
Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rhamnolipids
2.2. Cell Line and Viruses
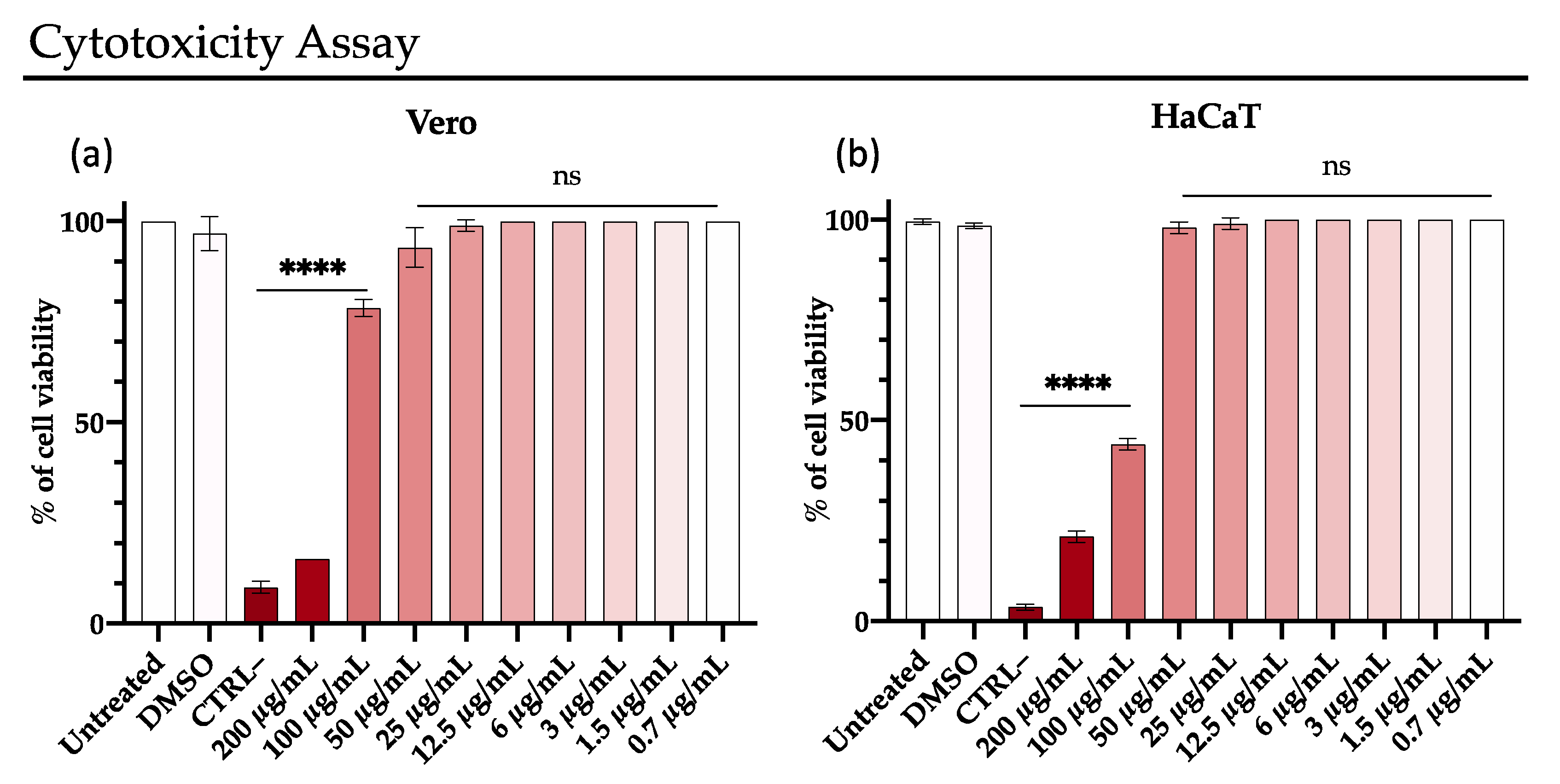
2.3. Cytotoxicity Assay
2.4. Antiviral Assays
2.4.1. Co-treatment Assay
2.4.2. Virus Pre-treatment Assay
2.4.3. Post-treatment Assay
2.4.4. Cell Pre-treatment
2.4.5. Viral Attachment and Entry/Fusion Assay
- 2 × 105 cells/well were seeded in a 12-well plate and incubated at 37 °C overnight. The plate was pre-chilled to 4 °C for 30 min before testing. The cells were co-treated with virus and substance in the 1:1 ratio, and the plate was incubated at 4 °C for 1 h. Then, the monolayer was washed twice with PBS, rinsed with fresh medium, and the plate was incubated at 37 °C for 48 h.
