respiratory Syndrome–Coronavirus-2 (SARS-CoV-2) Omicron Variant. Int. Forum Allergy Rhinol. 2022, 12, 1273–1281. [Google Scholar] [CrossRef]
- Aabenhus, R.; Thorsen, H.; Siersma, V.; Brodersen, J. The Development and Validation of a Multidimensional Sum-Scaling Questionnaire to Measure Patient-Reported Outcomes in Acute Respiratory Tract Infections in Primary Care: The Acute Respiratory Tract Infection Questionnaire. Value Health 2013, 16, 987–992. [Google Scholar] [CrossRef]
- Van der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to High-Confidence Variant Calls: The Genome Analysis Toolkit Best Practices Pipeline. Curr. Protoc. Bioinform. 2013, 43, 11.10.1–11.10.33. [Google Scholar] [CrossRef]
- Tarasov, A.; Vilella, A.J.; Cuppen, E.; Nijman, I.J.; Prins, P. Sambamba: Fast Processing of NGS Alignment Formats. Bioinformatics 2015, 31, 2032–2034. [Google Scholar] [CrossRef] [PubMed]
- Depristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A Framework for Variation Discovery and Genotyping Using Next-Generation DNA Sequencing Data. Nat. Genet. 2011, 43, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M. Twelve Years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.; Barnes, M.; Jordan, C. Do Experiences with Nature Promote Learning? Converging Evidence of a Cause-and-Effect Relationship. Front. Psychol. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Donnelly, P. A Comparison of Bayesian Methods for Haplotype Reconstruction from Population Genotype Data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Smith, N.J.; Donnelly, P. A New Statistical Method for Haplotype Reconstruction from Population Data. Am. J. Hum. Genet. 2001, 68, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Goodman, J.R.; Dando, R. To Detect and Reject, Parallel Roles for Taste and Immunity. Curr. Nutr. Rep. 2021, 10, 137–145. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, M.; Brand, J.; Huang, L. Inflammation and Taste Disorders: Mechanisms in Taste Buds. Ann. N. Y. Acad. Sci. 2009, 1170, 596–603. [Google Scholar] [CrossRef]
- Gibson, P.G.; Qin, L.; Puah, S.H. COVID-19 Acute Respiratory Distress Syndrome (ARDS): Clinical Features and Differences from Typical Pre-COVID-19 ARDS. Med. J. Aust. 2020, 213, 54–56.e1. [Google Scholar] [CrossRef]
- Harrington, E.O.; Vang, X.A.; Braza, J.; Shil, A.; Chichger, H. Activation of the Sweet Taste Receptor, T1R3, by the Artificial Sweetener Sucralose Regulates the Pulmonary Endothelium. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, 165–176. [Google Scholar] [CrossRef]
- Douglas, J.E.; Saunders, C.J.; Reed, D.R.; Cohen, N.A. A Role for Airway Taste Receptor Modulation in the Treatment of Upper Respiratory Infections. Expert. Rev. Respir. Med. 2016, 10, 157–170. [Google Scholar] [CrossRef] [PubMed]


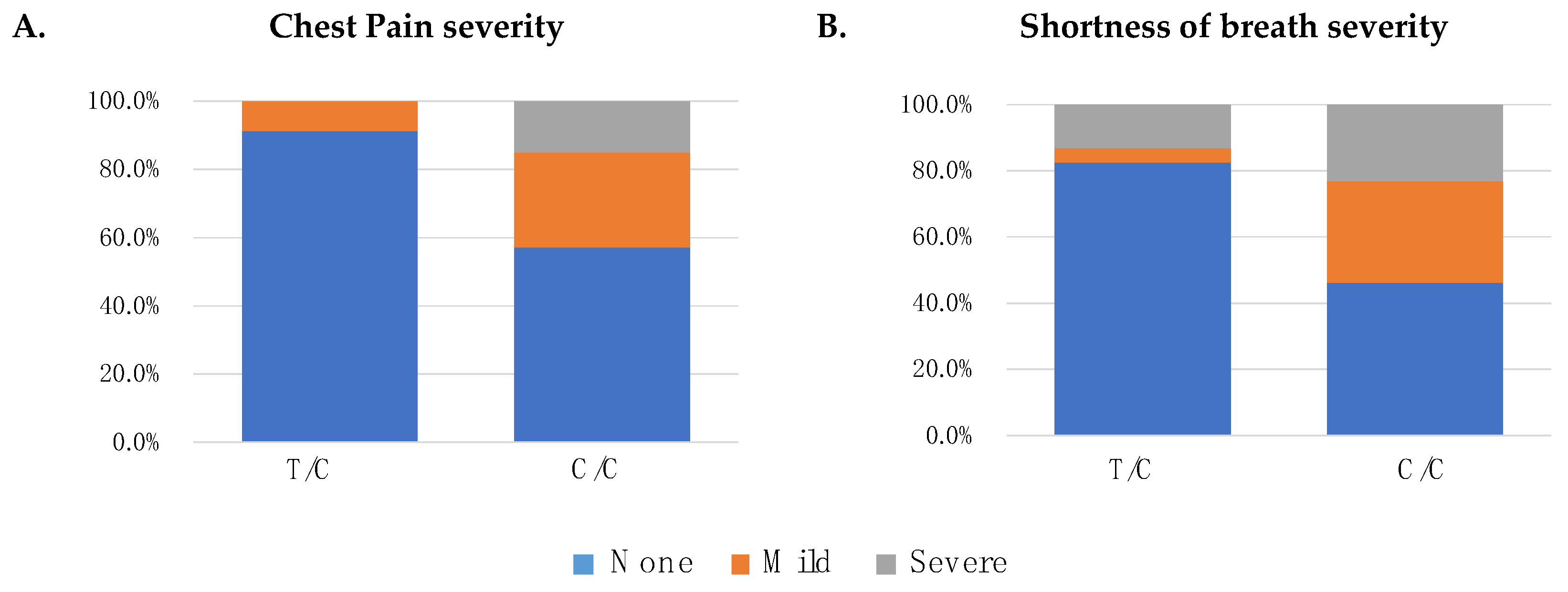{kind=link}