- Vero cells (2 × 105 cells/well) were plated in a 12-well plate and incubated for 24 h in a humidified atmosphere with 5% CO2 at 37 °C. Before testing, the cell monolayer was cooled to 4 °C. The cells were infected with the virus (103 PFU/mL) and were incubated at 4 °C for 1 h. Successively, the supernatant was removed, and the cell monolayer gently washed to remove the excess virus. Then, M15RL was added to each well, and the plate was incubated at 37 °C for 1 h. Finally, the supernatant was removed, rinsed with fresh medium, and incubated for 48 h.
2.5. Real-Time PCR
2.6. Microscopy
2.6.1. Fluorescent Microscopy
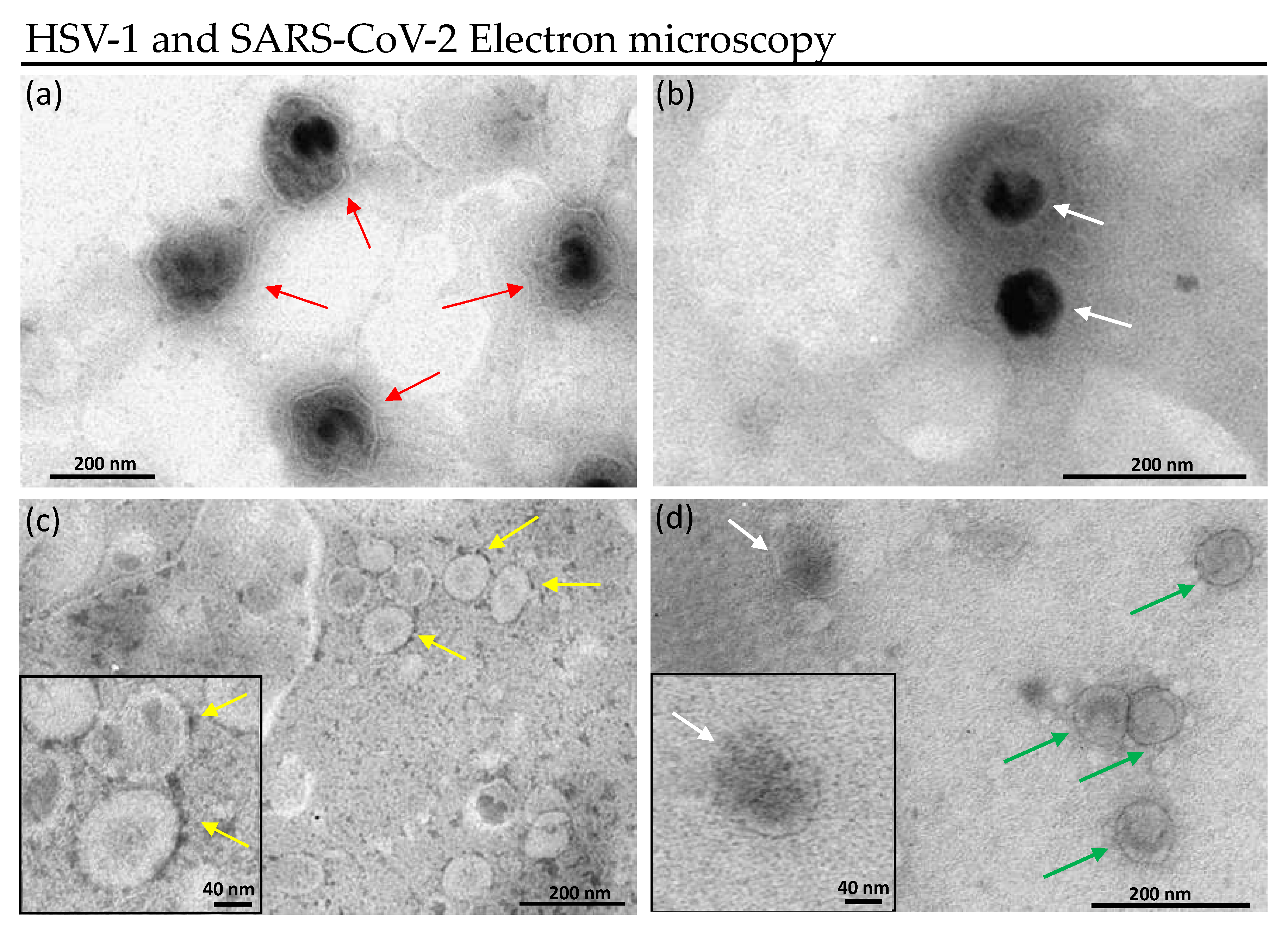
2.6.2. Transmission Electron Microscopy
2.7. Coating of Plastic Surface with M15RL Mixture
2.8. Statistical Analysis
3. Results
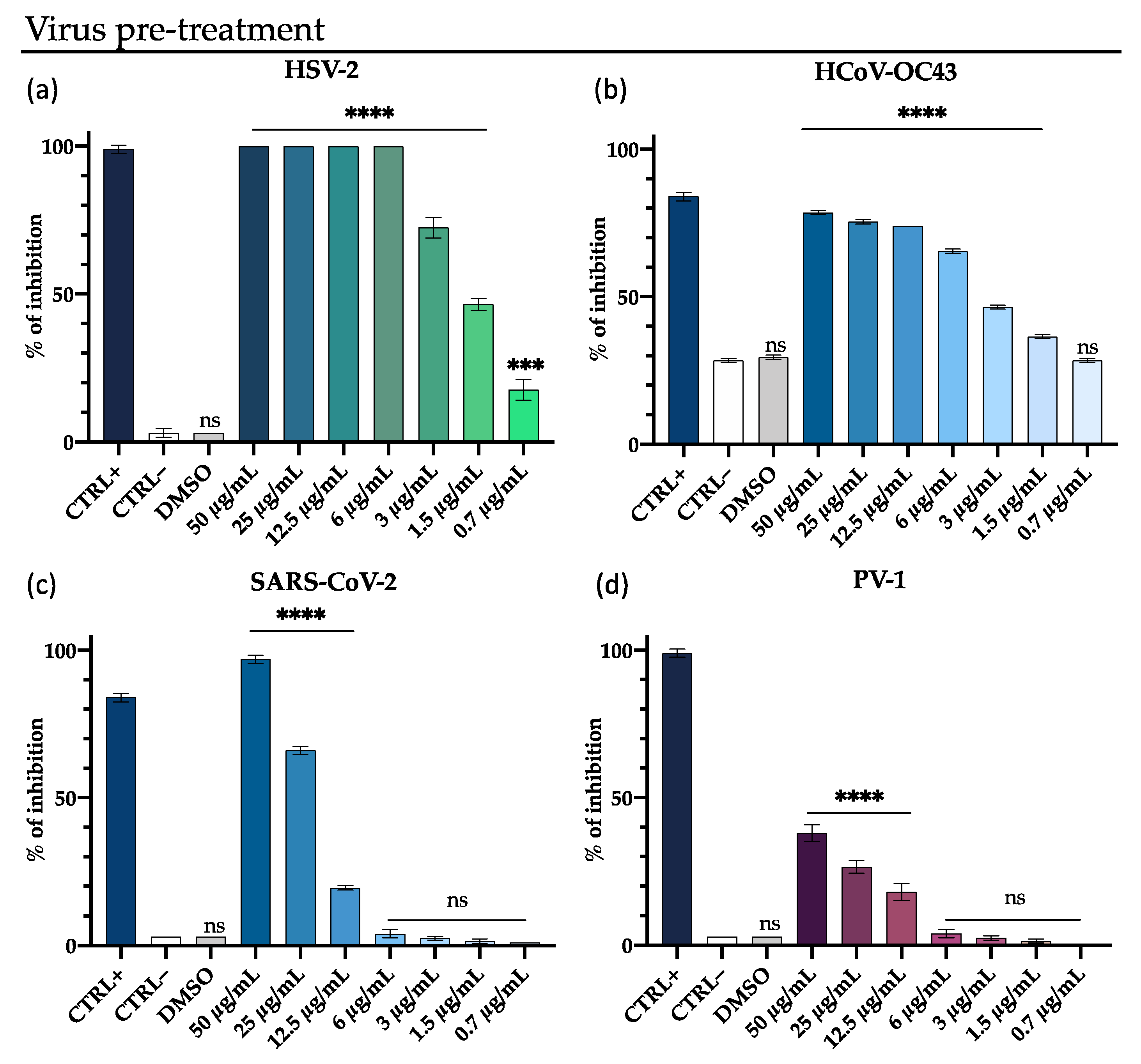
3.1. Antiviral Activity
3.2. M15RL Mode of Action on HSV-1
- Cells pre-treatment assay: the Vero cells were treated with M15RL and incubated for 1 h at 37 °C to investigate if action is also directed on the cells, preventing the viral infection. Then, cell monolayers were infected with HSV-1 for 1 h.
- Attachment assay: in most enveloped viruses, including HSV-1, viral entry/fusion is stopped at 4 °C [44,48]. Thus, cells were pre-chilled at 4 °C for 30 min, treated simultaneously with virus and M15RL, and incubated at 4 °C for 1 h, which allow viral attachment, but not penetration. This assay could evaluate if M15RL interferes with the virus binding to the cells.
- Fusion assay: cells were pre-cooled at 4 °C for 1 h, then infected and incubated at 4 °C for 1 h, allowing the virus to attach to the cells. Then the HSV-1 inoculum was removed, the cells treated with M15RL, and incubated for 1h to verify if the mixture could block the penetration of the virus in the host cell after its binding to the membrane.
- Post-treatment assay: the cells were first incubated with HSV-1 for 1 h at 37 °C, then M15RL was added and incubated for 1 h at 37 °C to study if the mixture could act on viral replication stages.
- Quantification of early and late viral genes: mRNA early gene UL52 levels, encoding for DNA primase, and late gene UL27, for structural envelope glycoprotein B, were quantified after 48 h of infection.
Treated Virions Visualization
3.3. Antiviral Activity on RLs-Coated Surface
3.4. Rhamnolipids Cytotoxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avula, K.; Singh, B.; Kumar, P.V.; Syed, G.H. Role of Lipid Transfer Proteins (LTPs) in the Viral Life Cycle. Front. Microbiol. 2021, 12, 3509. [Google Scholar] [CrossRef]
- Hong, C.; Oksanen, H.M.; Liu, X.; Jakana, J.; Bamford, D.H.; Chiu, W. A Structural Model of the Genome Packaging Process in a Membrane-Containing Double Stranded DNA Virus. PLoS Biol. 2014, 12, e1002024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, D.P. Virus Morphology, Replication, and Assembly. Viral Ecol. 2000, 63–124. [Google Scholar] [CrossRef]
- Rey, F.A.; Lok, S.-M. Common Features of Enveloped Viruses and Implications for Immunogen Design for Next-Generation Vaccines. Cell 2018, 172, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wang, Q.; Wen, Y.; Jiang, S.; Lu, L. Protein- and Peptide-Based Virus Inactivators: Inactivating Viruses Before Their Entry into Cells. Front. Microbiol. 2020, 11, 1063. [Google Scholar] [CrossRef]
- Nath, P.; Kabir, M.A.; Doust, S.K.; Ray, A. Diagnosis of Herpes Simplex Virus: Laboratory and Point-of-Care Techniques. Infect. Dis. Rep. 2021, 13, 518–539. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, C.; Lopez, L. Cutaneous Infections Caused by Herpesviridae: A Review. Arch. Pathol. Lab. Med. 2011, 135, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Sadowski, L.A.; Upadhyay, R.; Greeley, Z.W.; Margulies, B.J. Current Drugs to Treat Infections with Herpes Simplex Viruses-1 and -2. Viruses 2021, 13, 1228. [Google Scholar] [CrossRef] [PubMed]
- Deethae, A.; Peerapornpisal, Y.; Pekkoh, J.; Sangthong, P.; Tragoolpua, Y. Inhibitory effect of Spirogyra spp. algal extracts against herpes simplex virus type 1 and 2 infection. J. Appl. Microbiol. 2018, 124, 1441–1453. [Google Scholar] [CrossRef] [PubMed]
- WHO. Herpes Simplex Virus. Available online: https://www.who.int/news-room/fact-sheets/detail/herpes-simplex-virus (accessed on 27 August 2021).
- Tognarelli, E.I.; Palomino, T.F.; Corrales, N.; Bueno, S.M.; Kalergis, A.M.; González, P.A. Herpes Simplex Virus Evasion of Early Host Antiviral Responses. Front. Cell. Infect. Microbiol. 2019, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- WHO. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 27 August 2021).
- Liu, D.X.; Liang, J.Q.; Fung, T.S. Human Coronavirus-229E, -OC43, -NL63, and -HKU1 (Coronaviridae). Encycl. Virol. 2021, 428–440. [Google Scholar] [CrossRef]
- NCIRD COVID-19. Available online: https://www.cdc.gov/coronavirus/types.html (accessed on 27 August 2021).