| Demographic and Clinical Data (n = 196) | Mean ± sd or N (%) |
|---|---|
| Age | 48.8 ± 13.8 |
| Gender | |
| Female | 137 (69.9%) |
| Male | 59 (30.1%) |
| Diabetes (n = 194) | 6 (3.1%) |
| Cardiovascular disease (n = 194) | 11 (5.7%) |
| Active cancer (n = 194) | 2 (1.0%) |
| Chronic respiratory (n = 194) | 6 (3.1%) |
| Liver disease (n = 194) | 1 (0.5%) |
| ARTIQ evaluation | |
| Dry cough | 105 (53.6%) |
| Coughing up mucus | 50 (25.5%) |
| Hearing loss | 56 (35.4%) |
| Blocked nose | 105 (53.6%) |
| Rhinorrhoea | 83 (52.9%) |
| Sneezing | 83 (52.5%) |
| Lacrimation | 61 (38.6%) |
| Hoarseness | 49 (31%) |
| Fever | 129 (65.8%) |
| Swelling | 64 (40.8%) |
| Chills | 81 (51.3%) |
| Headache | 117 (59.7%) |
| Sore throat | 83 (42.3%) |
| Muscle pain | 135 (68.9%) |
| Joint pain | 132 (67.3%) |
| Chest pain | 76 (38.8%) |
| Sinonasal pain | 54 (27.6%) |
| Swollen glands | 23 (14.6%) |
| Loss of appetite | 86 (43.9%) |
| Breathing problems | 74 (37.8%) |
| Shortness of breath | 97 (49.5%) |
| Genetic Analyses | N (%) | |
|---|---|---|
| TAS2R38 haplotype distribution | ||
| PAV/PAV | 38 (19.4%) | |
| PAV/AVI | 103 (52.6%) | |
| AVI/AVI | 40 (20.4%) | |
| Others | 15 (7.7%) | |
| AAI/PAV | 2 (1.02%) | |
| AAV/PAV | 9 (4.6%) | |
| AVI/AAV | 4 (2.04%) | |
| TAS1R3 (rs307355) genotype frequencies | ||
| T/T | 0 (0.00%) | |
| T/C | 23 (11.7%) | |
| C/C | 173 (88.3%) | |
| TAS1R2 (rs35874116) genotype frequencies | ||
| T/T | 98 (50.0) | |
| T/C | 78 (39.8) | |
| C/C | 20 (10.2) | |
| COVID-19 Symptoms | Regression Models | rs307355 [C/C Genotype] | ||
|---|---|---|---|---|
| OR | CI 95% | p-Value | ||
| Chest Pain | Logistic | 8.11 | 2.26–51.99 | 0.001 |
| Ordinal | 8.30 | 2.32–53.13 | 0.0004 | |
| Shortness of breath | Logistic | 5.45 | 1.94–19.48 | 0.001 |
| Ordinal | 4.83 | 1.71–17.32 | 0.0021 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santin, A.; Spedicati, B.; Pecori, A.; Nardone, G.G.; Concas, M.P.; Piatti, G.; Menini, A.; Tirelli, G.; Boscolo-Rizzo, P.; Girotto, G. The Bittersweet Symphony of COVID-19: Associations between TAS1Rs and TAS2R38 Genetic Variations and COVID-19 Symptoms. Life 2024, 14, 219. https://doi.org/10.3390/life14020219
Santin A, Spedicati B, Pecori A, Nardone GG, Concas MP, Piatti G, Menini A, Tirelli G, Boscolo-Rizzo P, Girotto G. The Bittersweet Symphony of COVID-19: Associations between TAS1Rs and TAS2R38 Genetic Variations and COVID-19 Symptoms. Life. 2024; 14(2):219. https://doi.org/10.3390/life14020219
Chicago/Turabian StyleSantin, Aurora, Beatrice Spedicati, Alessandro Pecori, Giuseppe Giovanni Nardone, Maria Pina Concas, Gioia Piatti, Anna Menini, Giancarlo Tirelli, Paolo Boscolo-Rizzo, and Giorgia Girotto. 2024. "The Bittersweet Symphony of COVID-19: Associations between TAS1Rs and TAS2R38 Genetic Variations and COVID-19 Symptoms" Life 14, no. 2: 219. https://doi.org/10.3390/life14020219