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Inès, M.; Dhouha, G. Glycolipid biosurfactants: Potential related biomedical and biotechnological applications. Carbohydr. Res. 2015, 416, 59–69. [Google Scholar] [CrossRef] [PubMed]
- De Giani, A.; Zampolli, J.; Di Gennaro, P. Recent Trends on Biosurfactants With Antimicrobial Activity Produced by Bacteria Associated with Human Health: Different Perspectives on Their Properties, Challenges, and Potential Applications. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Jahan, R.; Bodratti, A.M.; Tsianou, M.; Alexandridis, P. Biosurfactants, natural alternatives to synthetic surfactants: Physicochemical properties and applications. Adv. Colloid Interface Sci. 2020, 275, 102061. [Google Scholar] [CrossRef] [PubMed]
- Seydlová, G.; Svobodová, J. Review of surfactin chemical properties and the potential biomedical applications. Cent. Eur. J. Open Med. 2008, 3, 123–133. [Google Scholar] [CrossRef]
- Sobrinho, H.; Luna, J.; Rufino, R.; Porto, A.; Porto, F.; Sarubbo, L. Biosurfactants: Classification, Properties and Environmental Applications. Recent Dev. Biotechnol. 2013, 11, 1–29. [Google Scholar]
- Tedesco, P.; Maida, I.; Palma Esposito, F.; Tortorella, E.; Subko, K.; Ezeofor, C.C.; Zhang, Y.; Tabudravu, J.; Jaspars, M.; Fani, R.; et al. Antimicrobial Activity of Monoramnholipids Produced by Bacterial Strains Isolated from the Ross Sea (Antarctica). Mar. Drugs 2016, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Çelik, P.A.; Manga, E.B.; Çabuk, A.; Banat, I.M. Biosurfactants’ Potential Role in Combating COVID-19 and Similar Future Microbial Threats. Appl. Sci. 2021, 11, 334. [Google Scholar] [CrossRef]
- Silva, V.L.; Lovaglio, R.B.; Von Zuben, C.J.; Contiero, J. Rhamnolipids: Solution against Aedes aegypti? Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Zu, Y.; Li, X.; Meng, Q.; Long, X. Recent progress towards industrial rhamnolipids fermentation: Process optimization and foam control. Bioresour. Technol. 2020, 298, 122394. [Google Scholar] [CrossRef] [PubMed]
- Herzog, M.; Tiso, T.; Blank, L.M.; Winter, R. Interaction of rhamnolipids with model biomembranes of varying complexity. Biochim. Et Biophys. Acta Biomembr. 2020, 1862, 183431. [Google Scholar] [CrossRef]
- Alan, S. Evonik Commercializes Biosurfactants. Focus Surfactants 2016, 2016, 3–4. [Google Scholar] [CrossRef]
- Insights, F.B. Surfactants MArket Size, Share & COVID-19 Impact Analysis, By Type (Anionic, Nonionic, Cationic, and Amphoteric), By Application (Home Care, Personal Care, Textile, Food & Beverages, Industrial & Istitutional Cleaning, Plastics, and Others), and Regional Forecast, 2020–2027. Available online: https://www.alliedmarketresearch.com/surfactant-market (accessed on 27 August 2021).
- Paulino, B.N.; Pessôa, M.G.; Mano, M.C.R.; Molina, G.; Neri-Numa, I.A.; Pastore, G.M. Current status in biotechnological production and applications of glycolipid biosurfactants. Appl. Microbiol. Biotechnol. 2016, 100, 10265–10293. [Google Scholar] [CrossRef]
- Roy, S.; Chandni, S.; Das, I.; Karthik, L.; Kumar, G.; Bhaskara Rao, K.V. Aquatic model for engine oil degradation by rhamnolipid producing Nocardiopsis VITSISB. 3 Biotech 2015, 5, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Kiran, G.S.; Ninawe, A.S.; Lipton, A.N.; Pandian, V.; Selvin, J. Rhamnolipid biosurfactants: Evolutionary implications, applications and future prospects from untapped marine resource. Crit. Rev. Biotechnol. 2016, 36, 399–415. [Google Scholar] [CrossRef]
- Thakur, P.; Saini, N.K.; Thakur, V.K.; Gupta, V.K.; Saini, R.V.; Saini, A.K. Rhamnolipid the Glycolipid Biosurfactant: Emerging trends and promising strategies in the field of biotechnology and biomedicine. Microb. Cell Factories 2021, 20, 1–15. [Google Scholar] [CrossRef]
- Abalos, A.; Pinazo, A.; Infante, M.R.; Casals, M.; García, F.; Manresa, A. Physicochemical and Antimicrobial Properties of New Rhamnolipids Produced by Pseudomonas aeruginosa AT10 from Soybean Oil Refinery Wastes. Langmuir 2001, 17, 1367–1371. [Google Scholar] [CrossRef]
- Nitschke, M.; Costa, S.G.V.A.O. Biosurfactants in food industry. Trends Food Sci. Technol. 2007, 18, 252–259. [Google Scholar] [CrossRef]
- Stipcevic, T.; Piljac, A.; Piljac, G. Enhanced healing of full-thickness burn wounds using di-rhamnolipid. Burns 2006, 32, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Borah, S.N.; Goswami, D.; Sarma, H.K.; Cameotra, S.S.; Deka, S. Rhamnolipid Biosurfactant against Fusarium verticillioides to Control Stalk and Ear Rot Disease of Maize. Front. Microbiol. 2016, 7, 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christova, N.; Tuleva, B.; Kril, A.; Georgieva, M.; Konstantinov, S.; Terziyski, I.; Nikolova, B.; Stoineva, I. Chemical structure and in vitro antitumor activity of rhamnolipids from Pseudomonas aeruginosa BN10. Appl. Biochem. Biotechnol. 2013, 170, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Remichkova, M.; Galabova, D.; Roeva, I.; Karpenko, E.; Shulga, A.; Galabov, A.S. Anti-herpesvirus activities of Pseudomonas sp. S-17 rhamnolipid and its complex with alginate. Z. Nat. C J. Biosci. 2008, 63, 75–81. [Google Scholar] [CrossRef]
- Jin, L.; Black, W.; Sawyer, T. Application of Environment-Friendly Rhamnolipids against Transmission of Enveloped Viruses Like SARS-CoV2. Viruses 2021, 13, 322. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactants Produced by Marine Microorganisms with Therapeutic Applications. Mar. Drugs 2016, 14, 38. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, C.; Tedesco, P.; Vitale, G.A.; Esposito, F.P.; Giugliano, R.; Monti, M.C.; D’Auria, M.V.; de Pascale, D. Characterization of a New Mixture of Mono-Rhamnolipids Produced by Pseudomonas gessardii Isolated from Edmonson Point (Antarctica). Mar. Drugs 2020, 18, 269. [Google Scholar] [CrossRef]
- Safrin, S.; Elbeik, T.; Mills, J. A rapid screen test for in vitro susceptibility of clinical herpes simplex virus isolates. J. Infect. Dis. 1994, 169, 879–882. [Google Scholar] [CrossRef]
- Zannella, C.; Giugliano, R.; Chianese, A.; Buonocore, C.; Vitale, G.A.; Sanna, G.; Sarno, F.; Manzin, A.; Nebbioso, A.; Termolino, P.; et al. Antiviral Activity of Vitis vinifera Leaf Extract against SARS-CoV-2 and HSV-1. Viruses 2021, 13, 1263. [Google Scholar] [CrossRef] [PubMed]
- Squillaci, G.; Zannella, C.; Carbone, V.; Minasi, P.; Folliero, V.; Stelitano, D.; Cara, F.; Galdiero, M.; Franci, G.; Morana, A. Grape Canes from Typical Cultivars of Campania (Southern Italy) as a Source of High-Value Bioactive Compounds: Phenolic Profile, Antioxidant and Antimicrobial Activities. Molecules 2021, 26, 2746. [Google Scholar] [CrossRef]
- Tai, C.J.; Li, C.L.; Tai, C.J.; Wang, C.K.; Lin, L.T. Early Viral Entry Assays for the Identification and Evaluation of Antiviral Compounds. J. Vis. Exp. 2015, e53124. [Google Scholar] [CrossRef] [Green Version]
- Montazeri Aliabadi, H.; Totonchy, J.; Mahdipoor, P.; Parang, K.; Uludağ, H. Suppression of Human Coronavirus 229E Infection in Lung Fibroblast Cells via RNA Interference. Front. Nanotechnol. 2021, 3, 34. [Google Scholar] [CrossRef]
- Mas, V.; Melero, J.A. Entry of enveloped viruses into host cells: Membrane fusion. Subcell. Biochem. 2013, 68, 467–487. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, J.; Nguyen, L.N.T.; Adkins, J.L.; Schank, M.; Khanal, S.; Nguyen, L.N.; Dang, X.; Cao, D.; Thakuri, B.K.C.; et al. Blockade of SARS-CoV-2 spike protein-mediated cell–cell fusion using COVID-19 convalescent plasma. Sci. Rep. 2021, 11, 5558. [Google Scholar] [CrossRef]
- Madan, R.P.; Mesquita, P.M.; Cheshenko, N.; Jing, B.; Shende, V.; Guzman, E.; Heald, T.; Keller, M.J.; Regen, S.L.; Shattock, R.J.; et al. Molecular umbrellas: A novel class of candidate topical microbicides to prevent human immunodeficiency virus and herpes simplex virus infections. J. Virol. 2007, 81, 7636–7646. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Shen, S.; Wang, L.; Deng, H. Role of tegument proteins in herpesvirus assembly and egress. Protein Cell 2010, 1, 987–998. [Google Scholar] [CrossRef]
- Heilingloh, C.S.; Krawczyk, A. Role of L-Particles during Herpes Simplex Virus Infection. Front. Microbiol. 2017, 8, 2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Z.; Oton, J.; Qu, K.; Cortese, M.; Zila, V.; McKeane, L.; Nakane, T.; Zivanov, J.; Neufeldt, C.J.; Cerikan, B.; et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature 2020, 588, 498–502. [Google Scholar] [CrossRef]
- Wilson, V.G. Growth and differentiation of HaCaT keratinocytes. Methods Mol. Biol. 2014, 1195, 33–41. [Google Scholar] [CrossRef]
- Feher, P.; Ujhelyi, Z.; Varadi, J.; Fenyvesi, F.; Roka, E.; Juhasz, B.; Varga, B.; Bombicz, M.; Priksz, D.; Bacskay, I.; et al. Efficacy of Pre- and Post-Treatment by Topical Formulations Containing Dissolved and Suspended Silybum marianum against UVB-Induced Oxidative Stress in Guinea Pig and on HaCaT Keratinocytes. Molecules 2016, 21, 1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firquet, S.; Beaujard, S.; Lobert, P.E.; Sane, F.; Caloone, D.; Izard, D.; Hober, D. Survival of Enveloped and Non-Enveloped Viruses on Inanimate Surfaces. Microbes Environ. 2015, 30, 140–144. [Google Scholar] [CrossRef] [Green Version]
- UNAIDS. Global HIV & AIDS Statistics—Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 7 September 2021).
- Welker, L.; Paillart, J.-C.; Bernacchi, S. Importance of Viral Late Domains in Budding and Release of Enveloped RNA Viruses. Viruses 2021, 13, 1559. [Google Scholar] [CrossRef]
- WHO. Influenza (Avian and Other Zoonotic). Available online: https://www.who.int/news-room/fact-sheets/detail/influenza-(avian-and-other-zoonotic) (accessed on 7 September 2021).
- WHO. Ebola Virus Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 7 September 2021).
- Bryant, A.; Lawrie, T.A.; Dowswell, T.; Fordham, E.J.; Mitchell, S.; Hill, S.R.; Tham, T.C. Ivermectin for Prevention and Treatment of COVID-19 Infection: A Systematic Review, Meta-analysis, and Trial Sequential Analysis to Inform Clinical Guidelines. Am. J. Ther. 2021, 28, e434–e460. [Google Scholar] [CrossRef]
- NCIRD Hand Hygiene Recommendations. Guidance for Healthcare Providers about Hand Hygiene and COVID-19. Available online: https://www.cdc.gov/coronavirus/2019-ncov/hcp/hand-hygiene.html (accessed on 15 September 2021).
- WHO. Guide to Local Production: WHO-Recommended Handrub Formulations. Available online: https://www.who.int/gpsc/5may/Guide_to_Local_Production.pdf (accessed on 28 August 2021).
- Palma Esposito, F.; Giugliano, R.; Della Sala, G.; Vitale, G.A.; Buonocore, C.; Ausuri, J.; Galasso, C.; Coppola, D.; Franci, G.; Galdiero, M.; et al. Combining OSMAC Approach and Untargeted Metabolomics for the Identification of New Glycolipids with Potent Antiviral Activity Produced by a Marine Rhodococcus. Int. J. Mol. Sci. 2021, 22, 9055. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; Veit, M.; Osterrieder, K.; Gradzielski, M. Surfactants—Compounds for inactivation of SARS-CoV-2 and other enveloped viruses. Curr. Opin. Colloid Interface Sci. 2021, 55, 101479. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.W.; Ng, L.T.; Chiang, L.C.; Lin, C.C. Antiviral effects of saikosaponins on human coronavirus 229E in vitro. Clin. Exp. Pharm. Physiol. 2006, 33, 612–616. [Google Scholar] [CrossRef]
- Ryu, W.-S. Virus Life Cycle. Mol. Virol. Hum. Pathog. Viruses 2017, 31–45. [Google Scholar] [CrossRef]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479–480, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, D.S. Virus entry: Molecular mechanisms and biomedical applications. Nat. Rev. Microbiol. 2004, 2, 109–122. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, J.; Wang, L.; Li, F.; Wang, Y.; Jiang, Y.; Song, X.; Qin, S.; Zheng, K.; Ye, J.; et al. Anti-HSV-1 activity of Aspergillipeptide D, a cyclic pentapeptide isolated from fungus Aspergillus sp. SCSIO 41501. Virol. J. 2020, 17, 41. [Google Scholar] [CrossRef] [Green Version]
- Korting, H.C.; Herzinger, T.; Hartinger, A.; Kerscher, M.; Angerpointner, T.; Maibach, H.I. Discrimination of the irritancy potential of surfactants in vitro by two cytotoxicity assays using normal human keratinocytes, HaCaT cells and 3T3 mouse fibroblasts: Correlation with in vivo data from a soap chamber assay. J. Derm. Sci. 1994, 7, 119–129. [Google Scholar] [CrossRef]
- Kyadarkunte, A.; Patole, M.; Pokharkar, V. In Vitro Cytotoxicity and Phototoxicity Assessment of Acylglutamate Surfactants Using a Human Keratinocyte Cell Line. Cosmetics 2014, 1, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Rusanov, A.L.; Luzgina, N.G.; Lisitsa, A.V. Sodium Dodecyl Sulfate Cytotoxicity towards HaCaT Keratinocytes: Comparative Analysis of Methods for Evaluation of Cell Viability. Bull. Exp. Biol. Med. 2017, 163, 284–288. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Family | Genus | Nucleic Acid | Symmetry | Envelope | Dimensions |
|---|---|---|---|---|---|---|
| HSV-1 | Herpesviridae | Simplexvirus | dsDNA | icosahedral | yes | 155–240 nm |
| HSV-2 | Herpesviridae | Simplexvirus | dsDNA | icosahedral | yes | 155–240 nm |
| HCoV-229E | Coronaviridae | Alphacoronavirus | ssRNA(+) | helical | yes | 80–120 nm |
| HCoV-OC43 | Coronaviridae | Betacoronavirus | ssRNA(+) | helical | yes | 80–120 nm |
| SARS-CoV-2 | Coronaviridae | Betacoronavirus | ssRNA(+) | helical | yes | ≈100 nm |
| PV-1 | Picornaviridae | Enterovirus | ssRNA(+) | icosahedral | no | ≈30 nm |
| Host | Gene | Forward Sequence | Reverse Sequence |
|---|---|---|---|
| HSV-1 | UL52 | GACCGACGGGTGCGTTATT | GAAGGAGTCGCCATTTAGCC |
| HSV-1 | UL27 | GCCTTCTTCGCCTTTCGC | CGCTCGTGCCCTTCTTCTT |
| SARS-CoV-2 | S | AGGTTGATCACAGGCAGACT | GCTGACTGAGGGAAGGAC |
| Vero | GAPDH | CCTTTCATTGAGCTCCAT | CGTACATGGGAGCGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giugliano, R.; Buonocore, C.; Zannella, C.; Chianese, A.; Palma Esposito, F.; Tedesco, P.; De Filippis, A.; Galdiero, M.; Franci, G.; de Pascale, D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics 2021, 13, 2121. https://doi.org/10.3390/pharmaceutics13122121
Giugliano R, Buonocore C, Zannella C, Chianese A, Palma Esposito F, Tedesco P, De Filippis A, Galdiero M, Franci G, de Pascale D. Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses. Pharmaceutics. 2021; 13(12):2121. https://doi.org/10.3390/pharmaceutics13122121
Chicago/Turabian StyleGiugliano, Rosa, Carmine Buonocore, Carla Zannella, Annalisa Chianese, Fortunato Palma Esposito, Pietro Tedesco, Anna De Filippis, Massimiliano Galdiero, Gianluigi Franci, and Donatella de Pascale. 2021. "Antiviral Activity of the Rhamnolipids Mixture from the Antarctic Bacterium Pseudomonas gessardii M15 against Herpes Simplex Viruses and Coronaviruses" Pharmaceutics 13, no. 12: 2121. https://doi.org/10.3390/pharmaceutics13122121